Simple Summary
Megalurothrips usitatus (Bagnall) is one of the most important pests that harm legumes—namely alfalfa (Medicago sativa L.). The pest is harmful, has strong reproductive abilities, and can transmit plant viruses. In addition, not only does the long-term use of chemical pesticides make the pest resistant, but pesticide residues also have a negative impact on the ecological environment, meaning that control is difficult. Orius strigicollis (Poppius) is a common predatory natural enemy in the field which can prey on several kinds of pests. In our experiment, we found that the predation ability of O. strigicollis on M. usitatus strengthened with an increase in prey density, while its searching efficiency decreased with an increase in prey density. However, the intensity of natural enemy apportionment competition increased with an increase in their own density. These results provide a scientific basis for the use of O. strigicollis to control M. sativa pests and develop biological controls.
Abstract
Megalurothrips usitatus (Bagnall) is one of the most important pests harming alfalfa (Medicago sativa L.) in Ningxia. In the field, Orius strigicollis (Poppius) is abundant. It has a high search efficiency and consumption rate of prey on a wide range of thrips and is often used as a biological control agent for these insects. To understand the predatory function and biological control potential of O. strigicollis on M. usitatus, the predatory ability of adults of O. strigicollis on second-instar larvae and adults of M. usitatus was measured under indoor conditions, and the Holling II and Hassell–Varley models were fitted to calculate the predatory function response, search efficiency, intraspecific interference, and predation preference of O. strigicollis on second-instar larvae and adults of M. usitatus. Our results showed that the predatory ability of O. strigicollis on second-instar larvae of M. usitatus was 78.62, and the theoretical maximum daily predation was 76.92. The predatory ability of O. strigicollis on adults of M. usitatus was 52.79, and the theoretical maximum daily predation was 52.62. The daily predation of O. strigicollis was positively correlated with prey density and negatively correlated with search efficiency. The rate of predatory action on prey decreased with an increasing O. strigicollis density, while the intensity of apportioned competition increased with an increasing O. strigicollis density. The results of the prey preference experiment showed that the preference of O. strigicollis for second-instar larvae of M. usitatus was significantly higher than for adults of M. usitatus. In conclusion, the indoor experiment data can provide an effective reference for the use of O. strigicollis to control M. usitatus infestation, providing an important biological control agent for controlling M. sativa thrips.
1. Introduction
Alfalfa (Medicago sativa L.) has an illustrious history, stretching back through the ages [1]. It is a high-quality perennial legume forage, which is not only nutritious but also has a high protein content and high crude fiber digestibility; therefore, it is known as the “king of forages” [2,3,4]. M. sativa can also play an important role in soil improvement because of its nitrogen fixation capacity [5]. In China, the planting area of M. sativa has been expanding with the ongoing reforms in the country’s crop industry, and with that expansion, the problem of pest control has become more prominent [6]. Investigations have shown that thrips (Thysanoptera: Thripidae) are the major pests of M. sativa in the northwestern regions of China [7], while Megalurothrips usitatus (Bagnall) is a major M. sativa pest infesting M. sativa in Ningxia [8].
Megalurothrips usitatus (Bagnall) is known as bean flower thrips, Asian bean thrips, blossom thrips, or flower thrips and is widely distributed in Asia [9,10]. M. usitatus undergoes six developmental stages on the host plant: egg, first- and second-instar larva, prepupa, pupa, and adult. The second-instar larval stage is the longest and most active [11]. Tang et al. [12] reported that the lifespan of an adult M. usitatus at 26 °C ranges from 10.63 to 18.11 days, and second-instar larvae and adult thrips cause the most damage to the legume, mainly by feeding on its flowers. These studies show that the second-instar larvae and adults of M. usitatus cause the most damage to legumes. However, when the flowers of the leguminous plant are no longer available, M. usitatus adapts and starts feeding on the young leaves and pods [12,13], not only deforming and corrupting the host plant but also transmitting Tobacco Streak Virus (TSV) [14,15]. It can cause an 80–100% yield loss in legume crops in severe cases, leading to complete crop failure and significant economic losses for farmers [16]. At present, chemical insecticides are the main method for controlling thrips [17,18]; however, the long-term use of chemical pesticides has a negative impact on the environment, humans, and other living organisms by making pests resistant and upsetting the biological balance between natural enemies and pests [19,20,21,22]. In addition, M. usitatus is difficult to control, because it has characteristics such as strong concealment, rapid reproduction, and a short generation cycle and can be hidden in closed spaces to avoid chemical sprays [19,23].
The release of predatory natural enemies does not affect the environment or destroy ecological diversity, and it is one of the most important measures for integrated pest management [24]. Of these, Orius spp. (Hemiptera: Anthocoridae) are the most important group of predators that prey on thrips [25], and O. strigicollis has been commonly used as a biocontrol agent against thrips by virtue of its high search ability and consumption rate [26,27]. Wang et al. [28] found that the release of O. strigicollis in the field reduced the population density of M. usitatus. Dai et al. [29] compared O. minutus (Linnaeus), O. nagaii (Yasunaga), O. sauteri (Poppius), and O. strigicollis found that the latter has a significant potential for controlling M. usitatus. Related studies have shown that O. strigicollis can effectively control Thrips hawaiiensis (Morgan) [30], Dendrothrips minowai (Priesner) [31], Frankliniella occidentalis (Pergande) [32], Thrips palmi (Karny) [26], and other pests.
Based on the above studies, our study was conducted under indoor constant-temperature conditions, using second-instar larvae and adults of M. usitatus as prey and O. strigicollis adults as their natural enemy. By constructing models of functional responses to predation and calculating intraspecific interferences and predation preferences, it is possible to more accurately predict and understand the complex interactions between predatory natural enemies and prey. This will not only help deepen our understanding of predatory natural enemies and pest biology but also facilitate the development of more effective tools for agricultural practices to reduce M. usitatus infestation on M. sativa.
2. Research Materials and Methodology
2.1. Materials and Methodology
Insects and plants for testing: O. strigicollis and M. usitatus were collected from a M. sativa planting base in Xixia District, Yinchuan City, Ningxia Hui Autonomous Region (N38°38′59′, E106°9′6′). M. usitatus were collected and then reared in an artificial climatic chamber with a photoperiod of 14L:10D in succession with cowpeas (Vigna unguiculata). O. strigicollis was fed with M. usitatus in a plastic container in the same artificial climatic chamber. After M. usitatus had laid eggs and hatched, 2-day-old second-instar larvae and adults were selected as prey, and healthy and active adults of O. strigicollis of unknown ages were selected as the natural enemies for experiments. The individuals were starved for 24 h before the experiment [33].
2.2. The Predatory Function and Feeding Ability of O. strigicollis on Second-Instar Larvae and Adults of M. usitatus
O. strigicollis were transferred individually to plastic tubes (d = 9 cm; h = 11 cm). Then, second-instar larvae and adults of M. usitatus were placed into each test tube at different densities (10, 20, 30, 40, and 50 prey per predator, respectively). After 24 h, the number of thrips that had been eaten by O. strigicollis in each of the tubes was counted. Each density included 4 replicates.
2.3. Intraspecific Interference Experiment Determining O. strigicollis’s Predatory Activities on Adults of M. usitatus
The intraspecific interference response is a natural enemy’s own density interference response. O. strigicollis were placed in tubes at densities of 1, 2, 3, 4, and 5 individuals per tube, and the density of M. usitatus adults was set as 100 individuals per tube. The tubes were then placed in an artificial climatic chamber, with the same feeding conditions as described in Section 2.1. After 24 h, the number of M. usitatus adults that had been eaten by O. strigicollis was counted. Each treatment included 4 replicates.
2.4. Predation Preference of the O. strigicollis on Second-Instar Larvae and Adults of M. usitatus
Predation selection tests were conducted by selecting second-instar larvae and adults of M. usitatus. The second-instar of M. usitatus and adults were placed in plastic tubes (d = 9 cm; h = 11 cm) in ratios of 10:10, 20:20, and 30:30, respectively, and then, one O. strigicollis was placed in the plastic tubes. After 24 h, the number of M. usitatus larvae and adults that had been eaten by O. strigicollis was counted. Each treatment included 3 replicates.
2.5. Data Analysis
The predation function responses were fitted according to the Holling II disk equation (Holling 1959) [34]: Na = aTN/(1 + aThN). The formula for the searching efficiency of O. strigicollis on M. usitatus, S = a/(1 + aThN), was fitted [33,34,35]. In the formulas, N is the density of prey, Na is the number of predators at the corresponding density, a is the instantaneous attack rate, T is the total time of the experiment (1d in this experiment), and Th is the predator handling time for 1 prey (= T divided by the maximum predation rate).
For the intraspecific interference experiment with the O. strigicollis and M. usitatus adults, the Hassell [36] model equation was used for the fitting of the calculations to assess the disturbance effects. The formula was as follows: E = QP−m = Na/NP. Here, E is the search rate, Q is the quest constant, p is the density of predators, and m is the mutual interference constant. The method of Zou et al. [37] was used to calculate the intensity of apportioned competition, with I = (E1 − EP)/E1, where E1 is the predation rate for 1 predator, and EP is the predation rate for the predators with a density of p [37].
The predation preference of the O. strigicollis on 2nd-instar larvae and adults of M. usitatus was evaluated using the evaluation method of Zhou et al. [38]: Ci = (Qi − Fi)/(Qi + Fi), Fi = Ni/∑Ni, and Qi = Nai/∑Na. Here, Qi is the predator’s proportion of prey for the ith prey species, Fi denotes the proportion of the ith prey species among all the prey species, Ni is the number of the ith prey species in the environment, Nai is the number of the ith prey species individuals that are preyed upon by the predator, and Na is the total amount of prey taken at the corresponding density. When Ci = 0, it means that the predator has no preference for the ith prey; when 0 < Ci < 1, it means that the predator has a positive preference for the ith prey; and when −1 < Ci < 0, it means that the predator has a negative preference for the ith prey [38].
The experimental data were collated using Office 2021, and SPSS 26.0 was used for statistical analysis of the data and one-way ANOVA. Duncan’s New Complex Polar Deviation method was applied for the significance of difference test, and GraphPad Prism 9.5.0 software was used for graphing.
3. Results
3.1. Functional Responses and Search Efficiency During Predation by O. strigicollis on Second-Instar Larvae and Adults of M. usitatus
The predation behavior of O. strigicollis on M. usitatus is to suck fluids from the thrips’ body by piercing the head (Figure 1D), thorax (Figure 1A,E), and abdomen (Figure 1B). The bodies of the M. usitatus larvae and adult thrips appeared to be emptied (Figure 1C,F).

Figure 1.
Feeding process of O. strigicollis adults on second-instar larvae and adults of M. usitatus. Note: Figures (A,B): feeding on second-instar larvae of M. usitatus; (C): larva’s condition after predation; (D,E): feeding on adults; (F): adult’s condition after predation.
The predation function response model of O. strigicollis on second-instar larvae and adults of M. usitatus was fitted according to the Holling type II disk equation, with Na = 1.022N/(1 + 0.013N) and Na = 1.003N/(1 + 0.019N), respectively. The instantaneous attack rate of O. strigicollis adults on second-instar larva thrips of M. usitatus was 1.022, the handling time was 0.013, the predation capacity was 78.62, and the maximum daily predation could reach 76.92. O. strigicollis had an instantaneous attack rate of 1.003 against adults of M. usitatus, while the handling time was 0.019, the predation capacity was 52.79, and the maximum daily predation was 52.62. The correlation coefficients of the resulting equations were 0.929 and 0.921, respectively (Table 1), indicating that predation on M. usitatus by O. strigicollis was significantly correlated with the prey density. The single daily consumption and search efficiency of O. strigicollis on second-instar larvae and adults of M. usitatus are shown in Figure 2. The daily consumption of O. strigicollis adults on M. usitatus increased with prey density with a positive relationship (Figure 2A), and the search efficiency decreased with the prey density with an inverse relationship (Figure 2B). The daily predation consumption and search efficiency on second-instar larvae of M. usitatus were significantly higher than those on adults of M. usitatus (p < 0.05).

Table 1.
Functional response equations and parameters for predation of O. strigicollis on M. usitatus of different ages.
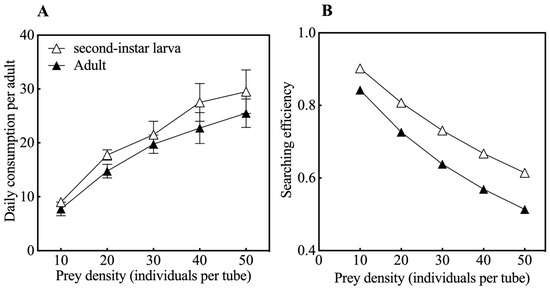
Figure 2.
Daily consumption (A) and searching efficiency (B) of O. strigicollis adults on M. usitatus.
3.2. Intraspecific Interference Experiment with O. strigicollis and Adults of M. usitatus
The densities of O. strigicollis were 1, 2, 3, 4 and 5 individuals per tube, and the density of M. usitatus adults was 100 individuals per tube. The predation amount, predation rate, and intensity of predation competition of O. strigicollis on M. usitatus were calculated for the different densities (Table 2). With a constant density and space of M. usitatus, the consumption and predation rates of O. strigicollis on M. usitatus decreased with their own density, and the intensity of apportioned competition increased with their own density, indicating mutual interference and competition among O. strigicollis individuals. The disturbance response equation for O. strigicollis predation on adults of M. usitatus was fitted with E = QP−m as E = 0.394P−0.731, with a correlation coefficient of 0.998, which can be used to describe the disturbance of O. strigicollis’s own density. Fitting the relationship between the intensity of apportioned competition (I) from O. strigicollis predation on M. usitatus and the logarithm of their own density (lgP), the resulting model was I = 1.003lgP + 0.037, with a correlation coefficient of 0.981, indicating that the intensity of apportioned competition was significantly correlated with the density of the predators.
Table 2.
Predation rate and intensity of apportioned competition of O. strigicollis adults against M. usitatus adults.
3.3. Predation Preference of O. strigicollis on Second-Instar Larvae and Adults of M. usitatus
The predation preferences of O. strigicollis on second-instar larvae and adults of M. usitatus are shown in Figure 3. O. strigicollis’s predation on second-instar larvae of M. usitatus was significantly greater than on adults in all cases (p < 0.05) (Figure 3A). When both the second-instar larvae and adults of M. usitatus were 10, 20, and 30 in number, O. strigicollis showed a preference index of Ci > 0 for second-instar larvae and −1 < Ci < 0 for M. usitatus adults (Figure 3B). This indicated that O. strigicollis showed a positive preference for second-instar larvae of M. usitatus and a negative preference for M. usitatus adults when the second-instar larvae and adults of M. usitatus coexisted in the same numbers.
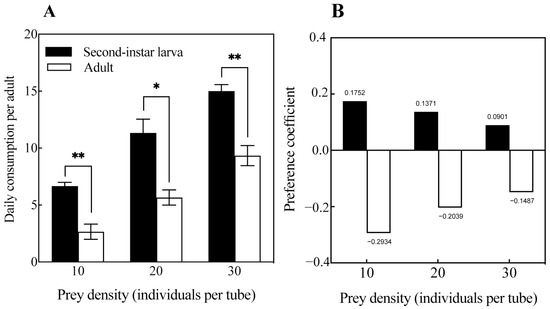
Figure 3.
Daily consumption (A) and predation coefficient (B) of O. strigicollis on second-instar larvae and adults of M. usitatus. Note: * = p < 0.05, ** = p < 0.001.
4. Discussion
The functional response is a pivotal characteristic within the intricate relationships between predator and prey and an essential component of predator–prey models. It unveils how predatory insects manage pest populations via their predatory behaviors, offering essential indicators for assessing the efficacy of biological control mechanisms. By monitoring and analyzing the ability of natural enemies to feed on pests under different environmental conditions, it is possible to accurately determine their actual role and potential impact in nature [39]. In this experiment, we recorded the predation ability and search efficiency of O. strigicollis on M. usitatus second-instar larvae and adults, as well as the intraspecific disturbance response of O. strigicollis. These observations not only reveal the behavioral adaptability of predators in hunting behavior, more importantly, they demonstrate how predators adapt to complex ecosystems by adjusting their own behavior. The search patterns of insects can respond to external and internal environmental factors, and have a high degree of plasticity, which is of great significance for understanding ecological balance and biodiversity. Through further research, we can gain a deeper understanding of the predation mechanisms of natural enemies and their roles in nature [40]. Additionally, the dried-out appearance of the M. usitatus bodies post-predation serves as a visual indication of the effectiveness of O. strigicollis’s feeding mechanism, highlighting the precision and efficiency of its predatory behavior. This is consistent with the predatory behavior of Orius sauter on Dendrothrips minowai [41].
The Holling II functional is considered the most common predatory functional response class in arthropods, characterized by a hyperbolics curve. Starting from low prey density on the abscissa, the predation rate increases almost linearly. It is generally accepted for use in evaluation systems for biological insect control and has strong applicability [39,42]. In our experiments, the predatory function response indicated that the predation ability and maximum predation amount of O. strigicollis were higher on second-instar larvae than on adults of M. usitatus. The predation of natural enemies increased with the prey density, while the search efficiency decreased with the prey density. These results were consistent with the predation function response that we calculated using Holling type II and similar to the predatory functional response of O. sauteri and O. tantillus on M. usitatus [33,43]. This shows that O. strigicollis has the potential to rapidly control M. usitatus, in addition, it exhibits greater effectiveness in managing pests in their young stages. Therefore, comprehensive pest management targeting different stages of life is crucial [44].
When the density of O. strigicollis itself increases, the intensity of competition becomes more significant, which means that O. strigicollis needs to compete with other individuals for limited resources. Thus, there are intensifying interactions and competitive relationships among individuals. The inter-individual interaction significantly increases the level of conflict among natural enemies, which in turn reduces the efficiency of predation [45]. But to a certain extent, it enhances the adaptive ability of individuals in the population to adverse environmental conditions, and O. strigicollis can adapt to environmental changes by regulating its own density in order to maintain the survival and reproduction of its population. This process also provides us with an important perspective to understand the behavioral ecology of insects, whose biodiversity is not always static and unchanging, but can be affected by a variety of factors such as environmental factors and the density of individuals [46,47].
The predation preference results showed that O. strigicollis had a positive preference for M. usitatus second-instar larvae and a negative preference for M. usitatus adults when the second-instar larvae and adults coexisted at the same density. This was similar to the results regarding the preference of O. strigicollis for F. occidentalis larvae and adults and indicates the complexity of the interaction between natural enemies and prey at different growth stages [48]. With age, prey grow in size and defense compared to their larval stage, making it more difficult for natural enemies to feed on them, which means that natural enemies have to spend more time capturing and feeding on the prey. Insects do not “keenly sense” environmental changes in the same way higher animals do. Therefore, predators have developed a complex set of adaptive mechanisms over a long period of co-evolution [49]. Physiological differences in the different age stages of M. usitatus may provide different nutritional values for O. strigicollis in order to obtain the maximum energy, and the ease of predation on larvae enables O. strigicollis to conserve its stamina, which in turn affects its predation strategy [50,51].
In conclusion, studying the functional response of O. strigicollis to predation by M. usitatus will not only deepen our understanding of the relationship between M. usitatus and its natural enemies but will also provide important data support for pest management. Understanding the inter-relationships among these organisms is essential for the development of effective pest management strategies while preserving the ecological balance [52] so that appropriate measures can be taken to protect natural enemies, control pests, and protect plants from insect pests.
Author Contributions
Conceptualization, S.W. and R.Z.; data curation, S.W., Z.F., Y.C. (Yuanrun Cheng), Y.C. (Yifan Cui) and C.X.; formal analysis, Z.F. and Z.C.; funding acquisition, S.W.; investigation, S.W., Z.F., Y.C. (Yuanrun Cheng), Y.C. (Yifan Cui) and C.X.; methodology, S.W., Y.T. and Z.F.; project administration, S.W., C.L., Y.T. and Y.W.; resources, S.W., Y.T. and Z.F.; software, Z.F., Z.C. and Y.C. (Yifan Cui); supervision, S.W., Y.T., W.S., L.B., Y.W. and R.Z.; validation, Z.F., Z.C., C.X. and Y.W.; visualization, Z.F. and C.X.; writing—original draft, Z.F. and S.W.; writing—review and editing, S.W., Y.T. and Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
The Ningxia Province Sci-Tech Innovation Demonstration Program of High-Quality Agricultural Development and Ecological Conservation (NGSB-2021-15-04), the Key Project of the Ningxia Natural Science Foundation (2024AAC02065), and China Agriculture Research System (CARS-34) funded this research.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Michaud, R.; Lehman, W.F.; Rumbaugh, M. World distribution and historical development. Alfalfa Alfalfa Improv. 1988, 29, 25–91. [Google Scholar]
- Acharya, J.P.; Lopez, Y.; Gouveia, B.T.; de Bem Oliveira, I.; Resende, M.F.R., Jr.; Muñoz, P.R.; Rios, E.F. Breeding alfalfa (Medicago sativa L.) adapted to subtropical agroecosystems. Agronomy 2020, 10, 742. [Google Scholar] [CrossRef]
- Marković, J.; Štrbanović, R.; Cvetković, M.; Anđelković, B.; Živković, B. Effects of growth stage on the mineral concentrations in alfalfa (Medicago sativa L.) leaf, stem and the whole plant. Biotechnol. Anim. Husb. 2009, 25, 1225–1231. [Google Scholar]
- Feng, Y.; Shi, Y.; Zhao, M.; Shen, H.; Xu, L.; Luo, Y.; Liu, Y.; Xing, A.; Kang, J.; Jing, H.; et al. Yield and quality properties of alfalfa (Medicago sativa L.) and their influencing factors in China. Eur. J. Agron. 2022, 141, 126637. [Google Scholar] [CrossRef]
- Huang, Z.; Sun, L.; Liu, Y.; Liu, Y.F.; López-Vicente, M.; Wei, X.H.; Wu, G.L. Alfalfa planting significantly improved alpine soil water infiltrability in the Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2019, 285, 106606. [Google Scholar] [CrossRef]
- Fan, P.A.; Gao, L.J.; Zhu, K.H.; Du, G.L.; Zhu, M.M.; Li, Z.H.; Gao, Y.L.; Tu, X.B.; Zhang, Z.H. Regional selection of insecticides and fungal biopesticides to control aphids and thrips and improve the forage quality of alfalfa crops. J. Integr. Agric. 2023, 22, 185–194. [Google Scholar]
- Wu, F.; Shi, S.; Li, Y.; Miao, J.; Kang, W.; Zhang, J.; Yun, A.; Liu, C. Physiological and biochemical response of different resistant alfalfa cultivars against thrips damage. Physiol. Mol. Biol. Plants 2021, 27, 649–663. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.; Xu, Y.; Zhang, H.; Wang, Y.; Zhang, R.; Wei, S.; Ban, L. Dynamic and correlation of major pests with natural enemies and response to climatic factors in alfalfa fields in Ningxia. J. Plant Prot. 2024, 51, 1169–1178. [Google Scholar]
- Mound, L.A.; Walker, A.K. Thysanoptera as tropical tramps: New records from New Zealand and the Pacific. N. Z. Entomol. 1987, 9, 70–85. [Google Scholar] [CrossRef]
- Palmer, J. Magalurothrips in the flowers of tropical legumes: A morphometric study. In Population Structure, Genetics and Taxonomy of Aphids and Thysanoptera; SPB Academic Publishing: Amsterdam, The Netherlands, 1987; pp. 480–495. [Google Scholar]
- Ke, T. Effect of Host Plants and Adults Sex Ratios on the Population of Megalurothrips usitatus. Ph.D. Thesis, Hainan University, Haikou, China, 2017. [Google Scholar]
- Tang, L.D.; Yan, K.L.; Fu, B.-L.; Wu, J.-H.; Liu, K.; Lu, Y.-Y. The life table parameters of Megalurothrips usitatus (Thysanoptera: Thripidae) on four leguminous crops. Fla. Entomol. 2015, 98, 620–625. [Google Scholar] [CrossRef]
- Duff, J.D.; Church, C.E.; Healey, M.A.; Senior, L. Thrips incidence in green beans and the degree of damage caused. In Proceedings of the XXIX International Horticultural Congress on Horticulture: Sustaining Lives, Livelihoods and Landscapes (IHC2014): 1105, Brisbane, QLD, Australia, 17–22 August 2014; pp. 19–26. [Google Scholar]
- Prasada Rao, R.D.; Reddy, A.S.; Reddy, S.V.; Thirumala-Devi, K.; Rao, S.C.; Manoj Kumar, V.; Subramaniam, K.; Yellamanda Reddy, T.; Nigam, S.N.; Reddy, D.V. The host range of Tobacco streak virus in India and transmission by thrips. Ann. Appl. Biol. 2003, 142, 365–368. [Google Scholar] [CrossRef]
- Shukla, S.; Kalyani, G.; Kulkarni, N.; Waliyar, F.; Nigam, S.N. Mechanism of transmission of Tobacco streak virus by Scirtothrips dorsalis, Frankliniella schultzei and Megalurothrips usitatus in groundnut, Arachis hypogaea L. J. Oilseeds Res. 2005, 22, 215–217. [Google Scholar]
- Oparaeke, A.M. The sensitivity of flower bud thrips, Megalurothrips sjostedti Trybom (Thysanoptera: Thripidae), on cowpea to three concentrations and spraying schedules of Piper guineense Schum. & Thonn. extracts. Plant Prot. Sci. 2006, 42, 106. [Google Scholar]
- Camara, I.; Cao, K.; Sangbaramou, R.; Wu, P.; Shi, W.; Tan, S. Screening of Beauveria bassiana (Bals.) (Hypocreales: Cordycipitaceae) strains against Megalurothrips usitatus (Bagnall) (Thysanoptera: Thripidae) and conditions for large-scale production. Egypt. J. Biol. Pest Control 2022, 32, 85. [Google Scholar] [CrossRef]
- Wu, J.H.; Yang, B.; Zhang, X.C.; Cuthbertson, A.G.S.; Ali, S. Synergistic Interaction between the Entomopathogenic Fungus Akanthomyces attenuatus (Zare & Gams) and the Botanical Insecticide Matrine against Megalurothrips usitatus (Bagrall). J. Fungi 2021, 7, 536. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yang, B.; Li, Z.; Yue, Y.; Tian, Q.; Chen, W.; Ali, S.; Wu, J. Immune-Related Genes of Megalurothrips usitatus (Bagrall) Against Beauveria brongniartii and Akanthomyces attenuatus Identified Using RNA Sequencing. Front. Physiol. 2021, 12, 671599. [Google Scholar] [CrossRef] [PubMed]
- Weisenburger, D.D. Human health effects of agrichemical use. Hum. Pathol. 1993, 24, 571–576. [Google Scholar] [CrossRef]
- Tang, L.D.; Guo, L.H.; Ali, A.; Desneux, N.; Zang, L.S. Synergism of adjuvants mixed with spinetoram for the management of bean flower thrips, Megalurothrips usitatus (Thysanoptera: Thripidae) in cowpeas. J. Econ. Entomol. 2022, 115, 2013–2019. [Google Scholar] [CrossRef]
- Yura, W.F.; Muhammad, F.R.; Mirza, F.F.; Maurend, Y.L.; Widyantoro, W.; Farida, S.S.; Aziz, Y.P.; Desti, A.; Edy, W.; Septy, M.; et al. Pesticide residues in food and potential risk of health problems: A systematic literature review. IOP Conf. Ser. Earth Environ. Sci. 2021, 894, 012025. [Google Scholar] [CrossRef]
- Liu, P.; Qin, Z.; Feng, M.; Zhang, L.; Huang, X.; Shi, W. The male-produced aggregation pheromone of the bean flower thrips Megalurothrips usitatus in China: Identification and attraction of conspecifics in the laboratory and field. Pest Manag. Sci. 2020, 76, 2986–2993. [Google Scholar] [CrossRef]
- Hajek, A.E.; Eilenberg, J. Natural Enemies: An Introduction to Biological Control; Cambridge University Press: Cambridge, UK, 2018. [Google Scholar]
- Khan, R.; Seal, D.; Adhikari, R. Bean Flower Thrips Megalurothrips usitatus (Bagnall) (Insecta: Thysanoptera: Thripidae); EDIS: Gainesville, FL, USA, 2022. [Google Scholar]
- Kim, D.I.; Park, J.D.; Kim, S.G.; Kim, S.S.; Paik, C.H. Biological control of Thrips palmi (Thysanoptera: Thripidae) with Orius strigicollis (Hemiptera: Anthocoridae) on cucumber in plastic houses in the southern region of Korea. J. Asia Pac. Entomol. 2004, 7, 311–315. [Google Scholar] [CrossRef]
- Tuan, S.J.; Lin, Y.H.; Peng, S.C.; Lai, W.H. Predatory efficacy of Orius strigicollis (Hemiptera: Anthocoridae) against Tetranychus urticae (Acarina: Tetranychidae) on strawberry. J. Asia-Pac. Entomol. 2016, 19, 109–114. [Google Scholar] [CrossRef]
- Wang, C.; Lee, P.; Wu, Y. Field augmentation of Orius strigicollis (Heteroptera: Anthocoridae) for the control of thrips in Taiwan. In International Seminar on Biological Control of Insect Pests in Economic Crops; FFTC: Taipei City, Taiwan, 2001; Volume 500, pp. 141–152. [Google Scholar]
- Dai, X.; Wang, R.; Liu, Y.; Su, L.; Yin, Z.; Wu, M.; Chen, H.; Zheng, L.; Zhai, Y. Control effect and field application of four predatory Orius species on Megalurothrips usitatus (Thysanoptera: Thripidae). J. Econ. Entomol. 2024, 117, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Zeng, Z.; Chen, Y.; You, Y.; Hu, J.; Yang, F.; Wei, H. Compatibility of six reduced-risk insecticides with Orius strigicollis (Heteroptera: Anthocoridae) predators for controlling Thrips hawaiiensis (Thysanoptera: Thripidae) pests. Ecotoxicol. Environ. Saf. 2021, 226, 112812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.R.; Liu, M.; Ban, F.X.; Shang, X.L.; Liu, S.L.; Mao, T.T.; Zhang, X.Y.; Zhi, J.R. Establishment of a faba bean banker plant system with predator Orius strigicollis for the control of thrips Dendrothrips minowai on tea plants under laboratory conditions. Insects 2021, 12, 397. [Google Scholar] [CrossRef] [PubMed]
- Ohta, I. Effect of temperature on development of Orius strigicollis (Heteroptera: Anthocoridae) fed on Frankliniella occidentalis (Thysanoptera: Thripidae). Appl. Entomol. Zool. 2001, 36, 483–488. [Google Scholar] [CrossRef][Green Version]
- Liu, P.; Jia, W.; Zheng, X.; Zhang, L.; Sangbaramou, R.; Tan, S.; Liu, Y.; Shi, W. Predation functional response and life table parameters of Orius sauteri (Hemiptera: Anthocoridae) feeding on Megalurothrips usitatus (Thysanoptera: Thripidae). Fla. Entomol. 2018, 101, 254–259. [Google Scholar] [CrossRef]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Ding, Y.Q. Insect Mathematical Ecology; Science Press: Beijing, China, 1994; pp. 257–258, 303–304. [Google Scholar]
- Hassell, M.P.; Varley, G.C. New inductive population model for insect parasites and its bearing on biological control. Nature 1969, 223, 1133–1137. [Google Scholar] [CrossRef]
- Zou, Y.; Geng, J.; Chen, G.; Meng, Q.; Wang, G. Predation of Harmonia axyridis nymph on Schizaphis graminum. Chin. J. Appl. Ecol. 1996, 7, 197–200. [Google Scholar]
- Zhou, J.Z.; Chen, C.M. Quantitative measurement of selectivityof predator for prey. Acta Ecol. Sin. 1987, 7, 50–56. [Google Scholar]
- Jeschke, J.M.; Kopp, M.; Tollrian, R. Predator functional responses: Discriminating between handling and digesting prey. Ecol. Monogr. 2002, 72, 95–112. [Google Scholar] [CrossRef]
- Bell, W.J. Searching behavior patterns in insects. Annu. Rev. Entomol. 1990, 35, 447–467. [Google Scholar] [CrossRef]
- Zhang, F.G.; Cai, X.M.; Xiu, C.L.; Wang, G.C.; Chen, Z.M. Predatory function of mulberry flower bug Orius sauteri on tea stick thrips Dendrothrips minowai adults. J. Plant Prot. 2023, 50, 668–675. [Google Scholar]
- Hassell, M.P.; Lawton, J.H.; Beddington, J.R. The components of arthropod predation: I. The prey death rate. J. Anim. Ecol. 1976, 45, 135–164. [Google Scholar] [CrossRef]
- Qiu, H.; Fu, B.; He, S.; Liu, K. Functional response and predation preference of Orius tantillus to Megalurothrips usitatus. Chin. J. Biol. Control 2022, 38, 1443. [Google Scholar]
- Luo, Z.W.; Gong, X.N.; Yu, X.S.; Wang, X.S.; Shen, S.Q.; Long, Y.Q. Evaluation of predation effectiveness of Arma chinensis to Agriophara rhombata larvae. For. Pest Dis. 2023, 42, 19–25. [Google Scholar]
- Wang, L.P.; Wang, Y.M.; Du, J.P.; Zhang, G.A. Predation of amblyseius swirskii on tetranychus cinnabarinus nymphs. Chin. J. Biol. Control 2011, 27, 171–175. [Google Scholar]
- Osawa, N. Sex-dependent effects of sibling cannibalism on life history traits of the ladybird beetle Harmonia axyridis. Biol. J. Linn. Soc. 2002, 76, 349–360. [Google Scholar] [CrossRef]
- Getto, P.; Diekmann, O.; de Roos, A.M. On the (dis) advantages of cannibalism. J. Math. Biol. 2005, 51, 695–712. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.X.; Lei, Z.Y.; Zhang, Q.; Fan, W.; Yin, H.H.; Xu, T.Y.; Chen, G.H.; Zhang, X.M. Predatory function of minute pirate bug Orius strigicollis against western flower thrips Frankliniella occidentalis adults and its intraspecific cannibalism behavior. J. Plant Prot. 2022, 49, 758–766. [Google Scholar]
- Schmitz, O.J. Effects of predator hunting mode on grassland ecosystem function. Science 2008, 319, 952–954. [Google Scholar] [CrossRef] [PubMed]
- Wiegert, R.G.; Petersen, C.E. Energy transfer in insects. Annu. Entomol. 1983, 28, 455–486. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Deng, B.D.; Zhang, W.J.; Ma, G.L.; Gao, G.Z. Predation of Harmonia axyridis (Coleoptera: Coccinellidae) on two walnut aphids, Chromaphis juglandicola and Panaphis juglandis. J. Appl. Entomol. 2024, 61, 827–834. [Google Scholar]
- Chailleux, A.; Mohl, E.K.; Teixeira Alves, M.; Messelink, G.J.; Desneux, N. Natural enemy-mediated indirect interactions among prey species: Potential for enhancing biocontrol services in agroecosystems. Pest Manag. Sci. 2014, 70, 1769–1779. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).