Collembola from the Gypsum Karst of Sorbas (Almería, Spain), with Descriptions of Three New Species




Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Caves Studied
Caves Without Permanent Water Regimens
Caves with Constant Water Regimens and Which Were Colder Than the Previous Group Included the Following
2.2. Sampling
2.3. Material Processing
2.4. Nomenclature
| MZNA | Museum of Zoology at the University of Navarra, Pamplona, Spain |
| PBAG | Pablo Barranco’s collections in Cueva del Agua |
| PBAP | Pablo Barranco’s collections in Cueva de los Apas |
| PBC3 | Pablo Barranco’s collections in Cueva C 3 |
| PBCA | Pablo Barranco’s collections in Sima del Camión |
| PBCO | Pablo Barranco’s collections in Cueva Covadura |
| PBGE | Pablo Barranco’s collections in Complejo GEP |
| PBTE | Pablo Barranco’s collections in Cueva del Tesoro |
3. Results
3.1. Biocenosis
3.2. Taxonomy
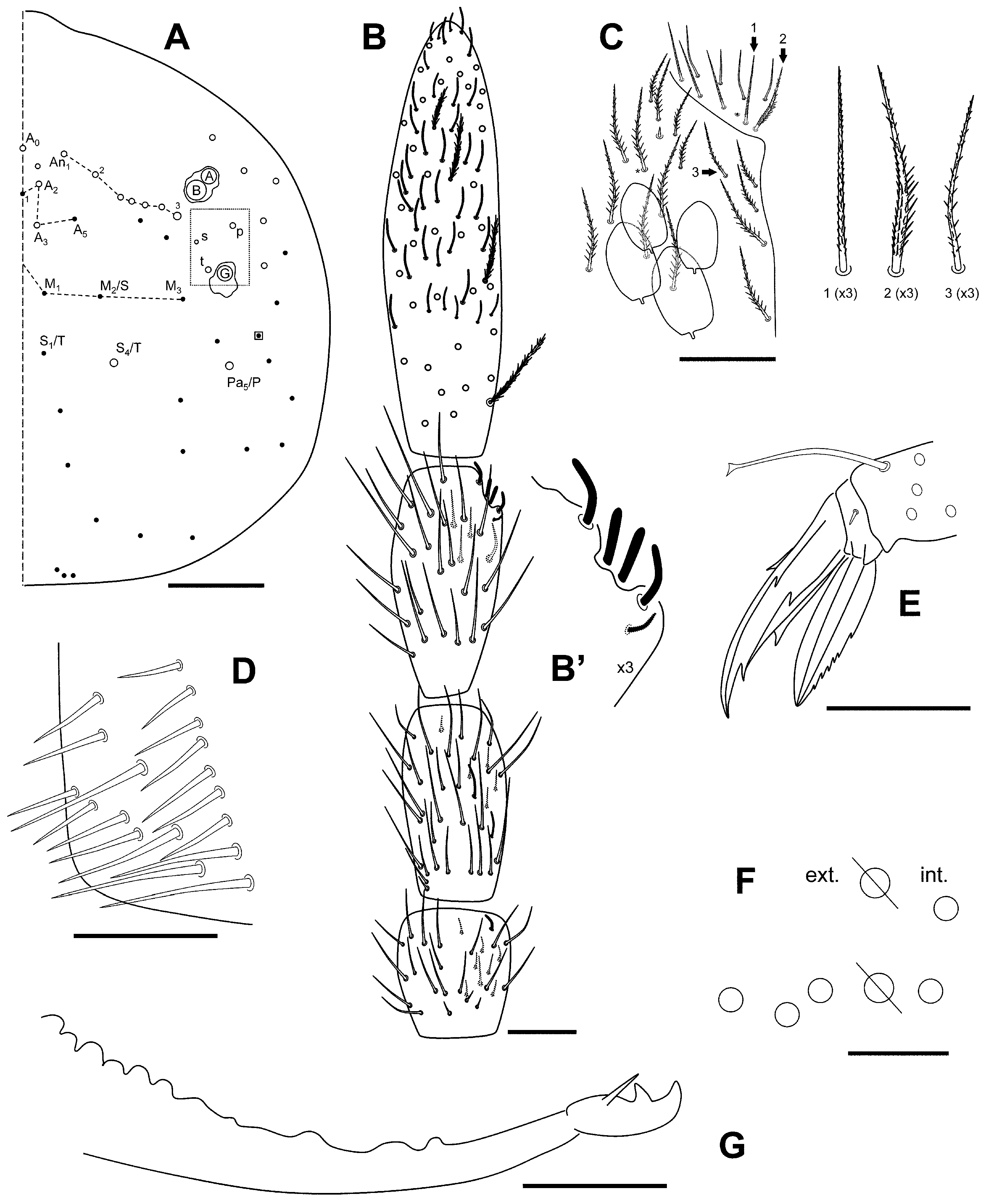
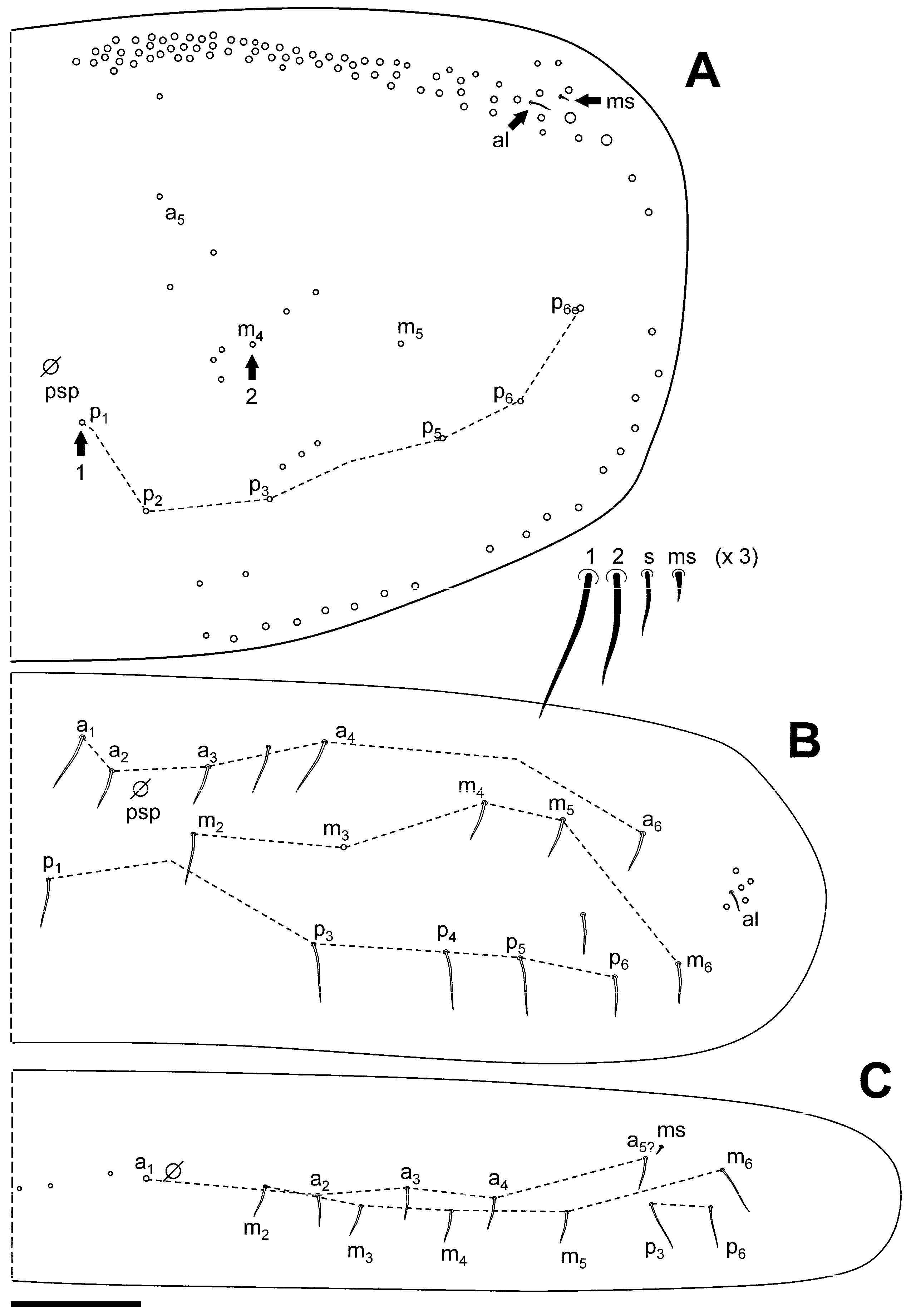
3.2.1. Pseudosinella sexocellata Jordana and Baquero sp. nov.
Type Locality
Type Material

Etymology
Diagnosis
Description
Remarks
3.2.2. Pseudosinella najtae Jordana and Baquero, 2017, in Jordana et al., 2017 [42]
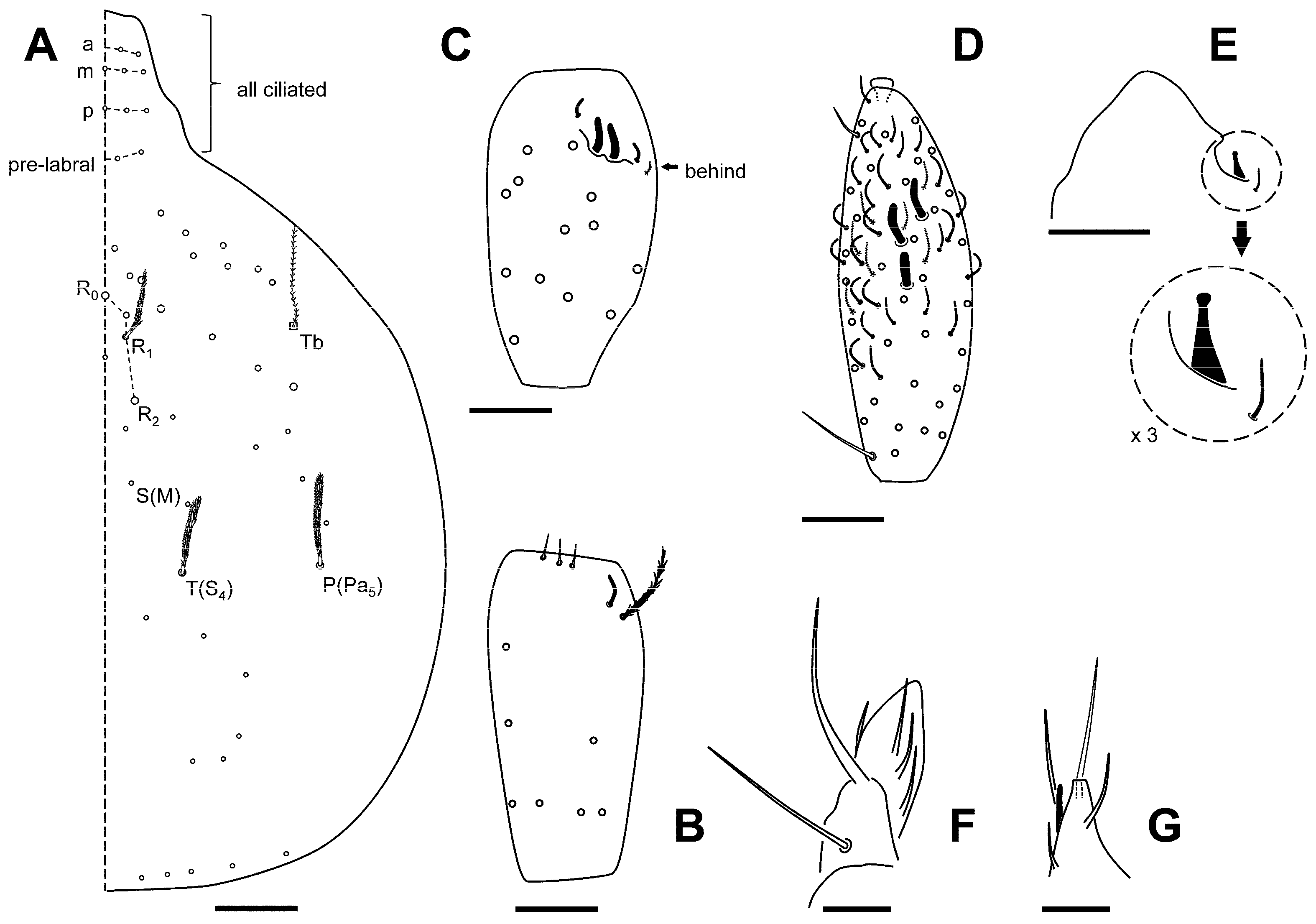
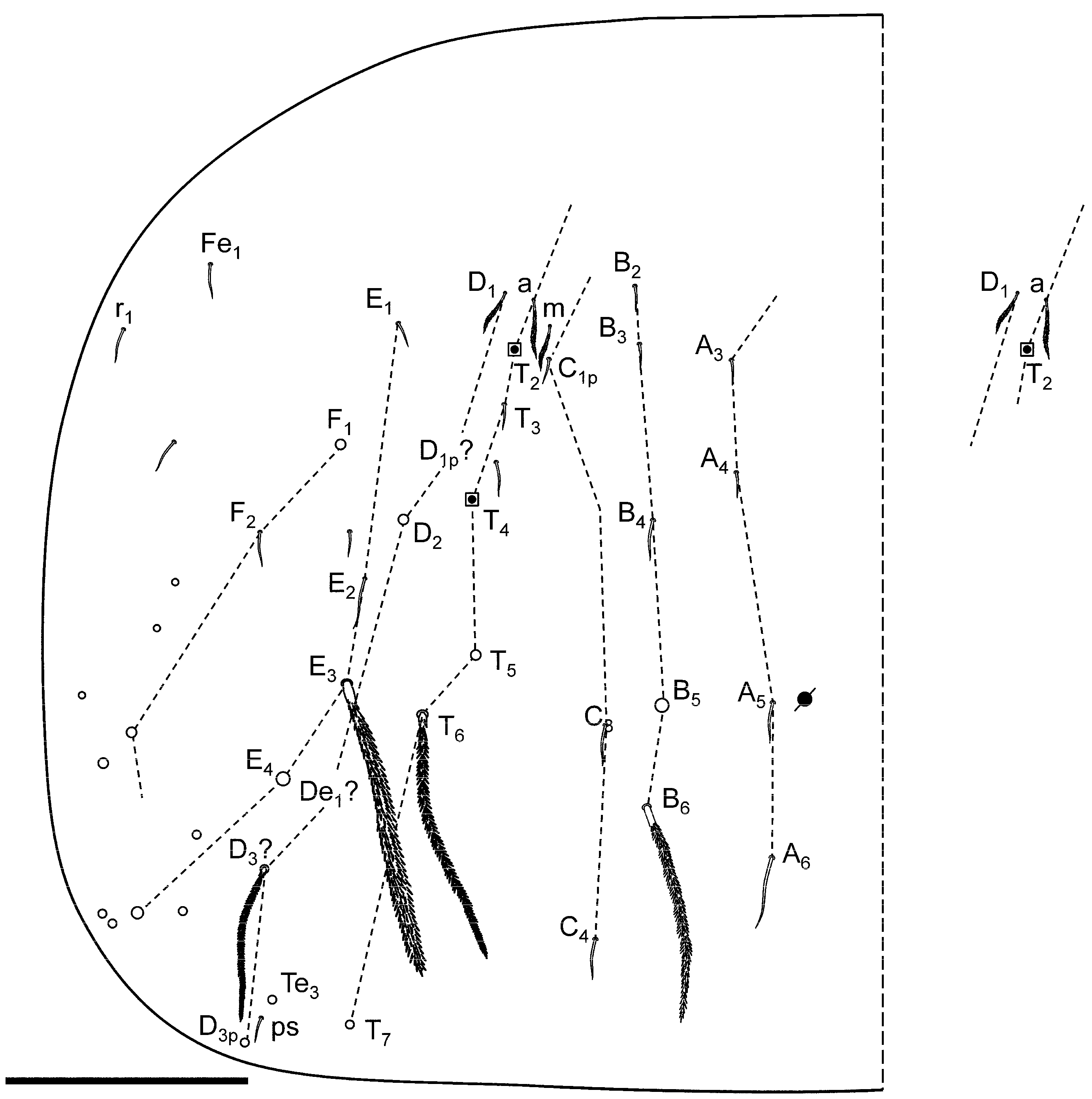
Additional Data to Original Description
3.2.3. Troglopedetes machadoi Delamare-Debouteville, 1946 [46]

Remarks
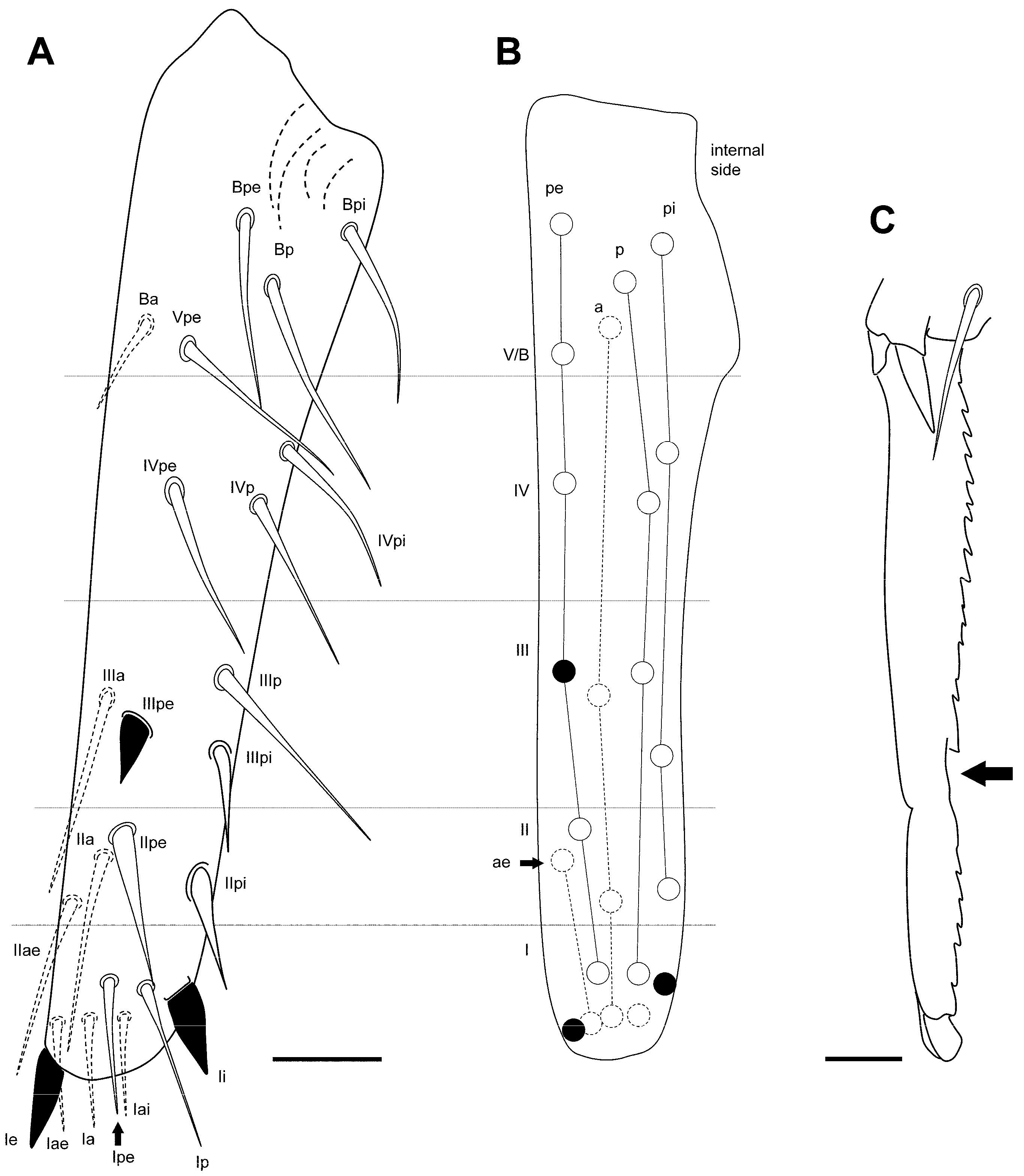
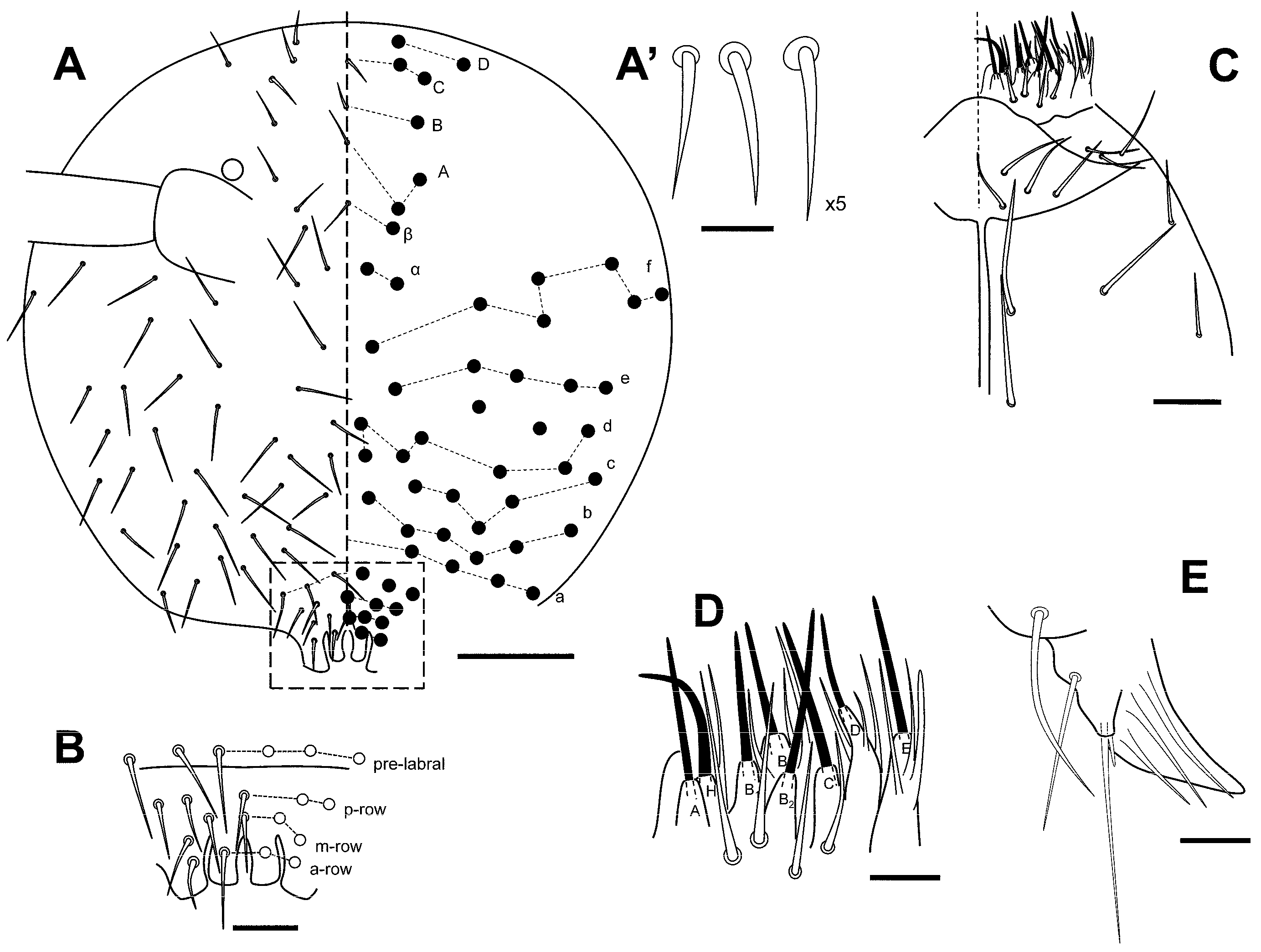
3.2.4. Pygmarrhopalites ruizporteroae Baquero and Jordana, sp. nov.
Type Locality
Type Material
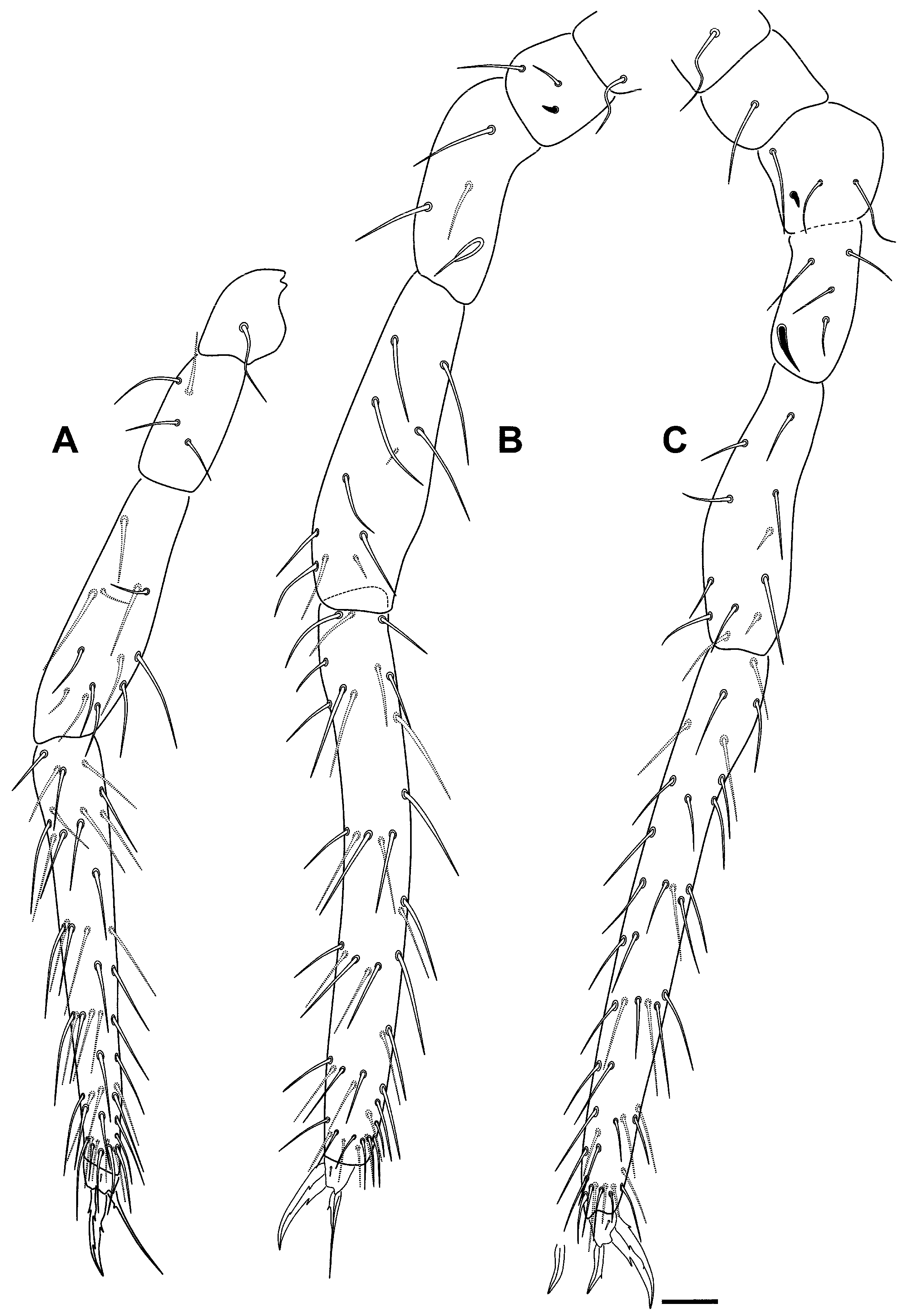
Etymology
Diagnosis
Description
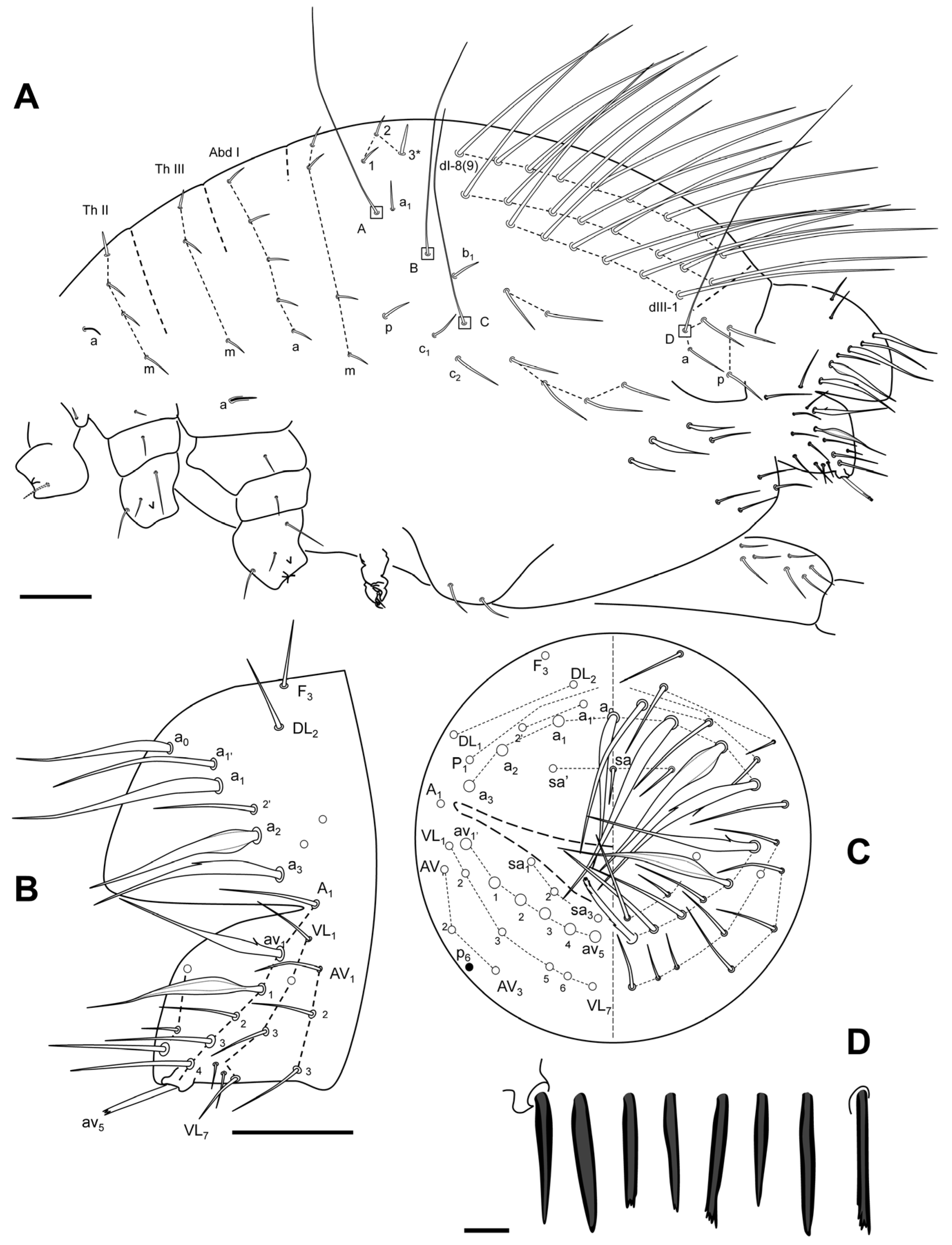

Remarks
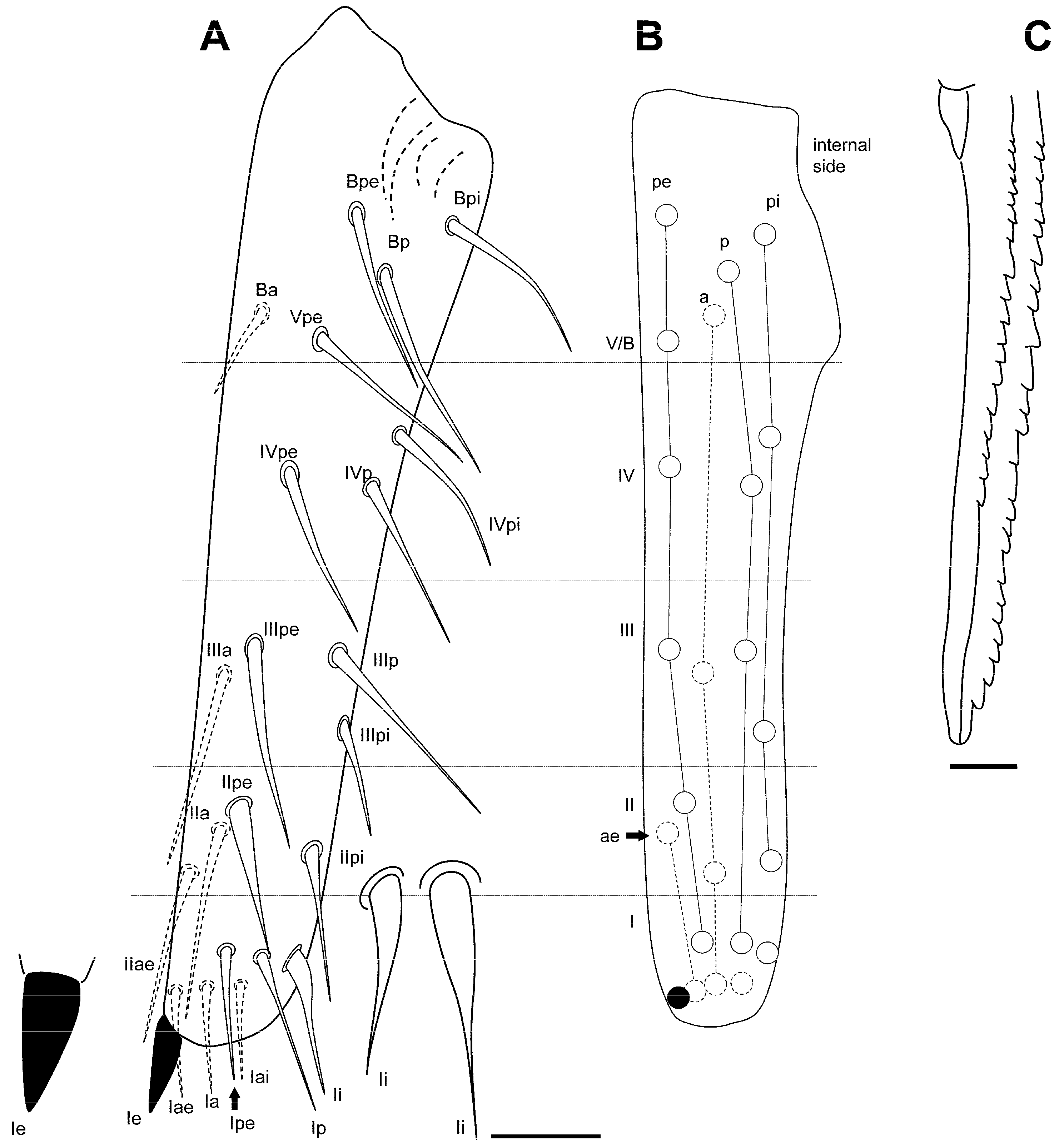
3.2.5. Pygmarrhopalites torresi Baquero and Jordana, sp. nov.
Type Locality
Type Material
Etymology
Diagnosis
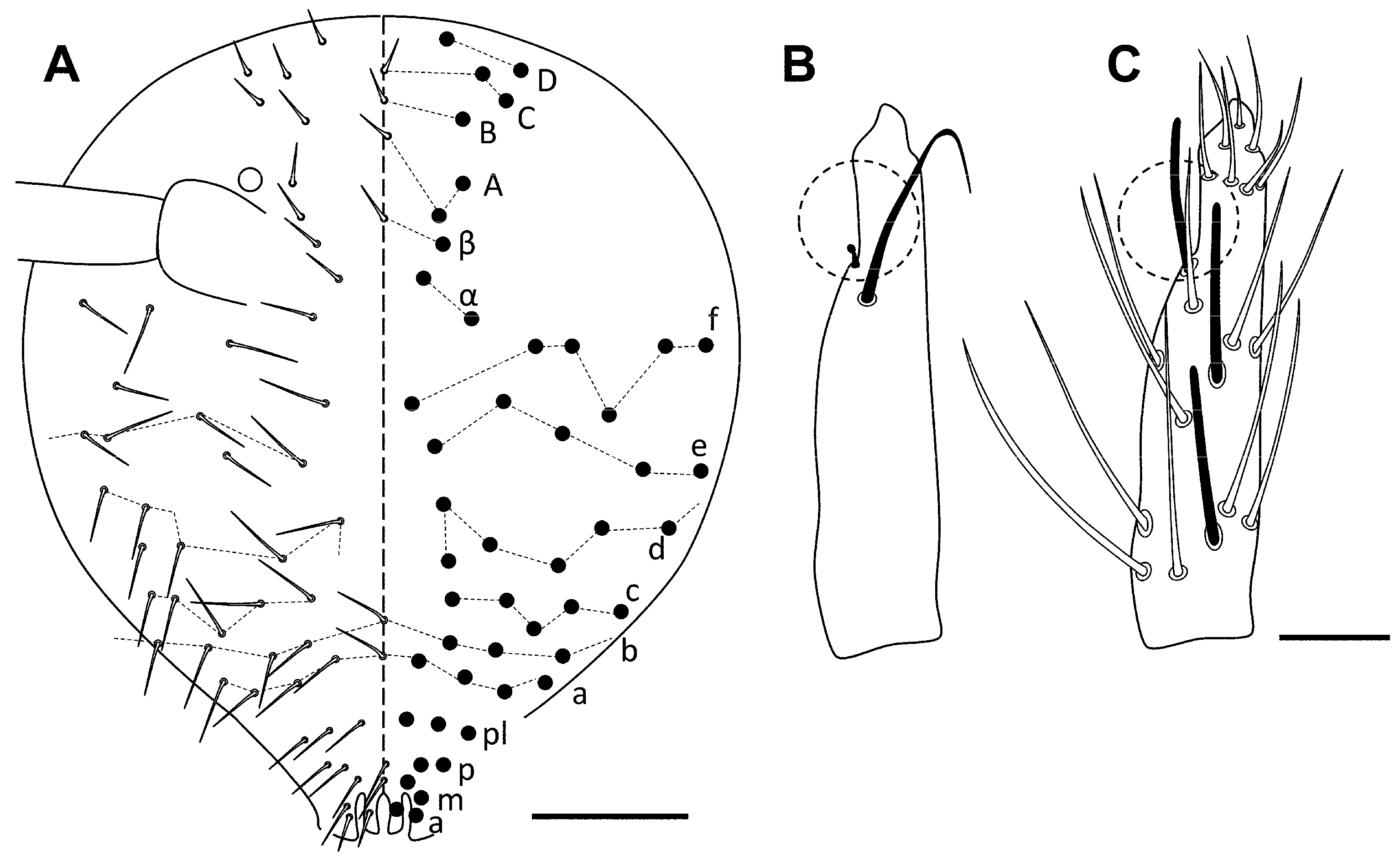
Description

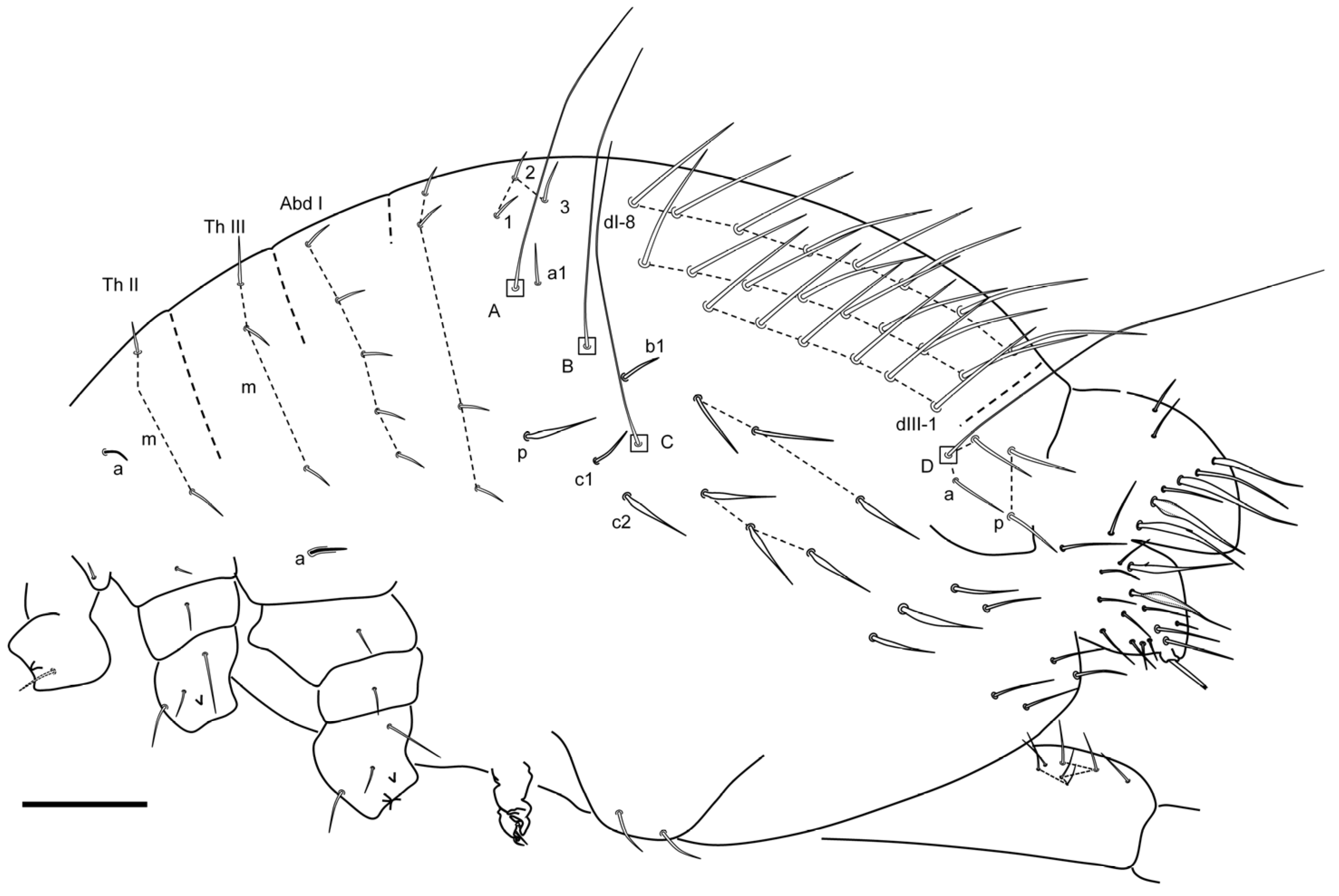
Remarks
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abd | abdominal segment |
| al | anterolateral s-chaeta |
| as | anterosubmedial s-chaeta |
| Ant | antennal or antenna/ae |
| a.s.l. | above sea level |
| GKS | gypsum karst of Sorbas |
| Mc | macrochaeta/ae |
| mes | mesochaeta |
| mic | microchaeta |
| ms | microsensillum/a |
| psp | pseudopore |
| Th | thorax or thoracic segments |
References
- BOJA Ley 3/1989 de 18 de julio por la que se Aprueba el Inventario de Espacios Protegidos de Andalucía. 1989, 60, 3367–3479, Available online: https://www.juntadeandalucia.es/boja/1989/60/1 (accessed on 19 February 2025).
- Calaforra, J.M. The Gypsum Karst of Sorbas—El Karst en yeso de Sorbas; Consejería de Medio Ambiente de la Junta de Andalucía, Ed.; Publicaciones Calle Mayor: Estella, Navarra, 2003; 90p, ISBN 9788460785743. [Google Scholar]
- Calaforra, J.M.; Pulido-Bosch, A. Evolution of the gypsum karst of Sorbas (SE Spain). Geomorphology 2003, 50, 173–180. [Google Scholar] [CrossRef]
- Calaforra, J.M.; Pulido-Bosch, A. Some examples of gypsum karst and the most important gypsum caves in Spain. Int. J. Speleol. 1996, 25, 225–237. [Google Scholar] [CrossRef]
- Calaforra, J.M.; Dell’Aglio, A.; Forti, P. Preliminary data on the chemical erosion in gypsum karst. The Sorbas region (Spain). In Proceedings of the 11th International Congress of Speleology, Beijing, China, 2–8 August 1993; pp. 97–99. [Google Scholar]
- Barranco, P.; Ruiz-Portero, C.; Fernández-Cortés, A.; Bellés, X.; Tinaut, A. Ptinidos de cavidades en yeso de Almería. (Coleoptera, Ptinidae). Boletín De La Asoc. Española De Entomol. 2003, 27, 53–69. [Google Scholar]
- Barranco, P.; Fernández-Cortés, A.; Ruiz-Portero, C.; Ortuño, V. Distribución espacio temporal de Trechus diecki Putzeys, 1870 en las cuevas del Karst en Yeso de Sorbas (Almería, España). (Coleoptera, Carabidae). Boletín SEDECK 2006, 6, 16–25. [Google Scholar]
- Castro, A. Una nueva especie de Blaps Fabricius, 1775 del sureste de España (Coleoptera, Tenebrionidae). Arquivos Entomolóxicos 2014, 12, 237–243. [Google Scholar]
- Sánchez-Camacho, F.J.; Palencia, S.; Barranco, P. Pseláfidos hipogeos de Andalucía (Coleoptera, Staphylinidae, Pselaphinae). In Proceedings of the V Encuentro de Biología Subterránea, Málaga, Spain, 15–17 November 2024; Trabajos del Instituto de Investigación de la Cueva de Nerja: Málaga, Spain, 2024; Volume 2, pp. 30–37, ISBN 978-84 124865 82. [Google Scholar]
- Ribera, C.; De Mas, E.; Barranco, P. Araneidos cavernícolas de la provincia de Almeria (I) y descripción de cuatro especies nuevas. Rev. Ibérica De Aracnol. 2003, 7, 3–17. [Google Scholar]
- Carabajal, E.; García, J.; Rodriguez, F. Descripción de cuatro nuevos pseudoscorpiones cavernícolas de Andalucía, España (Arachnida, Pseudoscorpionida, Chthoniidae). Zoolgica Baetica 2001, 12, 169–184. [Google Scholar]
- Molero, R.; Bach, C.; Sendra, A.; Montagud, S.; Barranco, P.; Gaju, M. Revision of the genus Coletinia (Zygentoma: Nicoletiidae) in the Iberian Peninsula, with description of nine new species. Zootaxa 2013, 3615, 1–60. [Google Scholar] [CrossRef]
- Gómez, M.S.; Barranco, P.; Pérez-Fernández, T. What can fleas tell us about animals inhabiting caves? Boletín Asoc. Española Entomol. 2013, 37, 75–83. [Google Scholar]
- Fernández-Cortés, A. Caracterización Microclimática de Cavidades y Análisis de la Influencia Antrópica de su uso Turístico. Ph.D. Thesis, University of Almería, Almería, Spain, 2005; 425p. [Google Scholar]
- Gázquez, F.; Calaforra, J.M.; Forti, P.; De Waele, J.; Sanna, L. The role of condensation in the evolution of dissolutional forms in gypsum caves: Study case in the karst of Sorbas (SE Spain). Geomorphology 2015, 229, 100–111. [Google Scholar] [CrossRef]
- Vargovitsh, R.S. New cave Arrhopalitidae (Collembola: Symphypleona) from the Crimea (Ukraine). Zootaxa 2009, 2047, 1–47. [Google Scholar] [CrossRef]
- Fjellberg, A. The maxillary outer lobe, an important systematic tool in Isotomidae (Collembola). Ann. Société R. Zool. Belg. 1984, 114, 83–88. [Google Scholar]
- Fjellberg, A. The labial palp in Collembola. Zool. Anz. 1999, 237, 309–330. [Google Scholar]
- Christiansen, K.; Bellinger, P.F. Cave Arrhopalites: New to science. J. Cave Karst Stud. 1996, 58, 168–180. [Google Scholar]
- Bretfeld, G. Synopses on Palaearctic Collembola, Vol. 2. Symphypleona. Abh. Berichte Naturkundemuseums Görlitz 1999, 71, 1–318. [Google Scholar]
- Christiansen, K. The genus Arrhopalites (Collembola: Sminthuridae) in the United States and Canada. Speleology 1966, 2, 43–73. [Google Scholar] [CrossRef]
- Christiansen, K.; Bellinger, P. The Collembola of North America North of the Rio Grande, A Taxonomic Analysis; Grinnell College: Grinnell, IA, USA, 1998; 1520p. [Google Scholar]
- Vargovitsh, R.S. Two new troglobiont Pygmarrhopalites species of the principalis group (Collembola: Arrhopalitidae) from the West Caucasus. Zootaxa 2017, 4250, 23–42. [Google Scholar] [CrossRef]
- Gisin, H.; da Gama, M.M. Espèces nouvelles de Pseudosinella cavernicoles (Insecta: Collembola). Rev. Suisse Zool. 1969, 76, 143–181. [Google Scholar] [CrossRef]
- Szeptycki, A. Chaetotaxy of the Entomobryidae and its phylogenetical significance. In Morpho-Systematic Studies on Collembola IV; Polska Akademia Nauk: Kraków, Poland, 1979; 219p. [Google Scholar]
- Soto-Adames, F.N. Two new species and descriptive notes for five Pseudosinella species (Hexapoda: Collembola: Entomobryidae) from West Virginian (USA) caves. Zootaxa 2010, 2331, 1–34. [Google Scholar]
- Mateos, E. Definition of Lepidocyrtus lusitanicus Gama, 1964 species-complex (Collembola, Entomobryidae), with description of new species and color forms from the Iberian Peninsula. Zootaxa 2008, 1917, 38–54. [Google Scholar] [CrossRef]
- Jordana, R.; Baquero, E.; Ariño, A.H. Collembola Delta Database: Pseudosinella Taxonomy. University of Navarra. (Continuously Updated). 2024. Available online: http://www.unav.es/unzyec/collembola/Pseudosinella/ (accessed on 5 December 2024).
- Gisin, H. Nouvelles notes taxonomiques sur les Lepidocyrtus. Rev. Ecol. Biol. Sol. 1965, 2, 524. [Google Scholar]
- Gisin, H. Deux Lepidocyrtus nouveaux pour l’Espagne (Collembola). Eos Rev. Esp. Entomol. 1967, 42, 393–395. [Google Scholar]
- Gisin, H. Espèces nouvelles et lignées évolutives de Pseudosinella endogés (Collembola). Mem. Estudos Mus. Zool. Univ. Coimbra 1967, 301, 1–25. [Google Scholar]
- Baquero, E.; Jordana, R.; Labrada, L.; González-Luque, C. A new species of Pseudosinella Schäffer, 1897 (Collembola, Entomobryidae) from Altamira Caves (Cantabria, Spain). Zookeys 2020, 989, 39–54. [Google Scholar] [CrossRef]
- Lubbock, J. Monograph of the Collembola and Thysanura; Ray Society: London, UK, 1873; 276p. [Google Scholar] [CrossRef]
- Börner, C. Die Familien der Collembolen. Zool. Anz. 1913, 41, 315–322. [Google Scholar]
- Soto-Adames, F.N.; Barra, J.A.; Christiansen, K.; Jordana, R. Suprageneric Classification of Collembola Entomobryomorpha. Ann. Entomol. Soc. Am. 2008, 101, 501–513. [Google Scholar] [CrossRef]
- Wahlgren, E. Apterygoten aus Ägypten und dem Sudan nebst Bemerkungen zur Verbreitung und Systematik der Collembolen. In Results of the Swedish Zoological Expedition to Egypt and the White Nile 1901 under the Direction of L.A. Jägerskiöld; K.W. Appalbergs Boktryckeri: Uppsala, Sweden, 1906; 72p. [Google Scholar]
- Zhang, F.; Deharveng, L. Systematic revision of Entomobryidae (Collembola) by integrating molecular and new morphological evidence. Zool. Scr. 2015, 44, 298–311. [Google Scholar] [CrossRef]
- Schäffer, C. Apterygoten. Hamb. Magal. Samm. 1897, 2, 1–48. [Google Scholar]
- Simón-Benito, J.C.; Palacios-Vargas, J.G. Four new Pseudosinella (Collembola: Entomobryidae) from La Rioja, Spain. Zootaxa 2008, 1728, 59–68. [Google Scholar] [CrossRef]
- Schött, H. Etudes sur les collemboles du nord. Kungl. Sven. Vetenskapsakademiens Handl. 1902, 28, 1–48. [Google Scholar]
- Bonet, F. Estudios sobre los Colémbolos cavernícolas con especial referencia a los de la Fauna Española. Mem. Real Soc. Española Hist. Nat. 1931, 14, 231–403. [Google Scholar]
- Jordana, R.; Barranco, P.; Amezcua, A.; Baquero, E. Two new species of Collembola (Hexapoda) from Saliente Cave (Almera, Spain). Zoosystema 2017, 39, 103–115. [Google Scholar] [CrossRef]
- Börner, C. Das System der Collembolen nebst Beschreibung neuer Collembolen des Hamburger Naturhistorischen Museums. Mitteilungen Aus Dem Naturhistorischen Mus. Hambg. 1906, 23, 147–188. [Google Scholar]
- Zhang, F.; Bellini, B.C.; Soto-Adames, F.N. New insights into the systematics of Entomobryoidea (Collembola: Entomobryomorpha): First instar chaetotaxy, homology and classification. Zool. Syst. 2019, 44, 249–278. [Google Scholar] [CrossRef]
- Joseph, G. Beobachtungen über die Lebensweise und Vorkommen den in den Krainer Gebirgsgrotten einheimischen Arten der blinden Gattungen Machaerites Leptodirus Oryotus Und Troglorrhynchus. Jahres-Ber. Schlesischen Ges. Vaterländische Cult. 1872, 49, 171–182. [Google Scholar]
- Delamare-Deboutteville, C. Collemboles cavernicoles du Portugal récoltés par A. de Barros Machado (Deuxième série). Rev. Française D’entomologie 1946, 13, 100–104. [Google Scholar]
- Gama, M.M. Colêmbolos das Canarias (Insectos, Apterigotas). In Proceedings of the Congreso Ibérico de Entomología, Córdoba, Spain, 1988; pp. 73–90. Available online: https://www.biodiversidadcanarias.es/biota/documento/A00488 (accessed on 19 February 2025).
- Martínez, M.; Baquero, E.; Barranco, P.; Ariño, A.H.; Jordana, R. A new genus and species of Collembola from caves of south Iberian Peninsula (Collembola, Poduromorpha, Onychiuridae). Zootaxa 2004, 734, 1–15. [Google Scholar] [CrossRef]
- Mañas-Jordá, S.; Acosta Rivas, C.R.; Ariño, A.H.; Baquero, E.; Bartomeus, I.; Bonada, N.; García-Barros, E.; García-Meseguer, A.J.; García Roselló, E.; Lobo, J.M.; et al. IberArthro: A Database Compiling Taxonomic and Distributional Data on Ibero-Balearic Arthropods; Version 2.4; Department of Ecology and Hydrology, University of Murcia: Murcia, Spain, 2025. [Google Scholar] [CrossRef]
- Börner, C. Neue Collembolenformen und zur Nomenclatur der Collembola Lubbock. Zool. Anz. 1901, 24, 696–712. [Google Scholar]
- Bretfeld, G. Phylogenetic systematics of the higher taxa of Symphypleona Börner, 1901 (Insecta, Entognatha, Collembola). In Proceedings of 2nd International Seminary of Apterygota Siena; Dallai, R., Ed.; University of Siena: Siena, Italy, 1986; Volume 1, pp. 302–311. [Google Scholar]
- Bretfeld, G. Sturmius epiphytus n. gen. n. spec. from Colombia, a taxon of the Symphypleona (Insecta, Colembola) with an unexpected character combination. Description and position in non-Linnean and Linnean classifications of the Symphypleona. Zool. Syst. Evol. Resour. 1994, 32, 264–281. [Google Scholar] [CrossRef]
- Stach, J. The Apterygotan Fauna of Poland in Relation to the World-Fauna of This Group of Insects, Family: Sminthuridae; Polska Akademia Nauk, PWN: Kraków, Poland, 1956; 287p. [Google Scholar]
- Jordana, R.; Fadrique, F.; Baquero, E. The collembolan fauna of Maestrazgo caves (Teruel, Spain) with description of three new species. Zootaxa 2012, 3502, 49–71. [Google Scholar] [CrossRef]
- Wankel, H. Beiträge zur fauna der Mäharichen Hohlen. Lotos 1860, 10, 201–206. [Google Scholar]
- Barranco, P.; Mayoral, J.G.; Ruiz-Portero, C.; Amate, J.; García-Pardo, J.; Piquer, M.; Ortega, M.; Salavert, V.; Ruiz Avilés, F.; Lara, M.D.; et al. Fauna endokárstica andaluza. In Investigaciones en Sistemas kársticos Españoles; Andreo, B., Durán, J.J., Eds.; Serie Hidrogeología y Aguas Subterráneas; Instituto Geológico y Minero de España: Madrid, Spain, 2004; Volume 12, pp. 351–366. ISBN 84-7840-551-8. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order and Species | Caves | |||||||
|---|---|---|---|---|---|---|---|---|
| Entomobryomorpha | CO | C3 | CA | GE | SP | TE | AP | AG |
| Entomobrya sp. | 25 | 2 | ||||||
| Lepidocyrus sp. | 3 | 4 | 24 | 1 | 12 | 7 | 3 | |
| Pseudosinella najtae | 41 | 129 | 47 | 247 | 661 | 15 | ||
| P. sexocellata sp. nov. | 254 | 2 | 297 | 12 | ||||
| Pseudosinella sp. | 2 | |||||||
| Seira sp. | 1 | |||||||
| Folsomia candida | 1 | |||||||
| Folsomia fimetaria | 1 | |||||||
| Folsomia sp. | 2 | |||||||
| Parisotoma notabilis | 80 | 3 | 5 | 3 | ||||
| Subisotoma variabilis | 1 | |||||||
| Heteromurus major | 16 | 65 | ||||||
| Heteromurus nitidus | 1 | |||||||
| Heteromurus sp. | 1 | 1 | ||||||
| Troglopedetes machadoi | 57 | 175 | 8 | 260 | 41 | 1 | 33 | |
| Tomocerus sp. | 3 | |||||||
| Neelipleona | ||||||||
| Neelus murinus | 2 | 6 | 20 | |||||
| Megalothorax sp. | 1 | |||||||
| Poduromorpha | ||||||||
| Mesogastrura lybica | 31 | |||||||
| Mesogastrura ojcoviensis | 1 | |||||||
| Symphypleona | ||||||||
| Deuterosminthurus sp. | 1 | 9 | ||||||
| Pseudosinella ruizporteroae sp. nov. | 1037 | 121 | 1949 | 332 | 1 | 716 | ||
| Pseudosinella torresi sp. nov. | 83 | 1 | 41 | |||||
| P. pygmaeus cf. | 2 | |||||||
| Pygmarrhopalites sp. | 17 | 1 | 71 | 4 | ||||
| P. subbifidus | 4 | 405 | 417 | 10 | 2 | |||
| Gisinurus malatestai | 29 | |||||||
| Sphaeridia pumilis | 6 | 2 | ||||||
| Analysis | ||||||||
| Total of individuals (per cave) | 1579 | 710 | 197 | 3073 | 6 | 1136 | 341 | 827 |
| Species number (per cave) | 11 | 4 | 6 | 15 | 3 | 12 | 9 | 12 |
| Average specimens/visit | 72 | 118 | 49 | 140 | 6 | 76 | 68 | 92 |
| Number of visits to the cave | 22 | 6 | 4 | 22 | 1 | 15 | 5 | 9 |
| An | T | S | M1 | M2 | R | E | L1 | L2 | Aa | Ab | Aq1 | Aq2 | A4 | Te | Cl | Ct | Em | Ma | Mp | Ha | Ml | D | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P. sexocellata sp. nov. | 1 | 2 | 2 | 4 | 4 | 1 | 4 | 4 | 4 | 2 | 4 | 4 | 2 | 2 | 2 | 4 | 2 | 1 | 2 | 2(3) | 1 | 1.34 | - |
| P. gutierrezae | 1 | 2 | 2 | 4 | 2 * | 0 * | 2 * | 2 * | 2 * | 2 | 4 | 4 | 1 * | 2 | 1 * | 4 | 2 | 3 * | 2 | 2 | 2 * | 1.25 | 9 |
| P. ops | 2 * | 1 * | U | 4 | 4(2) | 3 * | 4(2) | 4(3) | 4 | 0 * | 4 | 2 * | 0 * | 1 * | 2 | 3 * | 1 * | 1 | 2 | 6–8 * | 1 | 2.2 | 11 |
| P. sexoculata (USA) | 1 | 2 | 1 * | 4 | 2 * | 0 * | 2 * | 2 * | 2 * | 1 * | 3 * | 3 * | 1 * | 2 | 2 | 3 * | 1 * | 1 | 3 * | 4–10 * | 3 | 1.7 | 14 |
| P. sexoculata (Europe) | 1 | 2 | 2 | 4 | 2 * | 1 | 2 * | 2 * | 2 * | 1 * | 4 | 4 | 1 * | 2 | 2 | 3 * | 1 * | 3 * | U | U | 3 * | 1.4 | 10 |
| P. torcuatoensis | 1 | 2 | 2 | 4 | 2 * | 1 | 4 | 4 | 4 | 2 | 4 | 1 * | 1 * | 2 | 1 * | 4 | 2 | 3 * | 2 | 2 | 2 * | 1.1 | 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baquero, E.; Barranco, P.; Jordana, R. Collembola from the Gypsum Karst of Sorbas (Almería, Spain), with Descriptions of Three New Species. Insects 2025, 16, 309. https://doi.org/10.3390/insects16030309
Baquero E, Barranco P, Jordana R. Collembola from the Gypsum Karst of Sorbas (Almería, Spain), with Descriptions of Three New Species. Insects. 2025; 16(3):309. https://doi.org/10.3390/insects16030309
Chicago/Turabian StyleBaquero, Enrique, Pablo Barranco, and Rafael Jordana. 2025. "Collembola from the Gypsum Karst of Sorbas (Almería, Spain), with Descriptions of Three New Species" Insects 16, no. 3: 309. https://doi.org/10.3390/insects16030309
APA StyleBaquero, E., Barranco, P., & Jordana, R. (2025). Collembola from the Gypsum Karst of Sorbas (Almería, Spain), with Descriptions of Three New Species. Insects, 16(3), 309. https://doi.org/10.3390/insects16030309