3.1. Morphometric Measurements
Female cicada killer right wing length averaged 29.40 ± 0.19(46) mm. Cicada killers weighed 0.991 ± 0.021(46) g and had a thorax mass of 0.418 ± 0.009(46) g resulting in an unladen FMR of 0.401 ± 0.004(46).
All prey of
S. convallis were identified as
Tibicen parallelus Davis (Hemiptera: Cicadidae), not only for the present study, but also for the hundreds of specimens examined in the course of other research projects at this site during this period. Cicada right wing length was 36.21 ± 0.20(46) mm. Cicada body mass averaged 1.127 ± 0.024(46) g, resulting in a mean loaded wasp FMR
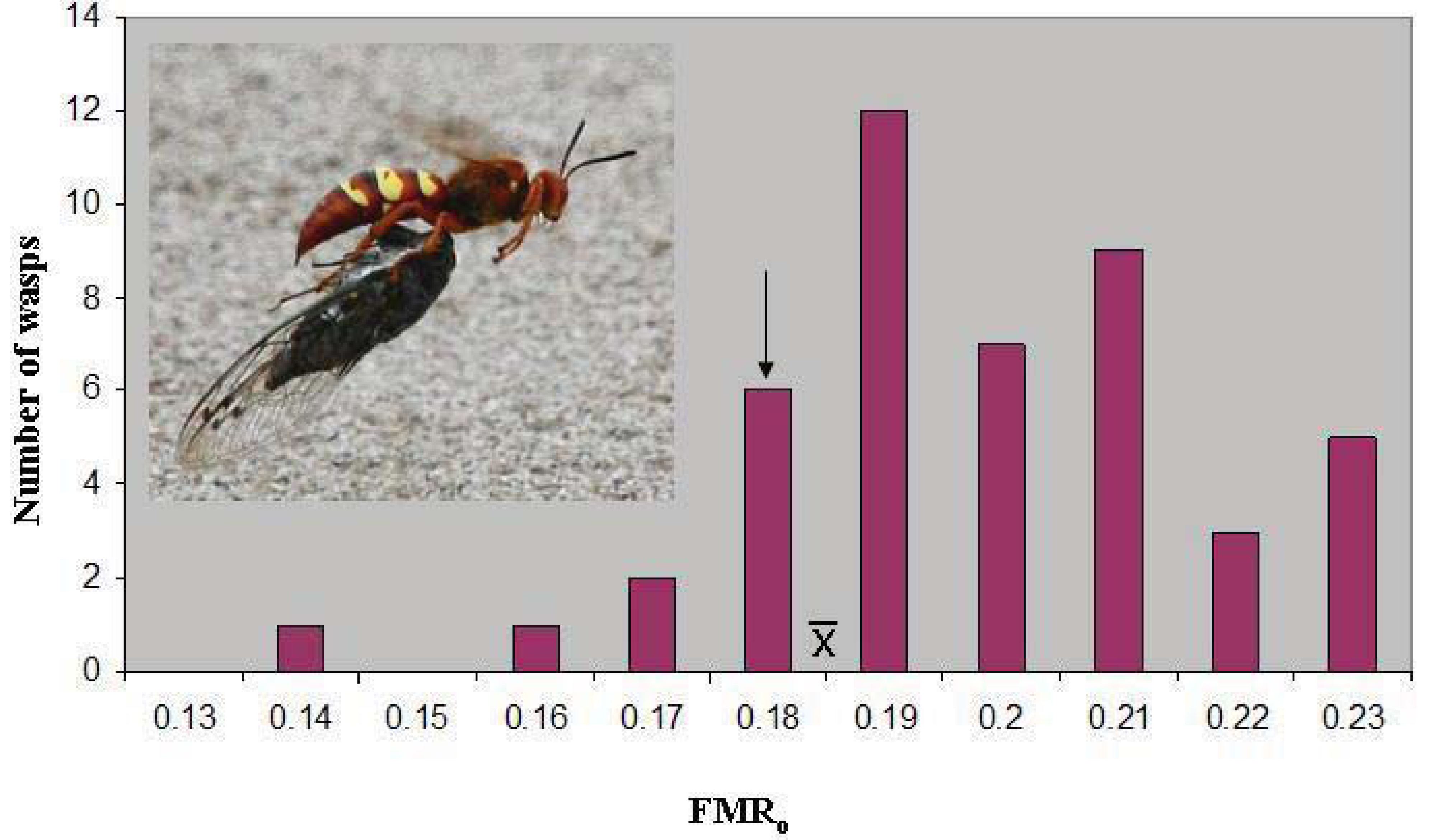
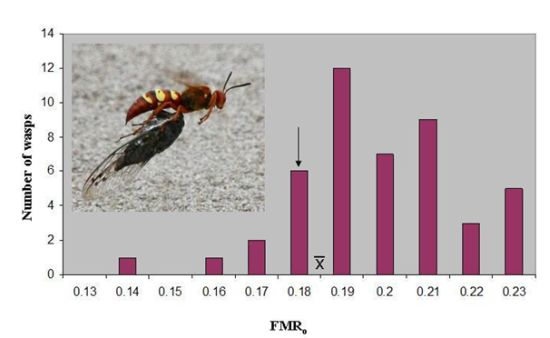
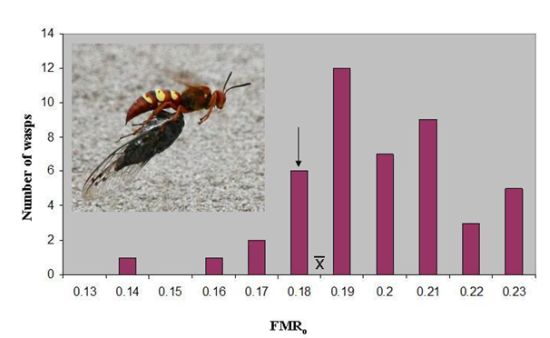
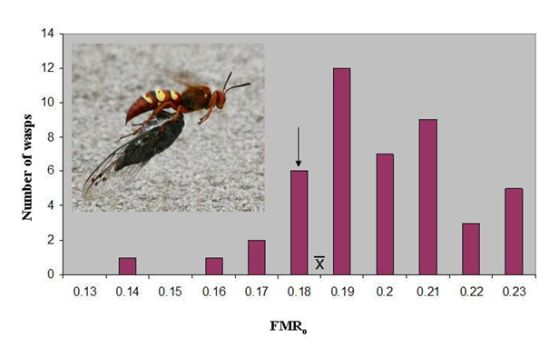
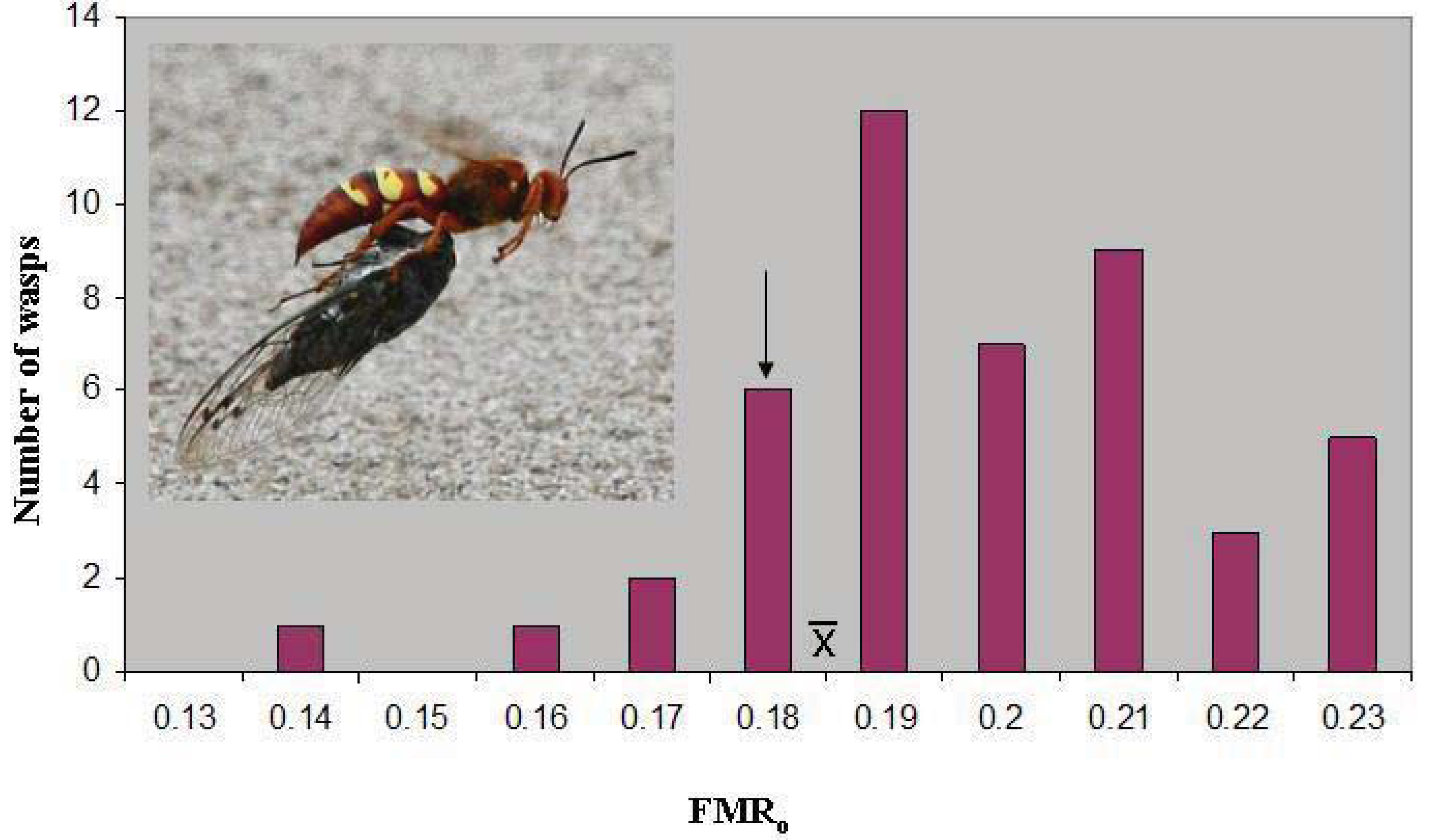
o of 0.187 ± 0.003(46). The latter value lies just above the marginal level, and only seven wasps (15%) were below 0.179 (
Figure 1). However, the marginal level lies outside the 95% confidence interval of the mean (0.186–0.189), and the distribution has a skewness of −0.387. The median FMR
o, 0.186, is essentially identical to the mean.
The nearness of the mean FMR
o to the marginal FMR suggests that
S. convallis is a nearly ideal flying predator with respect to loading. However, the mean loaded FMR is slightly above the marginal level, indicating slight underloading. The marginal FMR is defined as that at which the animal was just able to take off [
10]. Therefore, in most load-bearing female Pacific cicada killers a small safety margin exists. That is, wasps are slightly underloaded, which provides them with additional force production during flight beyond that required to lift the cicada and return to the burrow in flight.
Figure 1.
The distribution of operational flight muscle ratios in
Sphecius convallis foraging on
Tibicen parallelus. The arrow indicates the marginal flight muscle ratio, where vertical take-off is just possible.
![Insects 03 00133 i001]()
indicates the mean. Inset: Pacific cicada killer female carrying cicada in flight.
Figure 1.
The distribution of operational flight muscle ratios in
Sphecius convallis foraging on
Tibicen parallelus. The arrow indicates the marginal flight muscle ratio, where vertical take-off is just possible.
![Insects 03 00133 i001]()
indicates the mean. Inset: Pacific cicada killer female carrying cicada in flight.
This safety margin may have evolved in response to several selection pressures. Female wasps returning to their burrows with prey appear to have highly variable thorax temperatures [
12]. Wasps that do not have optimal thorax temperatures at the time of foraging should experience depressed force production and, hence, decreased ability to carry loads [
13]. Slight underloading may thus compensate for decreased load carrying capacity. Another factor potentially driving higher FMR and the resulting greater flight maneuverability is prey theft. We observed theft of prey by kingbirds while the wasps were carrying cicadas. Our observations suggest that theft pressure can be intense at times [
12]. Therefore, laden wasps with greater FMR would benefit from an improved ability to evade such thievery. Although female cicada killers with prey did not always land at the burrow entrance, they typically landed at least within a meter or two and reached the entrance by walking overland. We never observed wasps climbing objects with their prey, and, in any case, many areas of the mine tailings had no vertical objects available for climbing. Our data indicate that there would seldom be a need to do so, as the wasps have sufficient capacity to fly with their prey.
3.2. Effect of Wasp Size on Cicada Body Mass
The cicada was heavier than the wasp in 35 of 46 wasp/cicada pairs, and on average prey cicadas were significantly heavier than wasps by body mass (P < 0.00005, T-test). A nearly even sex ratio among the prey was observed, as 22 of 46 cicadas were males. Neither wasp body mass, thorax mass, nor right wing length (all indices of size) was significantly related to cicada body mass by linear regression (R2 < 0.05, N = 46). However, it appeared that small wasps used the smallest prey, while larger wasps carried larger cicadas. Hence, wasp data were divided into two groups: large wasps, those above the median thorax mass (0.4235 g), and small wasps, those below the median. Thorax mass was chosen as the best index of body size because body mass may vary with nutritional status, hydration state, and number of eggs laid, while wing length is influenced by wear. Large wasps had thorax mass of 0.463 ± 0.006g and carried cicadas weighing 1.181 ± 0.033 g. Small wasps had thorax mass of 0.372 ± 0.009 g and carried cicadas weighing 1.073 ± 0.032 g. Prey mass was significantly greater in large wasps as compared to small wasps (P = 0.011, T-test).
The size of available cicada prey is a good predictor of the sizes of male and female
S. speciosus, though it is not clear whether this effect is nutritional or evolved [
14].
S. speciosus preys on at least 30 species from five genera of cicadas which range a great deal in size [
9]. This large prey diversity may have led to the greater measured variation in wasp size. The coefficient of variation of wing size in female
S. speciosus is 10.1% in a Newberry, FL, population and 8.2% in an Easton, PA, population. The coefficient of variation of wing length was only 4.3% in female
S. convallis and 3.8% in
T. parallelus in the present study, suggesting that there is a tight relationship between the Ruby, AZ, predator and prey populations. Thus, it seems likely that the consistency in
T. parallelus size has led to low variation in
S. convallis size. The fact that this population of cicada killers preys on adult cicadas of a single species with low size variation may have allowed stabilizing selection to adjust the size of adult female wasps to a nearly optimal value.
If cicada killers have a 24.6% conversion efficiency [
12] of cicadas into wasp mass, then four cicadas of average mass will yield one 1.11-g female wasp, which is approximately the average observed mass at Ruby, AZ. A female of that mass is never overloaded. Consistency in provisioning of cells, therefore, is critical to the production of females large enough to carry the prey. Underprovisioning a female larva would result in an undersized female adult that would, in turn, be a poor provisioner. Selection must act on provisioning behavior to maintain the size of adult females.
3.3. Effect of Wasp Body Mass on FMRo
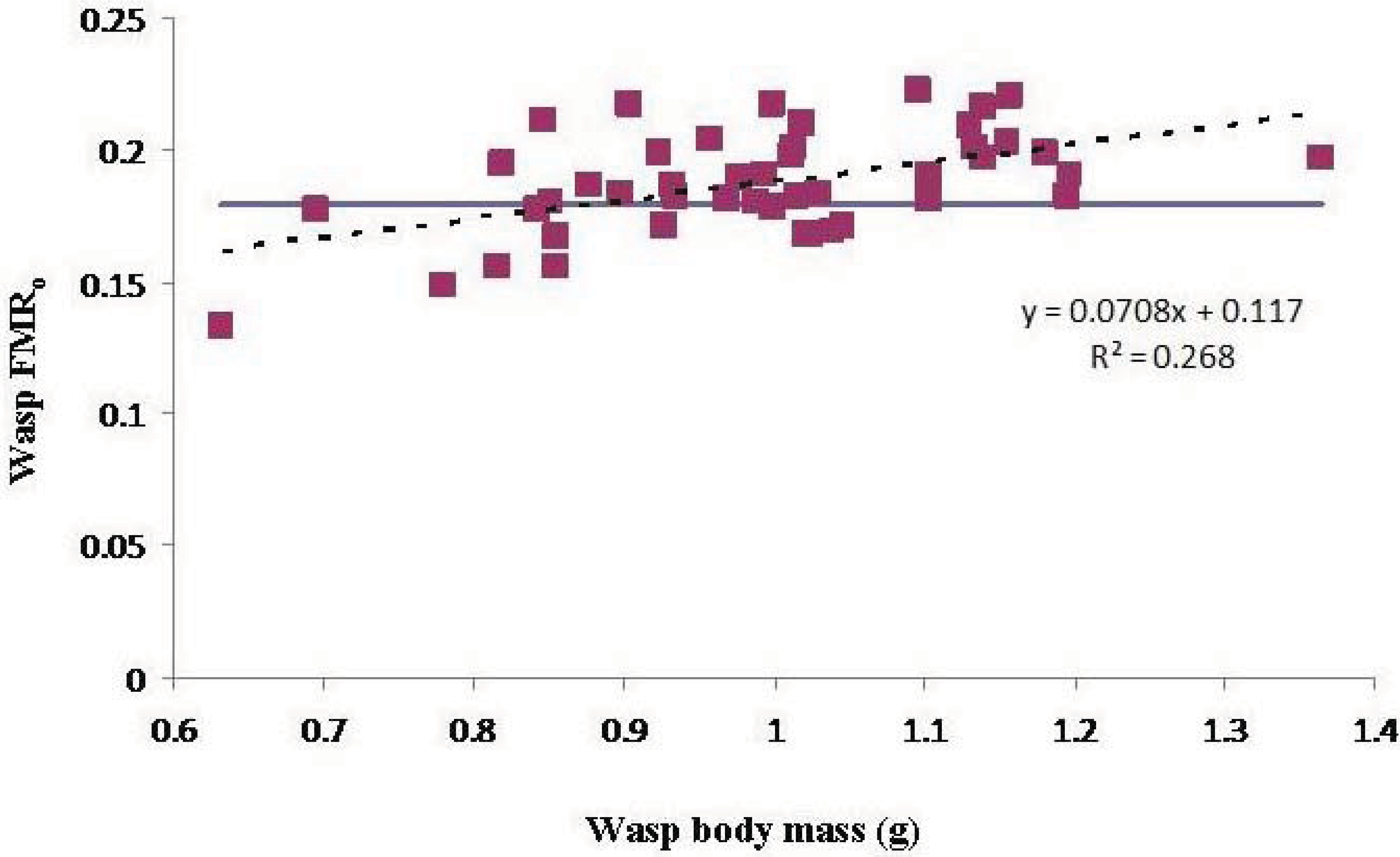
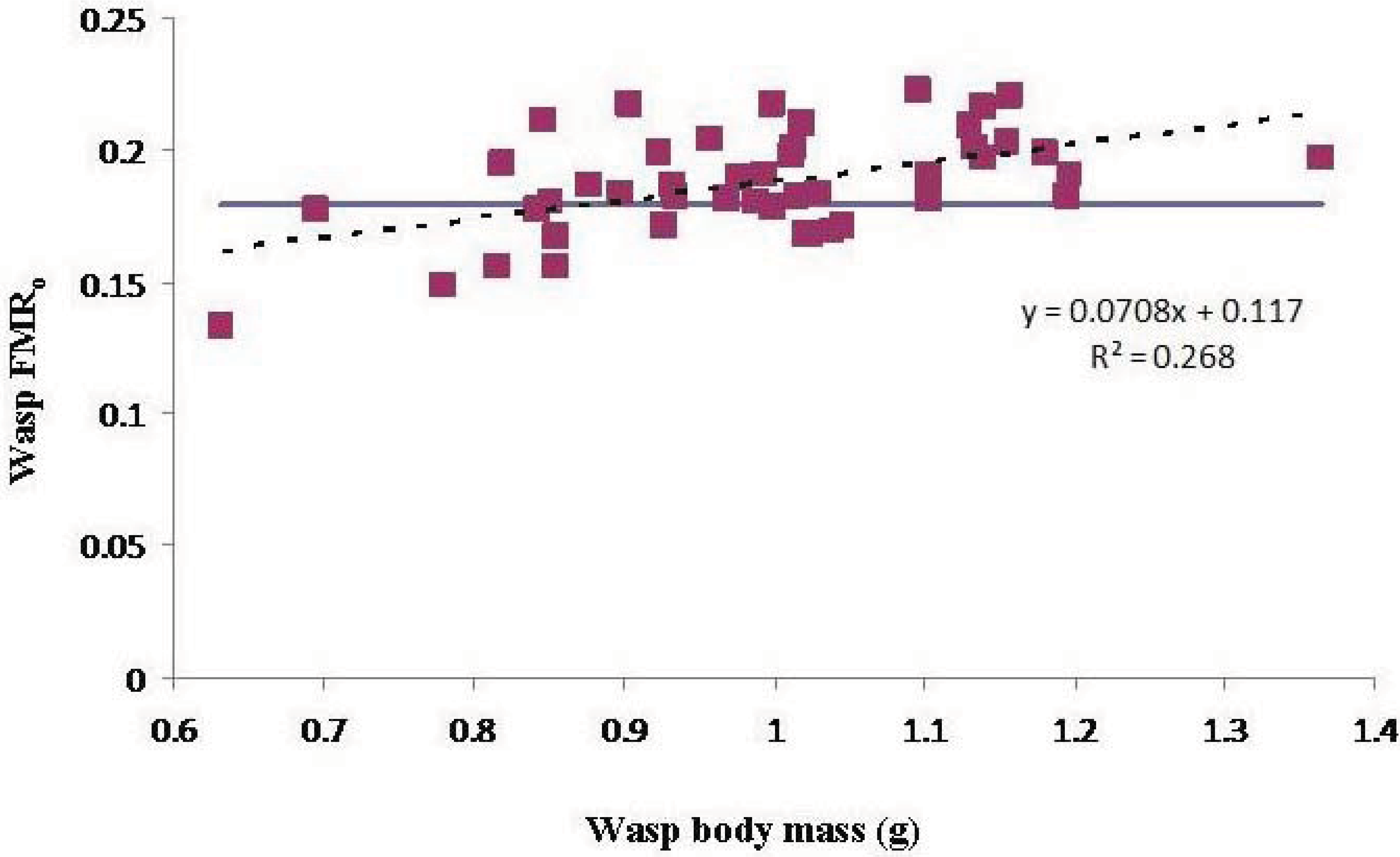
There was a significant effect of wasp M
b on FMR
o, indicating that smaller wasps were overloaded more often than larger wasps (
Figure 2).
Flight muscle mass of a subsample of cicadas was 0.255 ± 0.014 (12) g, resulting in FMR of 0.225 ± 0.012 (12) g. As the FMR of female wasps is 60% greater than that of the cicadas, the wasps should have much greater maneuverability and, hence, no difficulty capturing cicadas on the wing. A similar relationship was demonstrated between
S. speciosus and various
Tibicen prey species [
6], although we are not aware of any confirmed accounts of
Sphecius attacking cicadas in flight.
Some studies appear to indicate that cicada killers do not make choices based on cicada species during foraging. Generally, the species taken are often reflective of those available. Cicada killers may have a generalized cicada search image, and are opportunistic in what they will take. Occasionally, eastern cicada killers even take periodical cicadas [
15], species which they might encounter only every 13 or 17 years, and even then rarely, as the early summer season of
Magicicada overlaps little with the late summer season of cicada killers.
Figure 2.
The effect of wasp body mass on operational flight muscle ratio. The slope of the regression line (dashed) is significant (P < 0.01). The blue line indicates the marginal flight muscle ratio.
Figure 2.
The effect of wasp body mass on operational flight muscle ratio. The slope of the regression line (dashed) is significant (P < 0.01). The blue line indicates the marginal flight muscle ratio.
On the other hand, we never observed S. convallis provisioning with The Grand Western Flood Plain Cicada, Tibicen cultriformis (Davis), which was occasionally heard singing in the area of the S. convallis colony at Ruby, AZ. Its rarity at the site and large size may have resulted in its unsuitability as prey. We heard only one T. cultriformis singing at a time, suggesting that their numbers were few; though we were unable to collect any in the immediate vicinity, we collected several from lower altitudes, where they were much more abundant, and made morphometric measurements. The mean body mass was 3.246 ± 0.083 (14) g. We never observed S. convallis foraging on T. cultriformis. Indeed, if the average S. convallis attempted to carry the average T. cultriformis, a FMRo of 0.097 would result and vertical take-off would be impossible. Hence, even if S. convallis attempted to provision with T. cultriformis, we would be very unlikely to collect them at nest sites. As a result, it is unclear whether T. cultriformis plays a role in S. convallis foraging ecology.
There was some evidence that suggested the possibility that
S. convallis females made size-based choices among individuals of
T. parallelus. In spite of taking generally smaller cicadas, small wasps were the most overloaded. This pattern might be explained, not by prey choice, but by unsuccessful provisioning attempts by small, overloaded wasps. Several lines of evidence suggest indirectly that this occurs. In
Figure 1, the slight negative skewness indicates that the bulk of the values were higher than the mean. In other words, overloaded wasps (those with low FMR
o) were underrepresented in the distribution. If a female takes a cicada that is too large, then it is more likely to be abandoned on the flight back to the burrow; the foraging bout is a failure, and the female goes on to hunt for more cicadas. Consequently, we often find paralyzed cicadas under trees in areas where female eastern cicada killers are hunting [
12], though we did not observe this phenomenon directly in
S. convallis. It also seems more likely that overloaded wasps, having less maneuverability, would be successfully attacked by thieving birds. Failure of small wasps to provision with large cicadas could have produced the statistical result observed; it cannot be concluded that large wasps took cicadas that were larger than expected. Availability of prey was apparently low during the present study, as few cicadas were heard calling in the study area, and foraging bouts required ~8 h/cicada [
12]
vs. 24 min/cicada for
S. speciosus in Pennsylvania [
16]. It is not surprising, then, that clear evidence for prey choice was not detectable.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
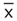 indicates the mean. Inset: Pacific cicada killer female carrying cicada in flight.
indicates the mean. Inset: Pacific cicada killer female carrying cicada in flight.