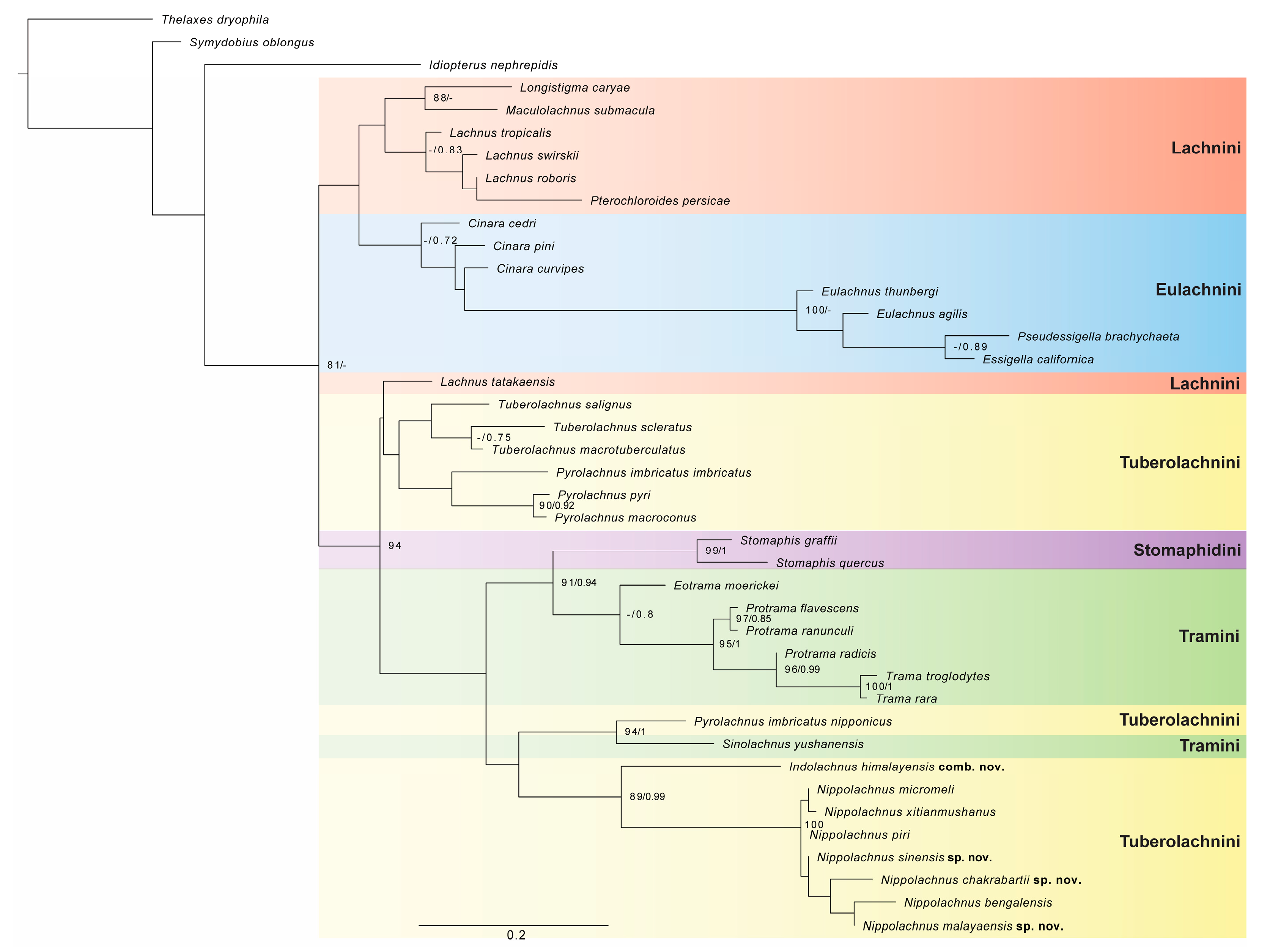
3.3.6. Review of Species of the Genus Nippolachnus
Nippolachnus bengalensis Basu and Hille Ris Lambers, 1968
=
Nippolachnus benzalensis Basu and Hille Ris Lambers [
20], p. 9
=
N. benzalensis Ghosh AK, 1974 [
29]
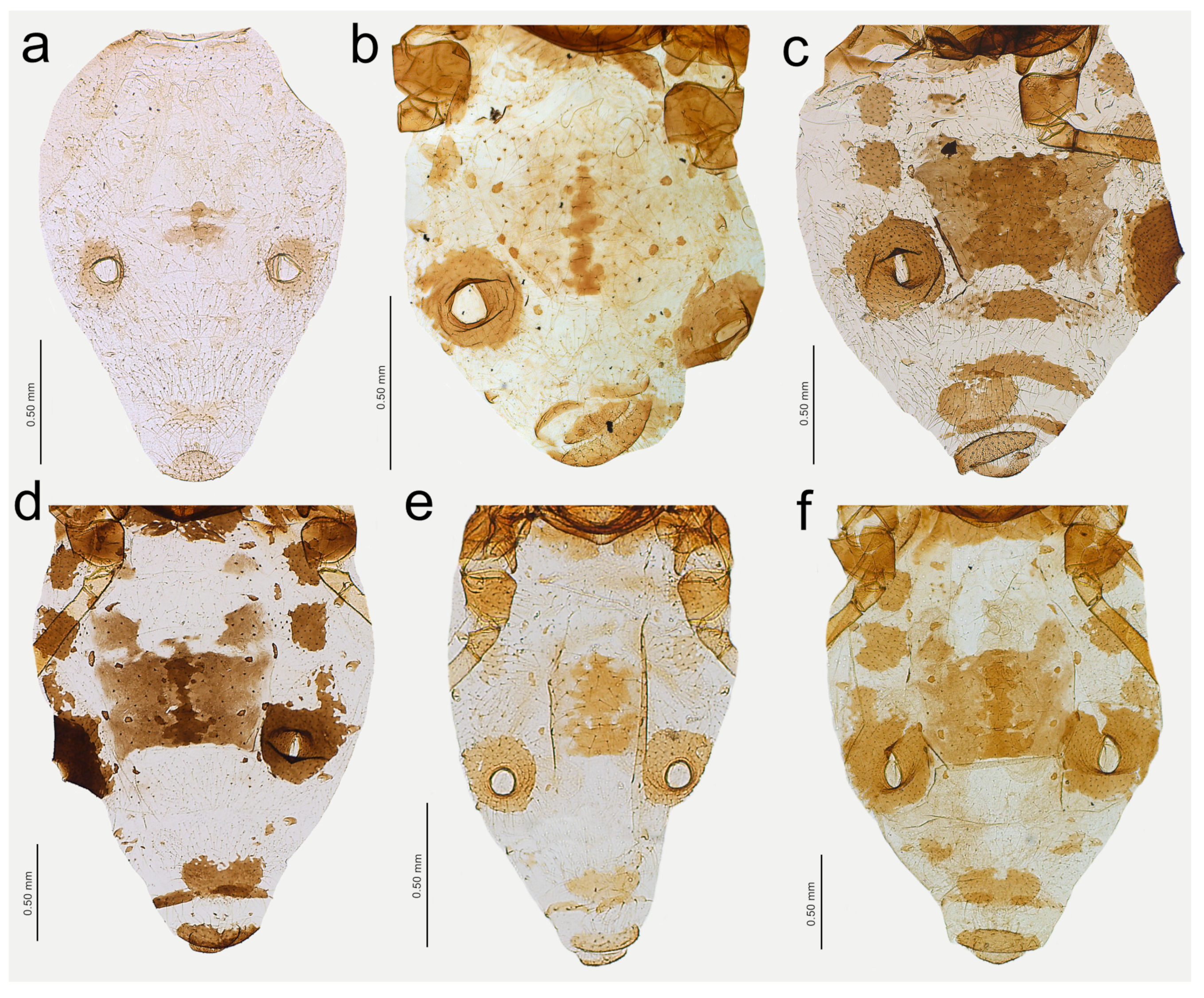
Figure 13.
Apterous viviparous females of Nippolachnus and Indolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. malayaensis sp. nov., (d) N. micromeli, (e) N. piri, (f) N. sinensis sp. nov., (g) N. xitianmushanus, (h) I. himalayensis.
Figure 13.
Apterous viviparous females of Nippolachnus and Indolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. malayaensis sp. nov., (d) N. micromeli, (e) N. piri, (f) N. sinensis sp. nov., (g) N. xitianmushanus, (h) I. himalayensis.
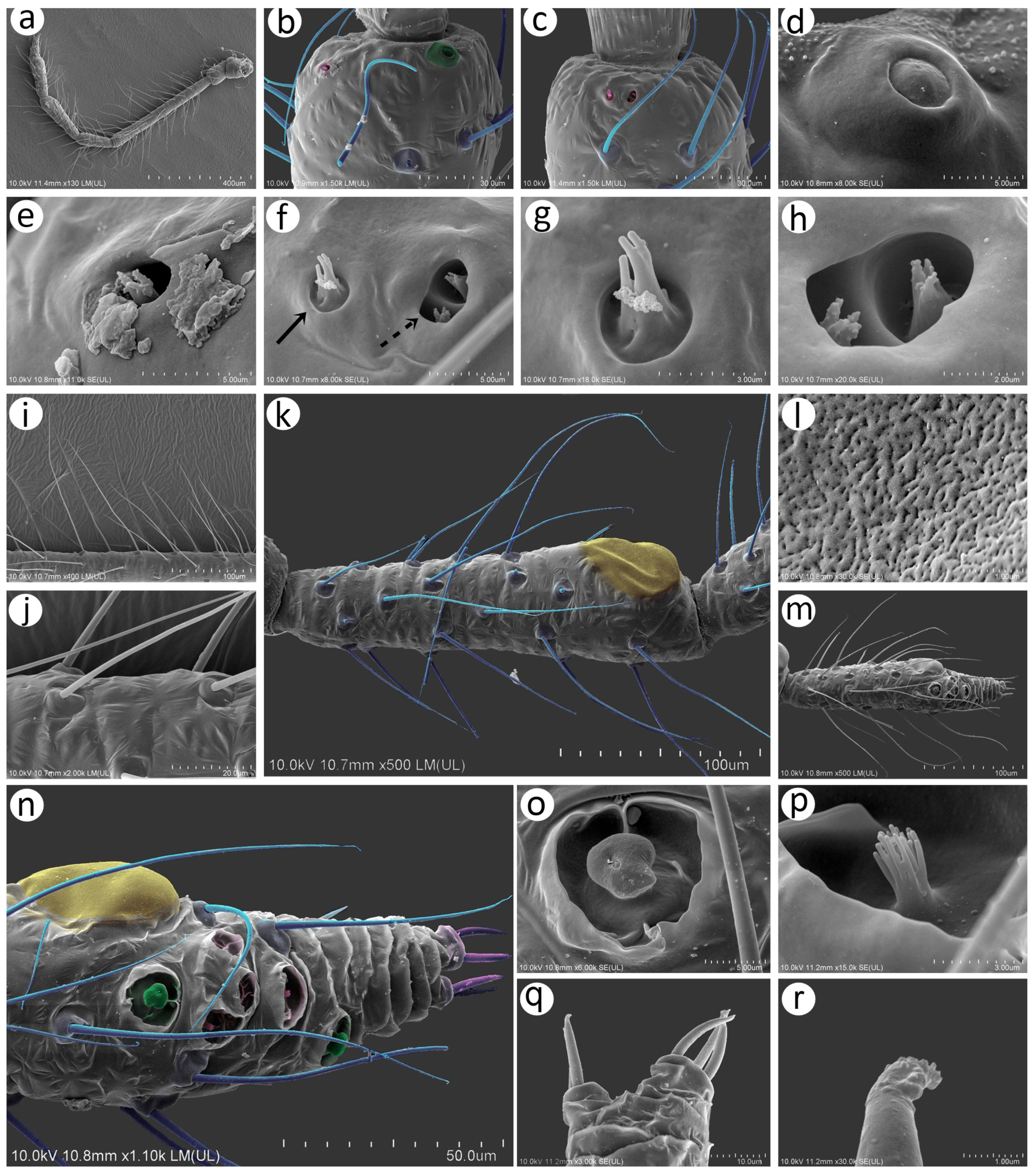
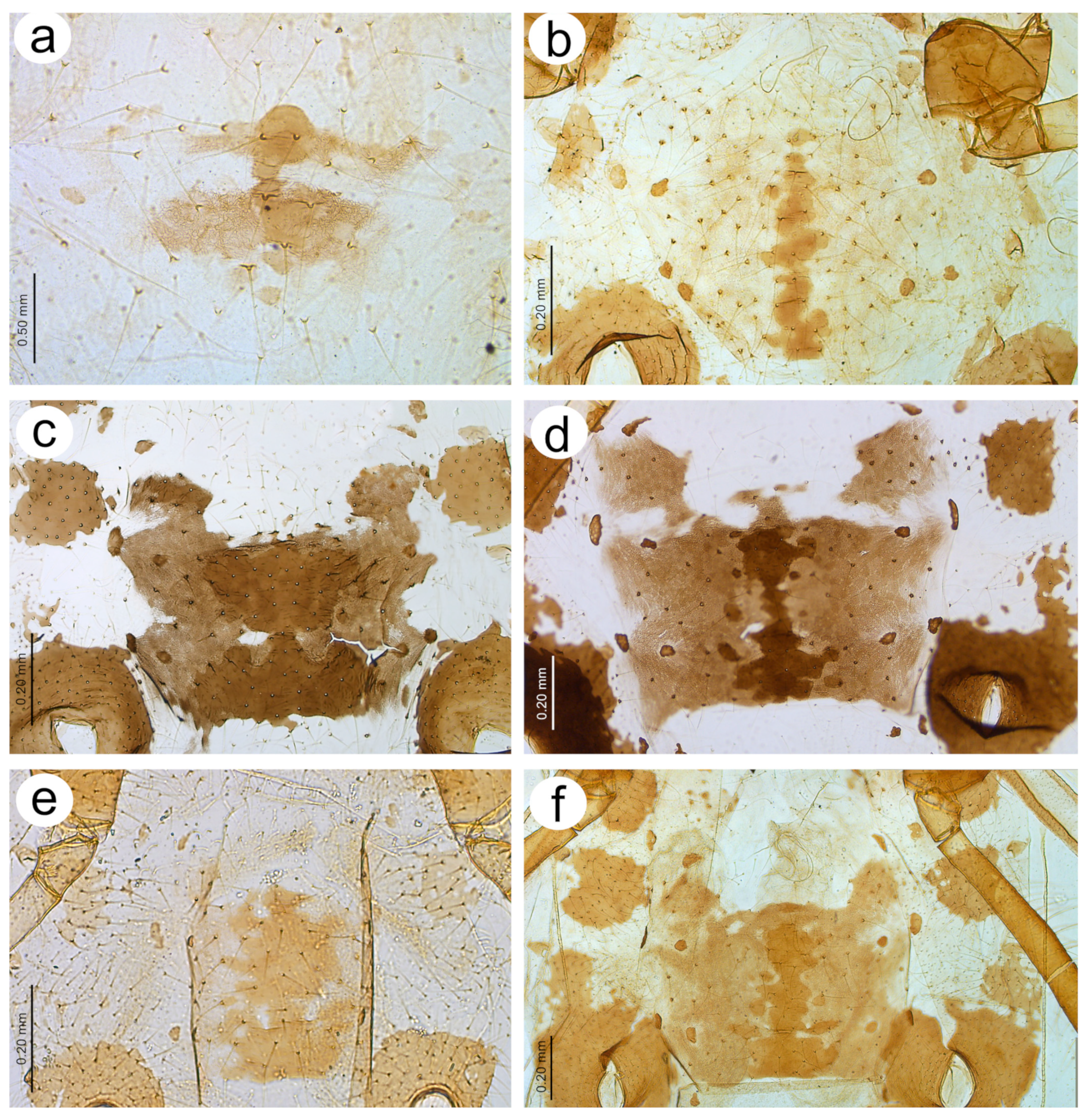
Figure 14.
Last antennal segment of apterous viviparous females: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. malayaensis sp. nov., (d) N. micromeli, (e) N. piri, (f) N. sinensis sp. nov., (g) N. xitianmushanus, (h) I. himalayensis; white dotted line indicates the major rhinarium, red dotted line indicates the position of accessory rhinaria relative to the major rhinarium.
Figure 14.
Last antennal segment of apterous viviparous females: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. malayaensis sp. nov., (d) N. micromeli, (e) N. piri, (f) N. sinensis sp. nov., (g) N. xitianmushanus, (h) I. himalayensis; white dotted line indicates the major rhinarium, red dotted line indicates the position of accessory rhinaria relative to the major rhinarium.
Figure 15.
Hind leg pigmentation of apterous viviparous females: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. malayaensis sp. nov., (d) N. micromeli, (e) N. piri, (f) N. sinensis sp. nov., (g) N. xitianmushanus, (h) I. himalayensis.
Figure 15.
Hind leg pigmentation of apterous viviparous females: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. malayaensis sp. nov., (d) N. micromeli, (e) N. piri, (f) N. sinensis sp. nov., (g) N. xitianmushanus, (h) I. himalayensis.
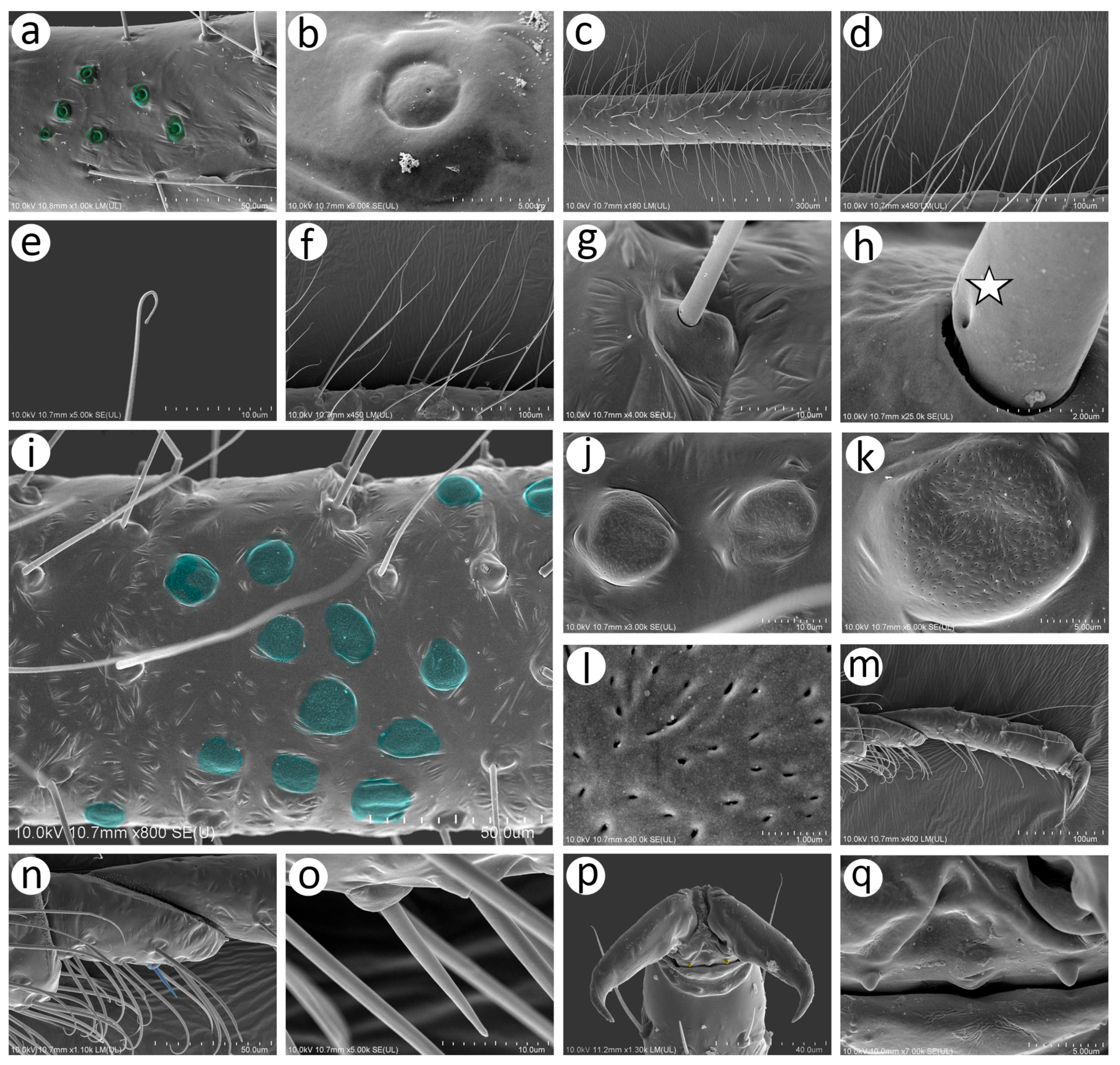
Figure 16.
Alate viviparous females of Nippolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus.
Figure 16.
Alate viviparous females of Nippolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus.
Figure 17.
Antennae of alate viviparous females of Nippolachnus and Indolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus, (g) I. himalayensis (after van der Goot, redrawn).
Figure 17.
Antennae of alate viviparous females of Nippolachnus and Indolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus, (g) I. himalayensis (after van der Goot, redrawn).
Figure 18.
Fore wing venation and scale-like element distribution in alate viviparous females (dark field microscopy): (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus, (g) I. himalayensis (after van der Goot, redrawn).
Figure 18.
Fore wing venation and scale-like element distribution in alate viviparous females (dark field microscopy): (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus, (g) I. himalayensis (after van der Goot, redrawn).
Table 1.
Measurements (in mm) of apterous viviparous females of Nippolachnus and Indolachnus.
Table 1.
Measurements (in mm) of apterous viviparous females of Nippolachnus and Indolachnus.
| Character | Nippolachnus | Indolachnus
himalayensis |
|---|
| bengalensis | chakrabartii | malayaensis | micromeli | piri | sinensis | xitianmushanus |
|---|
| BL | 2.96–3.62 | 2.65–3.32 | 2.45–2.85 | 2.25–2.27 | 2.27–3.55 | 2.35–2.85 | 3.35 | 3.45–4.72 |
| HW | 0.45–0.50 | 0.50–0.60 | 0.44–0.45 | 0.44–0.50 | 0.50–0.59 | 0.50–0.54 | 0.55 | 0.65–0.85 |
| ANT | 0.91–1.03 | 0.77–0.89 | 0.92–1.00 | 0.75–0.84 | 0.90–1.03 | 0.79–0.86 | 0.85–0.87 | 1.44–1.51 |
| ANT III | 0.33–0.40 | 0.25–0.27 | 0.30–0.33 | 0.24–0.29 | 0.30–0.34 | 0.27–0.29 | 0.28–0.30 | 0.53–0.58 |
| ANT IV | 0.13–0.15 | 0.10–0.12 | 0.11 | 0.07–0.10 | 0.10–0.14 | 0.08–0.10 | 0.11 | 0.20–0.22 |
| ANT V | 0.16–0.18 | 0.15–0.16 | 0.16–0.18 | 0.13–0.15 | 0.16–0.19 | 0.15 | 0.12–0.15 | 0.25–0.27 |
| ANT VI | 0.14–0.16 | 0.14–0.17 | 0.21–0.26 | 0.14–0.18 | 0.17–0.20 | 0.15–0.18 | 0.185 | 0.22–0.23 |
| BASE | 0.11–0.12 | 0.10–0.11 | 0.13–0.18 | 0.09–0.12 | 0.11–0.13 | 0.10–0.12 | 0.11–0.12 | 0.15 |
| PT | 0.03–0.04 | 0.04–0.06 | 0.08–0.09 | 0.04–0.06 | 0.05–0.07 | 0.05–0.06 | 0.06–0.07 | 0.07–0.08 |
| URS | 0.14–0.15 | 0.16–0.18 | 0.13–0.14 | 0.13–0.15 | 0.17–0.20 | 0.15 | 0.18 | 0.22–0.23 |
| III FEMORA | 1.42–1.45 | 1.12–1.35 | 1.07–1.17 | 0.90–1.12 | 1.20–1.60 | 0.98–1.27 | 1.37–1.45 | 1.08–2.05 |
| III TIBIAE | 2.72–2.75 | 2.15–2.50 | 2.18–2.47 | 1.70–2.15 | 2.37–2.82 | 2.00–2.37 | 2.50–2.62 | 2.87–3.30 |
| HT II | 0.22–0.23 | 0.20–0.22 | 0.20–0.22 | 0.19–0.22 | 0.22–0.25 | 0.20–0.21 | 0.22–0.23 | 0.35–0.36 |
Table 2.
Measurements (in mm) of alate viviparous females of Nippolachnus and Indolachnus. Available measurements of I. himalayensis after Basu and Hille Ris Lambers (1968).
Table 2.
Measurements (in mm) of alate viviparous females of Nippolachnus and Indolachnus. Available measurements of I. himalayensis after Basu and Hille Ris Lambers (1968).
| Character | Nippolachnus | Indolachnus
himalayensis |
|---|
| bengalensis | chakrabartii | micromeli | piri | sinensis | xitianmushanus |
|---|
| BL | 2.99–3.20 | 1.91–2.77 | 2.87–3.25 | 2.90–4.05 | 2.62–2.82 | 3.12–3.95 | 3.95–4.01 |
| HW | 0.50 | 0.57–0.58 | 0.56–0.64 | 0.55–0.70 | 0.55–0.62 | 0.66–0.78 | - |
| ANT | 0.90–0.96 | 0.83–0.84 | 0.98–1.04 | 0.92–1.16 | 0.82–0.87 | 0.95–1.08 | 1.57 |
| ANT III | 0.32–0.35 | 0.25–0.27 | 0.32–0.35 | 0.31–0.39 | 0.26–0.28 | 0.30–0.35 | 0.57–0.67 |
| ANT IV | 0.13–0.14 | 0.10–0.11 | 0.12–0.14 | 0.11–0.15 | 0.09–0.10 | 0.13–0.16 | 0.24–0.27 |
| ANT V | 0.15–0.17 | 0.16–0.17 | 0.18–0.21 | 0.16–0.19 | 0.15–0.16 | 0.19–0.20 | 0.26–0.27 |
| ANT VI | 0.14–0.15 | 0.16–0.17 | 0.19–0.20 | 0.18–0.23 | 0.18–0.19 | 0.18–0.23 | 0.24 |
| BASE | 0.11–0.12 | 0.10–0.11 | 0.13 | 0.12–0.16 | 0.12–0.13 | 0.12–0.15 | 0.17 |
| PT | 0.03–0.04 | 0.055 | 0.06–0.07 | 0.06–0.07 | 0.06 | 0.06–0.08 | 0.07 |
| URS | 0.17–0.19 | 0.15–0.16 | 0.15–0.17 | 0.17–0.19 | 0.15–0.17 | 0.19–0.21 | 0.17–0.19 |
| III FEMORA | 1.35–0.37 | 1.20 | 1.37–1.60 | 1.37–1.75 | 1.17–1.32 | 1.40–1.70 | - |
| III TIBIAE | 2.65–2.72 | 2.35 | 2.50–2.82 | 2.60–3.30 | 2.30–2.50 | 2.50–3.05 | - |
| HT II | 0.22–0.23 | 0.20–0.21 | 0.15–0.17 | 0.21–0.25 | 0.21–0.22 | 0.23–0.24 | - |
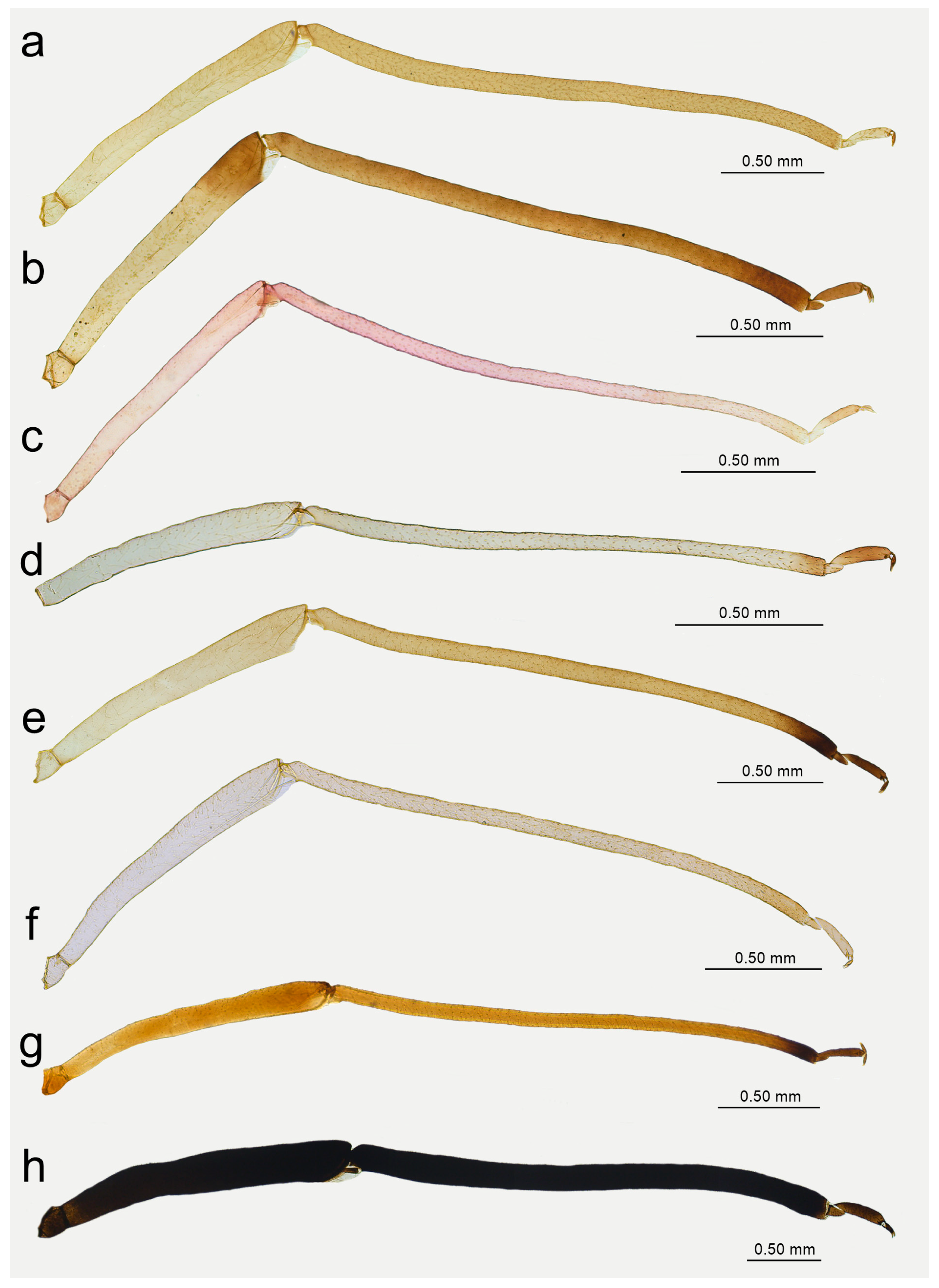
Figure 19.
Abdomen sclerotisation of alate viviparous females of Nippolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus.
Figure 19.
Abdomen sclerotisation of alate viviparous females of Nippolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus.
Figure 20.
Characters of the central abdominal sclerotisation of alate viviparous females of Nippolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus.
Figure 20.
Characters of the central abdominal sclerotisation of alate viviparous females of Nippolachnus: (a) N. bengalensis, (b) N. chakrabartii sp. nov., (c) N. micromeli, (d) N. piri, (e) N. sinensis sp. nov., (f) N. xitianmushanus.
Colour: in life, light green [
20]. Mounted specimens: body in general pale. ANT pale with yellowish ANT VI. Fore and middle legs pale yellow. Uniformly yellow (slightly darker than fore and middle legs) with paler tarsi (
Figure 13a and
Figure 15a).
Morphometric characters: HW 0.48 × ANT. ANT 0.27–0.32 × BL. ANT III with 0–2 (rarely) secondary rhinaria, ANT IV with 0–3 (rarely). ANT V longer than ANT VI with 0–2 (rarely) secondary rhinaria. ANT VI with 14–15 basal setae and without long, fine setae on PT (
Figure 14a). ANT VI PT 0.27–0.32 × BASE. Other antennal ratios: VI:III 0.37–0.42, V:III 0.45–0.48, IV:III 0.35–0.41. LS III 5.00–6.00 × BD III. URS 0.37–0.40 × ANT III, 0.90–0.96 × ANT VI, and 0.64–0.65 × HT II with 8–10 accessory setae. Hind tibiae setae 0.08–0.18 mm long. HT I with 7–9 ventral setae (except sense peg). HT II 0.57–0.62 × ANT III and 1.39–1.50 ANT VI. Dorsal setae, 0.11–0.13 mm long. ABD VIII with 40–42 setae.
Colour: in life, head and thorax greenish brown, abdomen pale green [
20]. Mounted specimens: antennae yellow with darker apical part of ANT V and ANT VI, fore and middle legs uniformly yellow, hind legs yellow with slightly darker apical half of femora, and brown very distal ends of tibiae, tarsi yellow. ABD with light brown SIPH sclerites and small spinal sclerotisation on ABD IV and V (
Figure 16a).
Morphometric characters: HW 0.54–0.55 × ANT. ANT 0.28–0.32 × BL. ANT III with 8–12, ANT IV shorter than ANT V with 4–5 secondary rhinaria. ANT V as long as or longer than ANT VI with 1–5 rounded secondary rhinaria (
Figure 17a). ANT VI with 10–12 basal setae and without long, fine setae on PT. ANT VI with PT 0.25–0.36 × BASE, 0–1 small secondary rhinarium. Other antennal ratios: VI:III 0.40–0.46, V:III 0.46–0.51, IV:III 0.37–0.43. LS III about 5.20 × BD III. URS 0.51–0.59 × ANT III, 1.13–1.35 × ANT VI, and 0.75–0.82 × HT II with 13 accessory setae. Fore wings with unbranched media. Scale-like elements distributed on the wing membrane in the distal and cubital part of the wing (
Figure 18a). Hind tibiae setae 0.13–0.21 mm long. HT I with 12 ventral setae. HT II 0.61–0.77 × ANT III and 1.06–1.33 ANT VI. Dorsal setae 0.11–0.14 mm long. Abdominal sclerotisation: ABD I with hardly visible spinal sclerites, ABD II-IV without marginal sclerites, only with more setose areas. ABD IV-V with small poorly sclerotised spinal area. Less sclerotised part of the patch on ABD IV and V cuticle wrinkled irregularly (
Figure 19a and
Figure 20a). ABD VIII with 27–34 setae.
Material examined. Paratypes. INDIA, West Bengal, Darjeeling, 26 May 1958, Eriobotrya dubia, two apt. viv. fem., one al. viv. fem., five nymphs, S. Das leg., BM 1984-340 (Basu 200) NHMUK; 06 Jan1958, two apt. viv. fem., one nymph, BM 1984-340 (Basu 181) NHMUK; other material: INDIA, West Bengal, Darjeeling 15 Nov. 1968, E. dubia, S.G. Rajasingh leg., BM 1984-340 NHMUK.
Diagnosis. This species is similar in the pigmentation of the legs of apterous viviparous females to N. piri and N. chakrabartii sp. nov., but may be easily distinguished from all other species in the genus Nippolachnus by the lack of long, fine setae on the PT. Alate viviparous females may be distinguished from other morphs by antennae with smaller secondary rhinaria and hardly developed sclerotisation on abdomen.
Host plants.
Eriobotrya dubia,
Photinia arguta. Mandal et al. [
30] also reported this species from
Quercus sp. It is most likely that the records on
Pyrus communis,
P. pashia should refer to
N. chakrabartii sp. nov.
Distribution. India: Himachal Pradesh, Meghalaya, West Bengal. Holman [
9] also mentioned a record from China, after Mandal et al. [
30], although in the latter paper there is no record about
N. bengalensis in China.
Colour: in life, unknown. Mounted specimens: body in general yellow. Antennae with light brown ANT IV, V, and VI. Fore and middle legs with light brown distal part of tibiae and tarsi. III FEMORA pale in the proximal part, then yellow and with distinct brown pigmentation at the distal ends (more on the dorsal side). III TIBIAE brown with distinct dark brown distal ends and brown tarsi (
Figure 13b and
Figure 15b).
Morphometric characters: HW 0.64–0.71 × ANT. ANT 0.26–0.29 × BL. ANT III with 0–1 secondary rhinarium, ANT IV shorter than ANT V with 0–1 secondary rhinarium. ANT V as long as or minimally shorter than ANT VI. ANT VI with 16–18 basal setae and 1–2 long, fine setae on PT (
Figure 14b). ANT VI PT 0.45–0.54 × BASE. Other antennal ratios: VI:III 0.55–0.62, V:III 0.57–0.60, IV:III 0.37–0.46. LS III 3.00–3.66 × BD III. URS 0.61–0.68 × ANT III, 1.08–1.10 × ANT VI, and 0.72–0.82 × HT II with 18 accessory setae. Hind tibiae setae 0.07–0.18 mm long. HT I with 10–11 ventral setae (except sense peg). HT II 0.75–0.84 × ANT III and 1.32–1.51 ANT VI. Dorsal setae, 0.11–0.16 mm long. ABD VIII with 40–44 setae.
Colour: in life, unknown. Mounted specimens: antennae uniformly brown with only slightly lighter basal part of ANT III. Fore and middle legs uniformly yellow to light brown with slightly darker distal parts of femora, tibiae, and tarsi. III FEMORA brown with paler basal part and dark brown distal half. III TIBIAE brown with dark brown distal 1/3 of their length (
Figure 15b).
Morphometric characters: HW 0.68–0.79 × ANT. ANT 0.29–0.43 × BL. ANT III with 7–8, ANT IV shorter than ANT V with two secondary rhinaria. ANT V as long as ANT VI with two secondary rhinaria (
Figure 17b). ANT VI with 19–21 basal setae and 2–3 long, fine setae on PT. ANT VI with PT 0.47–0.52 × BASE. Other antennal ratios: VI:III 0.59–0.68, V:III 0.59–0.68, IV:III 0.40. LS III 3.66 × BD III. URS 0.59–0.60 × ANT III, 0.88–1.00 × ANT VI and 0.71–0.80 × HT II with 20–22 accessory setae. Fore wings with unbranched media. Scale-like elements distributed on the wing membrane in the distal part of Cu1a area and in the distal half of the M area of the wing (
Figure 18b). Hind tibiae setae 0.08–0.18 mm long. HT I with 12 ventral setae. HT II 0.74–0.84 × ANT III and 1.23–1.25 ANT VI. Dorsal setae 0.11–0.14 mm long. Abdominal sclerotisation: ABD I with spino-pleural cross-band and small marginal sclerites, ABD II and III only with marginal sclerites, ABD IV-V only with small, broken spinal sclerites. ABD VII with small scattered marginal sclerites or scleroites at setal bases, ABD VIII with broken cross-band. Less sclerotised part of the patch on ABD IV and V with pale and hardly visible cuticle (
Figure 19b and
Figure 20b). ABD VIII with 28–33 setae.
Material examined. Holotype. INDIA, West Bengal, Kalimpong, 02 Oct. 1958, Pyrus sp., one apt. viv. fem. marked with a circle and “H” letter, S. Das leg., BM 1984-340 (3) NHMUK; Paratypes. One al. viv. fem., other data the same as in holotype, BM 1984-340 (3) NHMUK; one apt. viv. fem., one al. viv. fem., other data the same as in holotype, BM 1984-340 (1) NHMUK; one apt. viv. fem., other data the same as in holotype, BM 1984-340 (4) NHMUK; 07 Mar. 1958, one apt. viv. fem., two nymphs, other data as in holotype, BM 1984-340 (2) NHMUK; one ap. viv. fem., one nymph, BM 1984-340 (5) NHMUK; two apt. viv. fem., BM 1984-340 (6) DZUS;
Diagnosis. In the pigmentation of hind tibiae, the new species is similar to N. bengalensis and N. piri. Apterous viviparous females of the new species can be easily distinguished from both species first of all by the pigmentation of the legs: (1) fore and middle tibiae with darker apical parts (fore and middle tibiae uniformly pigmented in N. bengalensis and N. piri), (2) hind femora with brown distal part (hind femora uniformly pigmented). From N. bengalensis, apterae may be additionally distinguished by (1) 1–2 long setae on the PT (PT of N. bengalensis without long setae), (2) URS with 18 accessory setae (8–10 accessory setae in N. bengalensis), (3) PT base 0.45–0.54 (0.27–0.32 in N. bengalensis), (4) URS/ANT III 0.61–0.68 (0.37–0.40 in N. bengalensis). Additionally, apterae of N. chakrabartii sp. nov. differ from those of N. piri in having the following: (1) a smaller number of PT long setae—1–2 (3–4 in N. piri); (2) ANT V/ANT III 0.57–0.60 (0.47–0.56 in N. piri); (3) LS/BD III 3.00–3.66 (4.07–7.50 in N. piri); (4) HT I basal length/H I dorsal length 4.00–4.50 (1.60–2.00 in N. piri). Alate viviparous females of the new species together with the alatae of N. bengalensis differ from those of the other species in having the following: unbranched media of fore wings (once-branched in other Nippolachnus and twice-branched in Indolachnus); secondary rhinaria as wide as or narrower than half the segment width (secondary rhinaria much wider than half the width of the segment); and poorly developed spinal abdominal sclerotisation. The alate viviparous females of the new species differ from those of N. bengalensis in having the following: (1) much darker and pigmentation broader pigmentation of III FEMORAE (only slightly darker very distal ends in N. bengalensis), (2) dark brown 1/3 distal end of III TIBIAE (slightly darker very distal ends in N. bengalensis), (3) spinal abdominal sclerotisation on ABD III-V (spinal abdominal sclerotisation only on ABD III-IV in N. bengalensis), (4) HW/ANT 0.68–0.69 (0.54–0.55 in N. bengalensis), (5) ANT VI/ANT III 0.59–0.69 (0.40–0.46 in N. bengalensis).
Etymology. We have the pleasure of naming the new species to honour the outstanding Indian aphidologist Professor Samiran Chakrabarti, our colleague and friend, and for many years the Head of the Department of Zoology, University of Kalyani, India.
Host plants. The species has been collected from an unidentified species of Pyrus tree.
Distribution. Representatives of the new species are so far known from Kalimpong in West Bengal (India).
Colour: in life, unknown. Mounted specimens: body pale with pale appendages. Only ANT VI PT, distal end of hind tibiae and hind tarsi light brown (
Figure 13c and
Figure 15c).
Morphometric characters: HW 0.44–0.48 × ANT. ANT 0.34–0.40 × BL. ANT IV always shorter than ANT V. ANT V always shorter than ANT VI. ANT VI with 17–21 basal setae and 1–2 long, fine setae on PT (
Figure 14c). ANT VI PT 0.44–0.69 × BASE. Other antennal ratios: VI:III 0.66–0.81, V:III 0.53–0.54, IV:III 0.33–0.36. LS III 5.50–5.75 × BD III. URS 0.39–0.46 × ANT III, 0.51–0.66 × ANT VI and 0.59–0.70 × HT II with 14–15 accessory setae. Hind tibiae setae 0.05–0.15 mm long. HT I with 8–10 ventral setae (except sense peg). HT II 0.65–0.66 × ANT III and 0.80–1.00 ANT VI. Dorsal setae, 0.09–0.11 mm long. ABD VIII with 36–40 setae.
Material examined: Holotype. MALAYSIA, Cameron Highlands, 6 Oct. 1944, Pyrus granulosa, one apt. viv. fem. Marked with “H” and circle, R. Takahashi leg., 774/55, BM.1955.799 NHMUK; Paratypes. Two apt. vivi. fem., other data as in holotype, 774/55, BM.1955.799 NHMUK; three apt. vivi. fem. other data as in holotype, 773/55, BM.1955.799 NHMUK.
Diagnosis. Apterous viviparous females of Nippolachnus malayaensis sp. nov. are most similar to N. sinensis sp. nov. in having a very pale body and appendages in which both species differ from N. bengalensis and N. chakrabartii sp. nov. Both species differ, moreover, in the chaetotaxy of ANT BASE (16–17), URS (15–16), and ABD VIII (36–42) from N. micromeli, N. piri, and N. xitianmushanus, in which the values of those characters are 20–27, 18–25, and 45–52, respectively. Nippolachnus malayaensis sp. nov. can be easily distinguished from N. sinensis sp. nov. by the following differences: (1) HW/ANT 0.44–0.48 (0.62–0.63 in N. sinensis sp. nov.), (2) ANT VI/ANT III 0.66–0.81 (0.57–0.63 in N. sinensis sp. nov.), (3) URS/ANT III 0.39–0.46 (0.51–0.57), (4) URS/ANT VI 0.59–0.66 (0.81–1.00 in N. sinensis sp. nov.), (5) HT II/ANT III 0.65–0.66 (0.72–0.75 in N. sinensis), (6) HT II/ANT VI 0.80–1.00 (1.13–1.31 in N. sinensis sp. nov.), (7) HT I basal length/HT I dorsal length 1.20–1.50 (2.05–2.33 in N. sinensis sp. nov.).
Etymology. The name of the new species is derived from “Malaya”, the former name of Malaysia.
Host plants. The species has been collected from Sorbus granulosa (=Pyrus granulosa).
Distribution. So far, the species is known from the Cameron Highlands at 1524 m. above sea level (5000 ft) in Malaysia.
Nippolachnus micromeli Shinji, 1924
Shinji, 1924: 343
=N. micromelli Shinji, 1941 [
32], p. 227; Shiraki, 1952 [
33], p. 103
=Neonippolachnus betulae Shinji, 1924 syn. nov.
(
Figure 2,
Figure 4,
Figure 13,
Figure 14,
Figure 15,
Figure 16,
Figure 17,
Figure 18,
Figure 19,
Figure 20,
Figure 21,
Figure 22 and
Figure 23;
Table 1,
Table 2 and
Table 3)
Figure 21.
Previously unknown morphs of Nippolachnus micromeli and N. piri: (a) fundatrix of N. micromeli, (b) fundatrix of N. piri, (c) oviparous female N. micromeli, (d) oviparous female N. piri, (e) alate male N. micromeli, (f) alate male N. piri.
Figure 21.
Previously unknown morphs of Nippolachnus micromeli and N. piri: (a) fundatrix of N. micromeli, (b) fundatrix of N. piri, (c) oviparous female N. micromeli, (d) oviparous female N. piri, (e) alate male N. micromeli, (f) alate male N. piri.
Table 3.
Measurements (in mm) of fundatrices and sexual morphs of N. micromeli and N. piri.
Table 3.
Measurements (in mm) of fundatrices and sexual morphs of N. micromeli and N. piri.
| Character | N. micromeli | N. piri |
|---|
| Fundatrix | Oviparous Female | Alate male | Fundatrix | Oviparous Female | Alate Male |
|---|
| BL | 3.30–4.10 | 2.87–3.40 | 2.12–2.50 | 4.00–4.25 | 4.12–4.25 | 2.32–2.90 |
| HW | 0.52–0.56 | 0.48–50 | 0.50–0.55 | 0.57–0.65 | 0.55–0.58 | 0.56–0.52 |
| ANT | 0.86–0.92 | 0.97–1.01 | 1.13–1.31 | 1.05–1.12 | 1.11–1.16 | 1.22–1.41 |
| ANT III | 0.29–0.33 | 0.35–0.38 | 0.44–0.49 | 0.38–0.42 | 0.38–0.44 | 0.47–0.55 |
| ANT IV | 0.10–0.11 | 0.10–0.11 | 0.13–0.18 | 0.12–0.13 | 0.13–0.15 | 0.16–0.19 |
| ANT V | 0.13–0.14 | 0.18–0.19 | 0.19–0.26 | 0.16–0.17 | 0.18–0.20 | 0.19–0.23 |
| ANT VI | 0.17–0.20 | 0.18–0.19 | 0.21–0.23 | 0.19–0.21 | 0.22–0.23 | 0.25–0.29 |
| BASE | 0.11–0.13 | 0.12 | 0.15–0.16 | 0.13–0.15 | 0.15–0.16 | 0.17–0.20 |
| PT | 0.60–0.70 | 0.60–0.70 | 0.06–0.07 | 0.06–0.07 | 0.06–0.07 | 0.08–0.09 |
| URS | 0.20–0.21 | 0.18–0.19 | 0.15–0.17 | 0.21–0.22 | 0.20–0.21 | 0.18–0.20 |
| III FEMORA | 1.12–1.22 | 1.25–1.30 | 0.90–1.12 | 1.35–1.50 | 1.65–1.67 | 0.95–1.12 |
| III TIBIAE | 2.10–2.30 | 2.42–2.45 | 1.75–2.00 | 2.40–2.75 | 2.90–3.00 | 1.85–2.17 |
| HT II | 0.20–0.20 | 0.23–0.24 | 0.17–0.20 | 0.22–0.23 | 0.24–0.25 | 0.17–0.19 |
Figure 22.
Morphological characters of males: (a) antennal sensilla—secondary rhinaria on ANT III of N. micromeli, (b) antennal sensilla—primary and secondary rhinaria on ANT VI of N. micromeli, (c) genitalia of N. micromeli, (d) antennal sensilla—secondary rhinaria on ANT III of N. piri, (e) antennal sensilla—primary and secondary rhinaria on ANT VI of N. piri, (f) genitalia of N. piri, (g) abdomen of N. micromeli, (h) abdomen of N. piri; white dotted line indicates the major rhinarium, red dotted line indicates accessory rhinaria, black line indicates secondary rhinaria on PT.
Figure 22.
Morphological characters of males: (a) antennal sensilla—secondary rhinaria on ANT III of N. micromeli, (b) antennal sensilla—primary and secondary rhinaria on ANT VI of N. micromeli, (c) genitalia of N. micromeli, (d) antennal sensilla—secondary rhinaria on ANT III of N. piri, (e) antennal sensilla—primary and secondary rhinaria on ANT VI of N. piri, (f) genitalia of N. piri, (g) abdomen of N. micromeli, (h) abdomen of N. piri; white dotted line indicates the major rhinarium, red dotted line indicates accessory rhinaria, black line indicates secondary rhinaria on PT.
Figure 23.
Morphological characters of oviparous females: (a) scent plaques of N. micromeli, (b) scent plaques of N. piri, (c) perianal structures of oviparous female N. micromeli, VIII—abdominal segment VIII, c—cauda, ap—anal plate, gp—genital plate.
Figure 23.
Morphological characters of oviparous females: (a) scent plaques of N. micromeli, (b) scent plaques of N. piri, (c) perianal structures of oviparous female N. micromeli, VIII—abdominal segment VIII, c—cauda, ap—anal plate, gp—genital plate.
Colour: in life, body in general green. Head light green. Thorax and abdomen green with darker marginal areas and spinal area on abdomen. Antennae and legs pale to light green (
Figure 2a,b). Mounted specimens: body pale, ANT pale yellow, fore and middle legs pale yellow, hind femora yellow with brown patches on the distal ends, hind tibiae yellow with dark distal ends and dark tarsi (
Figure 21a).
Morphometric characters: HW 0.56–0.65 × ANT. ANT 0.20–0.28 × BL. ANT IV always shorter than ANT V. ANT V always shorter than ANT VI. ANT VI with 21–24 basal setae and 2–5 long, fine setae on PT. ANT VI PT 0.48–0.59 × BASE. Other antennal ratios: VI:III 0.58–0.65, V:III 0.42–0.46, IV:III 0.31–0.36. LS III 6.00–6.50 × BD III. URS 0.63–0.68 × ANT III, 1.05–1.14 × ANT VI and 0.95–1.00 × HT II with 25–27 accessory setae. Hind tibiae setae 0.08–0.14 mm long. HT I with 10 ventral setae (except sense peg). HT II 0.66–0.72 × ANT III and 1.11–1.14 ANT VI. Dorsal setae, 0.09–0.15 mm long. ABD VIII with 35–46 setae.
Remarks: The fundatrix can be easily distinguished from the apterous viviparous females by the following differences: (1) larger body 3.30–4.10 mm long (2.25–2.97 in apt. viv. fem.), (2) ANT V/ANT III 0.42–0.48 (0.50–0.58 in apt. viv. fem.), (3) ANT IV/ANT V 0.75–0.78 (0.50–0.66 in apt. viv. fem.), (4) HT II/ANT VI 1.10–1.14 (1.22–1.57 in apt. viv. fem).
Colour: in life, body whitish-green to pale green with pale green appendages and some darker spots in the spinal area of the abdomen, which is the same as in larvae and alatoid nymphs (
Figure 2c). Mounted specimens: body pale, almost colourless, only the legs are very pale yellowish with the same pale antennae. III TIBIAE pale yellowish with distal part light brown. HT I pale, HT II brown with paler proximal base (
Figure 13d and
Figure 15d).
Morphometric characters: HW 0.55–0.64 × ANT. ANT 0.26–0.37 × BL. ANT III without secondary rhinaria, ANT IV always shorter than ANT V. ANT V shorter or slightly longer than ANT VI. ANT VI with 22–24 basal setae and 4–6 long, fine setae on PT (
Figure 14d). ANT VI PT 0.45–0.60 × BASE. Other antennal ratios: VI:III 0.51–0.66, V:III 0.50–0.58, IV:III 0.25–0.34. LS III 4.33–6.00 × BD III. URS 0.48–0.60 × ANT III, 0.80–1.07 × ANT VI and 0.63–0.78 × HT II with 20–25 accessory setae. Hind tibiae setae 0.07–0.16 mm long. HT I with 10–12 ventral setae (except sense peg). HT II 0.70–0.84 × ANT III and 1.22–1.57 ANT VI. Dorsal setae, 0.11–0.15 mm long. ABD VIII with 40–45 setae.
Colour: in life, head and thorax dark brown to black with thin wax layer. Antennae dark, legs with brown femora and pale brown tibiae with darker distal ends. Abdomen with dark sclerotisation and white wax powder (
Figure 2d). Mounted specimens: body pale, almost colourless, only the legs are very pale yellowish with the same pale antennae. III TIBIAE pale yellowish with distal part light brown. HT I pale, HT II brown with paler proximal base (
Figure 16c).
Morphometric characters: HW 0.56–0.61 × ANT. ANT 0.31–0.34 × BL. ANT III with 5–7 secondary rhinaria, ANT IV shorter than ANT V with 2 secondary rhinaria. ANT V as long as or slightly shorter than ANT VI with 1–3 secondary rhinaria (
Figure 17c). ANT VI with 23–26 basal setae and 6–8 long, fine setae on PT. ANT VI with PT 0.50–0.51 × BASE. Other antennal ratios: VI:III 0.51–0.64, V:III 0.50–0.62, IV:III 0.34–0.40. LS III 4.75–6.00 × BD III. URS 0.43–0.46 × ANT III, 0.73–0.85 × ANT VI, and 0.65–0.73 × HT II with 28–30 accessory setae. Media of fore wings once-forked in the middle of the length. Scale-like elements distributed on the wing membrane at distal end of Cu1a, M, and M2 (M1 area uniformly covered) (
Figure 18c). Hind tibiae setae 0.08–0.17 mm long. HT I with 12 ventral setae. HT II 0.60–0.71 × ANT III and 1.07–1.23 ANT VI. Dorsal setae 0.11–0.14 mm long. Abdominal sclerotisation: ABD II and III marginal sclerite. On ABD III-V, large spino-pleural patch which is more sclerotised and darker in the spinal area, ABD VI with spinal sclerite and pleural scattered sclerites originating from scleroites at setal bases. ABD VII with spino-pleural cross-band with irregular edges. ABD VIII with spino-pleural cross-band with smooth edges. Less sclerotised part of the patch on ABD III–V cuticle only slightly wrinkled without regular polygons (
Figure 19c and
Figure 20c). ABD VIII with 38–42 setae.
Colour: in life, body light green with darker patches in the marginal areas of thorax, ABD II, III, and spinal area on ABD IV–V. Legs pale green with pale brown to brown distal ends of tibiae and brown tarsi (
Figure 2f,i). Mounted specimens: body pale with yellow appendages (ANT and legs), only hind legs with slightly darker apical ends of tibiae and light brown tarsi (
Figure 21c).
Morphometric characters: HW 0.47–0.51 × ANT. ANT 0.29–0.34 × BL. ANT IV always shorter than ANT V. ANT V shorter or longer than ANT VI. ANT VI with 19–21 basal setae and 6–7 long, fine setae on PT. ANT VI PT 0.54–0.58 × BASE. Other antennal ratios: VI:III 0.48–0.54, V:III 0.48–0.51, IV:III 0.27–0.28. LS III 5.50–6.25 × BD III. URS 0.47–0.51 × ANT III, 0.94–1.00 × ANT VI and 0.75–0.82 × HT II with 27–30 accessory setae. Hind tibiae setae 0.05–0.17 mm long. Hind tibiae poorly swollen with several dozen small, rounded scent plaques (
Figure 24a). HT I with 10–12 ventral setae (except sense peg). HT II 0.60–0.68 × ANT III and 1.21–1.26 ANT VI. Dorsal setae, 0.11–0.15 mm long. ABD VIII with 50–60 setae (
Figure 24c).
Colour: in life, head brown, slightly covered in wax, eyes black, antennae brown. Thorax brown, in some places slightly wax powdered. Legs pale with brown distal ends of hind femora, very distal ends of tibiae and tarsi. Wings hyaline with brown pterostigma. Abdomen greenish with brown to dark sclerotisation (
Figure 4g,h). Mounted specimens: head and thorax sclerotised, light brown, ANT yellow with slightly darker apical part of ANT VI, legs yellow with light brown distal ends of tibiae and light brown tarsi, wings hyaline with yellow pterostigma, ABD sclerotisation light brown (
Figure 21e).
Morphometric characters: HW 0.41–0.44 × ANT. ANT 0.46–0.53 × BL. ANT III with 53–64 in general small and rounded secondary rhinaria of which 2–5 on the basal part are much larger than the others (
Figure 22a). ANT IV shorter than ANT V with 20–25 secondary rhinaria. ANT V always shorter or longer than ANT VI with 17–22 secondary rhinaria. ANT VI with 12–16 secondary rhinaria, of which few (2–3) are located not only on the BASE but also on PT (
Figure 22b), 24–26 basal setae, and 5–8 long, fine setae on PT. ANT VI PT 0.38–0.43 × BASE. Other antennal ratios: VI:III 0.46–0.52, V:III 0.43–0.53, IV:III 0.29–0.36. LS III 3.33–4.00 × BD III. URS 0.34–0.38 × ANT III, 0.65–0.79 × ANT VI and 0.87–1.00 × HT II with 32–34 accessory setae. Hind tibiae setae 0.06–0.13 mm long. HT I with 13–15 ventral setae (except sense peg). HT II 0.38–0.40 × ANT III and 0.73–0.86 ANT VI. Dorsal setae, 0.08–0.11 mm long. ABD VIII with 24–26 setae. Abdomen with spinal sclerites on ABD I–VI, spino-pleural cross-bar on ABD VII and cross-band on ABD VIII. Marginal sclerites on ABD II–IV (
Figure 22g). Genitalia sclerotised with robust and setose parameres (
Figure 22c).
Material examined: Neotype. JAPAN, Amakubo, Tsukuba, 15 Oct. 2016, Rhaphiolepis indica var. umbellata, one al. viv. fem., M. Miyazaki leg., Jap/16/10/2 DZUS (designated by Kanturski et al. 2018); other material: one al. viv. fem., other data as in Neotype, Jap/16/10/03 DZUS; one al. viv. fem., other data as in Neotype, Jap/16/10/04 DZUS; one ♀, other data as in Neotype, Jap/16/10/01 DZUS; Sapporo, 19 Sep. 1962, Ulmus japonica (=Ulmus davidiana var. japonica), four al. viv. fem., two nymphs, M. Sorin leg., 14,494 MNHN; 21 Sep. 1962, three al. viv. fem., three nymphs, 14,497 MNHN; five al. viv. fem., one nymph, 210,102,830 NIAES, 22 Sep. 1966, Sorbus alnifolia, one al. viv. fem., M. Miyazaki leg., 1996 (1) NIAES, one al. viv. fem., 1996 (2) NIAES; 12 apt. viv. fem., M. Sorin leg., 14,502 MNHN; Senami, Murakami, 17 Jan. 1992, Rhaphiolepis indica var. umbellata, four ♀, K. Baba leg., 14,513 MNHN; two ♀, 210102831, NIAES; Nikko, 28 Sep. 1975, S. alnifolia, five al. viv. fem., M. Sorin leg., 14,506 MNHN; 24 Aug. 1980, one apt. viv. fem., two nymphs, R. Blackman leg., 2141 NHMUK; KOREA, Mt. Oseo, 15 Oct. 2011, Sorbus alnifolia, one al. viv. fem., Y. Lee leg., Kor/11/10/01 DZUS; one al. viv. fem., Kor/11/10/02 DZUS; Jeonnan Univ. Arborettum, 7 May 2016, Rhaphiolepis indica var. umbellata, one al. viv. fem., Y. Lee leg., Kor/16/05/01 DZUS; one al. viv. fem., Kor/16/05/02 DZUS; two al. viv. fem., Kor/16/05/03 DZUS; R. indica, two al. viv. fem., Kor/16/05/4; Geumoch Island, 5 Jun. 2016, Sorbus sp., two apt. viv. fem., J. Choi leg., Kor/16/06/1 DZUS; two apt. viv. fem., Kor/16/06/2; Cheollipo 1-gil, Sawon-myeon, Taean-gun, Chungcheongnam-do, 7 Nov. 2020, Rhaphiolepis indica, var. umbellata, four ♀, three ♂, M. Lee leg., 201107-LMH-1(1–7) SNU; 14 May 2023, three Fx, 230514-LMH-1 (1–3), DZUS.
Diagnosis. In pigmentation of mounted specimens, very pale apterous viviparous females of N. micromeli are similar to N. malayaensis sp. nov. and N. sinensis sp. nov. Both new species are characterised by almost pigmentless appendages (especially legs), in which they differ from N. micromeli (and other species in which the legs are yellow or with brown distal ends). Additionally, N. micromeli differs from the new species in the chaetotaxy of ANT VI BASE (20–22 setae), URS (20–25), and ABD VIII (45–50), which in N. malayaensis sp. nov. and N. sinensis sp. nov. have 16–21, 14–16, and 36–42 setae, respectively. The large number of setae of remaining body parts makes N. micromeli similar to N. piri and N. xitianmushanus together with darker pigmented distal ends of tibiae. N. micromeli differs from both species in the following: (1) unpigmented ANT VI (pigmented in N. piri and N. xitianmushanus); (2) 20–22 basal setae (25–27 in N. piri and N. xitianmushanus); (3) 4–6 long setae on PT (3–4 in N. piri and N. xitianmushanus). Apterous viviparous females of N. micromeli differ from those of N. piri additionally by having a higher ratio of HT I basal to HT I dorsal length, 2.00–3.50 (1.60–2.00 in N. piri), and from those of N. xitianmushanus by ANT IV/ANT V 0.50–0.66 (0.73–0.91 in the latter). Alate viviparous females can be easily distinguished by abdominal sclerotisation with spinal and spino-pleural cross-bars on ABD VI and VII, which are absent in the known alatae of the other species.
Host plants. Rhaphiolepis indica, R. umbellata, Sorbus alnifolia, Sorbus sp. Ulmus davidiana var. japonica.
Biology. In the Korean Peninsula, the first representatives of this species—fundatrices and first instar fundatrigeniae—were observed in mid-May on
R. indica var.
umbellata (
Figure 4a) and with some time, each observed colony exhibited the presence of the fundatrix and alatoid nymphs without apterous female larvae. From the beginning of the colonies’ activity, the aphids were attended by
Crematogaster matsumurai ants (
Figure 4e). In the autumn, the colonies consisted of alate viviparous females, oviparous females, and male nymphs. In late October, alate viviparous females gave birth to the sexual generation, and after mating, the oviparous females laid eggs in a circular and disordered pattern around the main vein on the undersides of the host plant where they overwinter.
Distribution. This species is known from Japan and the Korean Peninsula.
Remarks. For a long time,
Nippolachnus micromeli has been treated as a synonym of
N. piri. Kanturski et al. [
13] performed detailed morphological analyses supported by molecular studies and restored
N. micromeli as a separate species. All previous records of
N. piri from the above-mentioned host plants should be referred to
N. micromeli.
Fundatrix—description
Colour: in life, head light brown. ANT pale yellowish. Fore legs yellow, middle legs with green-yellowish femora, and pale yellowish tibiae. Hind legs with light green proximal parts, and yellow distal parts of femora and yellow tibiae with dark distal ends and dark tarsi. Dorsal side of body yellow with green marginal areas of thorax and ABD I–VI. On ABD VII and VII, green pleuro-marginal cross-bands. Spinal areas of thorax and abdomen with brown patches, especially on ABD I and ABD IV–VI (
Figure 3a,b). Mounted specimens: head slightly sclerotised, yellow, ANT pale yellow with light brown distal part of ANT VI. Fore and middle legs yellow. Hind legs yellow with dark distal ends of tibiae and dark tarsi (
Figure 21b).
Morphometric characters: HW 0.54–0.59 × ANT. ANT 0.25–0.26 × BL. ANT IV always shorter than ANT V. ANT V always shorter than ANT VI. ANT VI with 22–25 basal setae and 0–4 long, fine setae on PT. ANT VI PT 0.40–0.50 × BASE. Other antennal ratios: VI:III 0.48–0.55, V:III 0.40–0.42, IV:III 0.28–0.32. LS III 6.50–8.00 × BD III. URS 0.52–0.57 × ANT III, 1.04–1.07 × ANT VI, and 0.95–1.00 × HT II with 18–22 accessory setae. Hind tibiae setae 0.11–0.17 mm long. HT I with 9–10 ventral setae (except sense peg). HT II 0.54–0.57 × ANT III and 1.04–1.12 ANT VI. Dorsal setae, 0.10–0.16 mm long. ABD VIII with 43–55 setae.
Remarks: The fundatrix can be easily distinguished from the apterous viviparous females by the following differences: (1) larger body 4.00–4.25 mm long (2.27–3.55 in apt. viv. fem.); (2) longer ANT III 0.38–0.42 (0.30–0.34 in apt. viv. fem.); (3) ANT V/ANT III 0.40–0.42 (0.47–0.56 in apt. viv. fem.); (4) URS/HT II 0.95–1.00 (0.70–0.86 in apt. viv. fem.).
Colour: in life, head yellowish-light green, ANT pale to light green with light brown PT. Pronotum green, rest of thorax light green with green longitudinal stripes on the margin, fore and middle legs pale green with dusky, hind legs with greenish femora, light brown tibiae and dark tarsi, abdomen light green with green longitudinal marginal stripes, spinal patches on ABD I, IV, and V, and green cross-bars on ABD VII (
Figure 2c,d). Mounted specimens: body in general pale. Head pale or yellow, ANT pale with light brown to brown distal part of ANT V and distal part of ANT VI, fore and middle legs pale to yellow, hind femora yellow with pale proximal part, hind tibiae pale yellow to yellow with dark distal end and both segments of dark tarsi (
Figure 13e and
Figure 15e).
Morphometric characters: HW 0.54–0.62 × ANT. ANT 0.26–0.42 × BL. ANT III 1.03–1.23 × ANT IV + ANT V with 0–1 secondary rhinarium, ANT IV with 0–1 secondary rhinarium. ANT V shorter or as long as ANT VI. ANT VI with 25–27 basal and 3–4 long, fine setae on PT (
Figure 14c). ANT VI PT 0.41–0.54 × BASE. Other antennal ratios: VI:III 0.52–0.60, V:III 0.47–0.56, IV:III 0.30–0.41. LS III 4.07–7.50 × BD III. URS 0.56–0.62 × ANT III, 0.97–1.14 × ANT VI and 0.70–0.86 × HT II with 18–21 accessory setae. Hind tibiae setae 0.06–0.18 mm long. HT I with 8–9 ventral setae (except sense peg). HT II 0.67–0.80 × ANT III and 1.14–1.47 ANT VI. Dorsal abdominal setae, 0.11–0.16 mm long. ABD VIII with 50–55 setae.
Colour: in life, head and thorax brown. Antennae dark, femora and tibiae of legs brown with dark distal ends. Abdomen dark brown with white wax patches (
Figure 3g). Very freshly moulted specimens with brown head (with darker antennae), yellow thorax, and brown-greenish legs with darker distal ends. Abdomen light green with darker patches in the central and marginal part, without wax (
Figure 3e). The freshly moulted specimens after some time become darker and start to be slightly visible on the abdomen (
Figure 3f). Mounted specimens: body with brown head, yellowish-brown ANT with brown apex of ANT IV, ANT V, and VI (
Figure 1c,d,f). Fore and middle legs uniformly pale yellow to yellow, hind legs with yellow femora, and yellow to light brown tibia with dark distal parts and whole tarsi (
Figure 16d).
Morphometric characters: HW 0.59–0.65 × ANT. ANT 0.28–0.33 × BL. ANT III with 8–11 secondary rhinaria, ANT IV shorter than ANT V with 2–3 secondary rhinaria. ANT V shorter or as long as ANT VI with 1–2 secondary rhinaria (
Figure 17d). ANT VI with XX basal setae and 3–4 long, fine setae on PT. ANT VI with PT 0.42–0.50 × BASE. Other antennal ratios: VI:III 0.55–0.60, V:III 0.47–0.55, IV:III 0.35–0.38. LS III 3.20–5.20 × BD III. URS 0.48–0.57 × ANT III, 0.80–1.00 × ANT VI, and 0.70–0.90 × HT II with 18–20 accessory setae. Scale-like elements distributed in the distal part of Cu1a, M1, and M2 areas of the membrane. Media one-branched with the fork in the distal half of the pterostigma length under the Rs rising (
Figure 18d). Hind tibiae setae 0.13–0.21 mm long. HT I with 12 ventral setae. HT II 0.61–0.77 × ANT III and 1.06–1.33 ANT VI. Dorsal setae 0.11–0.14 mm long. Abdominal sclerotisation: ABD I with spino-pleural cross-band and small marginal sclerites, ABD II and III with pleural and marginal sclerites and very small sclerites at setal bases between them, ABD IV with marginal sclerites more or less fused with SIPH sclerites and large sclerotic spino-pleural cross-band, ABD V proximal part with spino-pleural cross-band fused with the previous one, ABD VI and VII without sclerites, ABD VIII with pleuro-marginal cross-band. Less sclerotised part of the patch on ABD IV and V cuticle wrinkled or rugose in the form of polygonal reticulation (
Figure 19d and
Figure 20d). ABD VIII with 30–33 setae.
Colour: in life, head pale yellow, antennae pale yellow with brown distal part of ANT VI. Prothorax green with yellowish spinal part. The rest of the thorax and abdomen yellowish with green marginal stripes and green spinal patches on metathorax, ABD I, IV, V, VI, and VII. Legs yellowish with brown distal ends of tibiae and tarsi (
Figure 3h,i,k). Mounted specimens: head and thorax pale yellow, abdomen pale, ANT yellow with slightly darker apical part of ANT VI, legs yellow with dark apical ends of hind tibiae and dark hind tarsi (
Figure 21d).
Morphometric characters: HW 0.54–0.59 × ANT. ANT 0.25–0.26 × BL. ANT IV always shorter than ANT V. ANT V always shorter than ANT VI. ANT VI with 27–30 basal setae and 3–5 long, fine setae on PT. ANT VI PT 0.37–0.46 × BASE. Other antennal ratios: VI:III 0.50–0.60, V:III 0.45–0.52, IV:III 0.29–0.39. LS III 6.80–10.00 × BD III. URS 0.45–0.52 × ANT III, 0.86–0.95 × ANT VI, and 0.83–0.95 × HT II with 15–18 accessory setae. Hind tibiae setae 0.08–0.20 mm long. Hind tibiae with several dozen small, rounded scent plaques (
Figure 23b). HT I with 8–10 ventral setae (except sense peg). HT II 0.54–0.63 × ANT III and 1.04–1.13 ANT VI. Dorsal setae, 0.10–0.16 mm long. ABD VIII with 38–44 setae.
Colour: in life, head blackish, slightly covered in wax powder, antennae brown with darker ANT IV-VI. Thorax brown, femora of legs brown with darker distal halves, tibiae pale brown with dark distal ends and dark tarsi. Wing hyaline with brown pterostigma. Abdomen whitish with greenish-brown sclerotised cross-bands (
Figure 3j,k). Mounted specimens: head and thorax sclerotised, brown. ANT light brown with slightly darker distal part of ANT V and ANT VI. Legs yellow with brown distal parts of tibiae and brown tarsi of fore and middle legs. Distal ends of hind tibiae and tarsi dark brown. Wings hyaline with brown pterostigma. Abdominal sclerotisation brown (
Figure 21f).
Morphometric characters: HW 0.43–0.45 × ANT. ANT 0.47–0.54 × BL. ANT III with 95–115 small and rounded secondary rhinaria of which 3–4 on the basal part are much larger (
Figure 22b), ANT IV always shorter than ANT V with 25–43 secondary rhinaria. ANT V always shorter than ANT VI with 26–35 secondary rhinaria. ANT VI with 21–28 secondary rhinaria also on PT (
Figure 22e), 20–24 basal setae, and 3–6 long, fine setae on PT. ANT VI PT 0.40–0.47 × BASE. Other antennal ratios: VI:III 0.50–0.55, V:III 0.40–0.44, IV:III 0.32–0.34. LS III 3.33–4.00 × BD III. URS 0.34–0.38 × ANT III, 0.62–0.72 × ANT VI, and 0.92–1.05 × HT II with 20–25 accessory setae. Hind tibiae setae 0.06–0.15 mm long. HT I with 10 ventral setae (except sense peg). HT II 0.34–0.37 × ANT III and 0.67–0.70 ANT VI. Dorsal setae, 0.10–0.13 mm long. ABD VIII with 17–22 setae. Abdomen with spino-pleural cross-bars on ABD I–VI and cross-bands on ABD VII and VIII. Marginal sclerites on ABD I–IV (
Figure 22h). Genitalia strongly sclerotised with both robust and sclerotised parameres and the basal part of phallus (
Figure 22f).
Material examined: Neotype. JAPAN, Shizuoka-ken, 14 Apr. 2015, Eriobotrya japonica, one al. viv. fem., M. Sano leg., Jap/15/04/1 DZUS (designated by Kanturski et al. 2018); Fukoka Univ., 01 May 1964, two al. viv. fem., R. v.d. Bosh leg., BM 1984-340 NHMUK; Tokio, 23 Jul. 1956, Pyrus sp., five apt. viv. fem., one al. viv. fem., R. Takahashi leg., BM 1984-340 NHMUK; Mt. Koya, 23 Sep. 1960, Pyrus sp., two al. viv. fem., two nymphs, M. Sorin leg., 14,487 MNHN; Kuroyama, Osaka, 21 Jun. 1961, Pyrus sp., one apt. viv. fem., one al. viv. fem., M. Sorin leg., 210,102,829 NIAES; Yatabe, 01 Oct.1980, Pyrus communis, two apt. viv. fem., M. Miyazaki leg., 3754 (1) NIAES; one al. viv. fem., 3754 (2) NIAES; one al. viv. fem., 3754 (3) NIAES; two al. viv. fem., 3754 (4) NIAES; Fukoka Univ., 21 Apr. 1964, Eriobotrya sp., two al. viv. fem., R. v.d. Bosh leg., BM 1984-340 (1) NHMUK; two al. viv. fem., one nymph, BM 1984-340 (2) NHMUK; two al. viv. fem., one nymph, BM 1984-340 (3) NHMUK; one Fx, one al. viv. fem., BM 1984-340 (4) NHMUK; 15.04.1964, Fukoka Univ., 21 Apr. 1964, two Fx, BM 340-1984 (1) NHMUK; two Fx, BM 340-1984 (2) NHMUK; two Fx, one nymph, BM 340-1984 (3) NHMUK; two fundatrices, BM 340-1984 (4) NHMUK; two Fx, BM 340-1984 (5) NHMUK; KOREA, Chusan experimental forest, 17 Jun. 2016, Pyrus pyrifolia, one apt. viv. fem., Y. Lee leg., Kor/16/06/1 DZUS; one apt. vivi. fem., one al. viv. fem., Kor/16/06/2 DZUS; one apt. viv. fem., one al. viv. fem., Kor/16/06/3 DZUS; one apt. viv. fem., one al. viv. fem., Kor/16/06/4 DZUS; Odong-do, 14 Jul. 2014, P. pyrifolia, two apt. viv. fem., Y. Lee leg., Kor/14/07/1; two apt. viv. fem., Kor/14/07/2 DZUS; Sangbal-ri, Yongsan-myeon, Jangheung-gun, Jeollanam-do, 12 Nov. 2020, Eriobotrya indica, four ♀, three ♂, M. Lee leg., 221112-LMH-1 (1–7) SNU; Expo-daero, Sora-myeon, Yeosu-si, Jeollanam-do, four Fx, 230513-LMH-1(1–4) SNU.
Diagnosis. Apterous viviparous females of N. piri are similar to N. bengalensis, N. chakrabartii sp. nov. and N. xitianmushanus in the pigmentation of the legs, in which they differ from the other apterae, which have colourless or only slightly pigmented very distal ends of antennae, tibiae, and tarsi. Apterae of N. piri can be easily distinguished from N. bengalensis by the much darker distal area of hind tibiae and dark hind tarsi (only slightly darker very distal apices of hind tibiae and yellow tarsi in N. bengalensis). From N. chakrabartii sp. nov., N. piri apterae differ in the lack of dark pigment in the distal part of the hind femora (femora with dark distal patches in N. chakrabartii sp. nov.), and ANT V and VI with darker apical halves (ANT IV-VI uniformly brown in N. chakrabartii sp. nov.). Nippolachnus piri apterae differ from those in N. xitianmushanus as follows: (1) only ANT VI distal part pigmented (brown pigmented distal part of ANT IV and V and brown ANT VI in N. xitianmushanus); (2) 8–9 HT I setae (11–12 in N. xitianmushanus); (3) ANT VI/ANT III 0.52–0.60 (0.61–0.66 in N. xitianmushanus). Alate viviparous females of N. piri are most similar to N. sinensis sp. nov. and N. xitianmushanus. The three species differ from the other species in abdominal sclerotisation (poorly developed in N. bengalensis and N. chakrabartii sp. nov. and with sclerotised cross-bars on ABD VI and VII in N. micromeli). Alatae of N. piri differ from those of N. sinensis sp. nov. in having much more developed spino-pleural sclerotisation on ABD III–V, and ABD IV with marginal plates which are fused with SIPH sclerites (ABD III–V mostly with spinal sclerotisation and ABD IV without marginal sclerites in N. sinensis sp. nov.). Alatae of N. piri differ from those of N. xitianmushanus in having the following: (1) poorly developed marginal sclerites on ABD VI (well-developed in N. xitianmushanus); (2) more than 30 setae on ABD VIII (21–25 in N. xitianmushanus); (3) URS/ANT III 0.48–0.57 (0.58–0.63 in N. xitianmushanus).
Host plants. Representatives of N. piri feed on Eriobotrya japonica, Eriobotrya sp., Pyrus communis, P. pyrifolia, Pyrus sp.
Biology. In mid-May, three colonies of this species were discovered on the undersides of leaves of
Eriobotrya japonica (
Figure 4c). Each colony exhibited one fundatrix, alate viviparous female adults, and nymphs; however, apterous viviparous females were not observed. Two colonies were observed to be approached by
Camponotus japonicus (
Figure 4f), and in one colony,
Pristmyrmex punctatus (
Figure 4h) was observed. Considering the presence of alate viviparous female nymphs in all colonies and the absence of apterous viviparous females, it can be inferred that the second generations likely consist of alate viviparous females for host plant migration. From mid-May, alate viviparous females moved to another host plant species and began forming colonies on
Pyrus pyrifolia (
Figure 4b) in early to mid-June, with apterous viviparous females outnumbering them at a ratio of 9 to 1. On
Pyrus, the colonies have been attended by
Formica japonica (
Figure 4d,g). At the turn of summer and autumn in September,
Pyrus colonies of alatoid nymphs and alate viviparous females were observed. Meanwhile, in mid-October, just before returning to Eriobotrya for wintering, the number of late-born females was higher at 9 to 1 compared to June. In November, alate viviparous females that returned to the primary host produced oviparae and males through reproduction. After mating, they laid eggs in a row with consistent spacing on the undersides of leaves near the main vein of the host plant where they overwintered.
Distribution. Despite a lot of historic publications in which
N. piri is described as a part of the fauna (e.g., India), our studies indicate that this species is restricted to Japan, the Korean Peninsula, China (north-eastern part), Taiwan [
34], and Laos [
35]. This species could be present in the Russian Far East.
Remarks. Otake [
36] described the oviposition behaviour as well as the survival and hatching of the eggs and fate of young colonies [
37,
38] of
N. piri on
Rhaphiolepis umbellta (=
R. indica). However, according to Kanturski et al. [
13], this plant genus is the host plant of
N. micromeli, and therefore the results of those publications should refer to this species and not
N. piri.
Colour: in life, unknown. Mounted specimens: body very pale, only tibiae yellow with pale brown distal ends. Tarsi yellow (
Figure 13f and
Figure 15f).
Morphometric characters: HW 0.62–0.63 × ANT. ANT 0.30–0.33 × BL. ANT III with 0–1 secondary rhinarium, ANT IV always shorter than ANT V with 0–1 secondary rhinarium. ANT V as long as or shorter than ANT VI. ANT VI with 16–17 basal setae and 2–3 long, fine setae on PT (
Figure 14f). ANT VI PT 0.54–0.55 × BASE. Other antennal ratios: VI:III 0.57–0.63, V:III 0.51–0.55, IV:III 0.31–0.34. LS III 6.25–7.50 × BD III. URS 0.51–0.57 × ANT III, 0.01–1.00 × ANT VI, and 0.71–0.75 × HT II with 15–16 accessory setae. Hind tibiae setae 0.10–0.18 mm long. HT I with about 7 ventral setae (except sense peg). HT II 0.72–0.75 × ANT III and 1.13–1.32 ANT VI. Dorsal setae, 0.13–0.15 mm long. ABD VIII with 38–42 setae.
Colour: in life, unknown. Mounted specimens: head and thorax light brown. Antennae yellow with slightly darker distal end of ANT V and VI. Fore and middle legs yellow with light brown distal ends of femora and tarsi. Hind femora brown with darker distal 1/3. Hind femora yellow to light brown with darker distal ends of tibiae and darker tarsi (
Figure 16e).
Morphometric characters: HW 0.67–0.71 × ANT. ANT 0.30–0.31 × BL. ANT III with 7–8 secondary rhinaria, ANT IV shorter than ANT V with 1–2 secondary rhinaria. ANT V shorter than ANT VI with one secondary rhinarium (
Figure 17e). ANT VI with 16–17 basal setae and 2–3 long, fine setae on PT. ANT VI with PT 0.46–0.50 × BASE. Other antennal ratios: VI:III 0.67–0.69, V:III 0.57, IV:III 0.34–0.35. LS III 2.80–3.00 × BD III. URS 0.53–0.65 × ANT III, 0.78–0.94 × ANT VI, and 0.68–0.79 × HT II with about 15–16 accessory setae. Scale-like elements distributed in the very distal part of Cu1a, M1, and M2 areas of the membrane. Media one-branched with the fork in the distal part of the pterostigma length, directly under the Rs rising (
Figure 18e). Hind tibiae setae 0.06–0.180 mm long. HT I with 10 ventral setae. HT II 0.78–0.82 × ANT III and 1.15–1.19 ANT VI. Dorsal setae 0.10–0.13 mm long. Abdominal sclerotisation: ABD I with spino-pleural broken cross-band, ABD II with marginal sclerites, ABD III with marginal sclerites and spinal sclerite fused with poorly developed central abdominal sclerotisation, ABD IV with only a few small marginal scleroites at setal bases and large spinal sclerite. ABD VI without sclerites, ABD VII with residual marginal sclerites, ABD III with broken cross-band. Less sclerotised part of the patch on ABD IV and V poorly developed and rather smooth (
Figure 19e and
Figure 20e). ABD VIII with about 14–17 setae.
Material examined. Holotype. CHINA, Junnan, Kunming, 15 May 1980, Pyrus pyrifolia, one apt. viv. fem., Tieseng Zhong and Lin-Yao Wang leg., BM 2004-145 NHMUK; Paratypes. One al. viv. fem., other data as in the holotype, BM 2004-145(2) NHMUK; one al. viv. fem., one nymph, other data as in the holotype, BM 2004-145(1) DZUS; Jilin Province, 29 Sep. 2008, host unknown, one apt. viv. fem., Kim et al. leg., 08/09/01 DZUS.
Diagnosis. Apterous viviparous females of Nippolachnus sinensis sp. nov. are most similar to N. malayaensis sp. nov. in having a very pale body and appendages in which both species differ from N. bengalensis and N. chakrabartii sp. nov. Both species differ, moreover, in the chaetotaxy of ANT BASE (16–17), URS (15–16), and ABD VIII (36–42) from N. micromeli, N. piri, and N. xitianmushanus, in which the values of those characters are 20–27, 18–25, and 45–52, respectively. Nippolachnus sinensis sp. nov. can be easily distinguished from N. malayaensis sp. nov. by the following differences: (1) HW/ANT 0.62–0.63 (0.44–0.48 in N. malayaensis sp. nov.), (2) ANT VI/ANT III 0.57–0.63 (0.66–0.81 in N. malayaensis sp. nov.), (3) URS/ANT III 0.51–0.57 (0.39–0.46 in N. malayaensis sp. nov.), (4) URS/ANT VI 0.81–1.00 (0.59–0.66 in N. malayaensis sp. nov.), (5) HT II/ANT III 0.72–0.75 (0.65–0.66 in N. malayaensis sp. nov.), (6) HT II/ANT VI 1.13–1.31 (0.80–1.00 in N. malayaensis sp. nov.), (7) HT I basal length/HT I dorsal length 2.05–2.33 (1.20–1.50 in N. malayaensis sp. nov.). Alate viviparous females differ from other species in having one- branched media (N. micromeli, N. piri and N. xitianmushanus), poorly developed abdominal sclerotisation, e.g., ABD IV without marginal sclerites (present in N. piri and N. xitianmushanus), and without spino-pleural sclerites on ABD VI and VII (present on N. micromeli). The fore wing membrane of N. sinensis sp. nov. is, moreover, characterised by less developed scale-like elements distributed in the distal parts of the wing membrane.
Host plants. The species has been collected from Pyrus betulifolia.
Distribution. Nippolachnus sinensis sp. nov. is known from two localities in China, in Kunming, and from an unspecified locality in the Jilin Province.
Colour: in life, unknown. Mounted specimens: body and appendages in general pale and poorly pigmented. ANT yellow with darker distal parts of ANT IV and ANT V. ANT VI light brown with paler proximal part. Fore and middle legs pale to yellow. Hind legs pale to yellow with slightly darker distal ends of femora, dark distal ends of tibiae, and dark tarsi (
Figure 13g and
Figure 15g).
Morphometric characters: HW 0.62–0.64 × ANT. ANT 0.25–0.26 × BL. ANT IV shorter than ANT V. ANT V shorter than ANT VI. ANT VI with 25–27 basal setae and 3–4 long, fine setae on PT (
Figure 14g). ANT VI PT 0.41–0.54 × BASE. Other antennal ratios: VI:III 0.61–0.66, V:III 0.40–0.53, IV:III 0.36–0.39. LS III 5.80–6.80 × BD III. URS 0.60–0.64 × ANT III, 0.97 × ANT VI, and 0.78–0.81 × HT II with 20 accessory setae. Hind tibiae setae 0.10–0.18 mm long. HT I with 11–12 ventral setae (except sense peg). HT II 0.73–0.82 × ANT III and 1.18–1.24 ANT VI. Dorsal setae, 0.11–0.16 mm long. ABD VIII with 52 setae.
Colour: in life, unknown. Mounted specimens: head and thorax brown. Antennae brown with darker distal end of ANT III, darker halves of ANT IV–VI. Fore and middle legs yellow to light brown with slightly darker distal ends of femora and darker tarsi. Hind femora brown with darker distal 1/3. Hind femora yellow to light brown with darker distal ends of tibiae and darker tarsi (
Figure 16f).
Morphometric characters: HW 0.60–0.72 × ANT. ANT 0.27–0.30 × BL. ANT III with 4–10 secondary rhinaria, ANT IV shorter than ANT V with two secondary rhinaria. ANT V shorter than ANT VI with 1–2 secondary rhinaria (
Figure 17f). ANT VI with 15–20 basal setae and 4–6 long, fine setae on PT. ANT VI with PT 0.48–0.59 × BASE. Other antennal ratios: VI:III 0.60–0.65, V:III 0.54–0.63, IV:III 0.37–0.47. LS III 4.16–5.60 × BD III. URS 0.58–0.63 × ANT III, 0.91–1.05 × ANT VI, and 0.82–0.87 × HT II with about 33 accessory setae. Scale-like elements distributed in the distal part of Cu1a, M, and M1 areas of the membrane. Media one-branched with the fork in the middle of pterostigma length (
Figure 18f). Hind tibiae setae 0.10–0.20 mm long. HT I with 11 ventral setae. HT II 0.68–0.76 × ANT III and 1.04–1.27 ANT VI. Dorsal setae 0.13–0.16 mm long. Abdominal sclerotisation: ABD I with spino-pleural cross-band, ABD II with marginal sclerites, ABD III with marginal sclerites and small pleural sclerite fused with the rest of the central abdominal sclerotisation, ABD IV with marginal sclerites more or less fused with SIPH sclerites and large sclerotic spino-pleural cross-band. Central abdominal sclerotised patch in spinal and pleural position of ABD IV and ABD V. ABD VI without sclerites, ABD VII with marginal sclerites, ABD III with broken cross-band. Less sclerotised part of the patch on ABD IV and V cuticle forms more or less regular polygonal reticulation (
Figure 19f and
Figure 20f). ABD VIII with about 23–26 setae.
Material examined. Lectotype (here designated). CHINA, Zhejiang, Xitianmu Shan, 12 May 1975, Eriobotrya japonica, one al. viv. fem. marked with “L”, Tie-sen Zhong and Guang-xue Zhang leg., 5686-1(-1-6), 939,351 IOZ; Paralectotype. One al. viv. fem., other data the same as in Lectotype, 939,357 IOZ; other material. CHINA, Jiangxi, Jiulianshan, 08 Jun. 1975, Pyrus sp., one apt. viv. fem., one al. viv. fem., Wang Huifu leg., 6190(-1-3), 939343, 939,337 IOZ; Zhejiang, Xitianmu Shan, 13 May 1975, Eriobotrya japonica, two al. viv. fem., Tie-sen Zhong and Guang-xue Zhang leg., 5686-1(-1-1), BM 2004-145(1) NHMUK; one al. viv fem., other data as in the previous number, 5686-1(-1-2), BM 2004-145(2) NHMUK.
Diagnosis. Apterous viviparous females of N. xitianmushanus are similar to N. bengalensis, N. chakrabartii sp. nov., N. micromeli, and N. piri in pigmentation of the legs; they differ from the other apterae in having pale or only slightly pigmented very distal ends of antennae, tibiae, and tarsi. From N. bengalensis, apterae of N. xitianmushanus can be easily distinguished by the much darker distal area of hind tibiae and dark hind tarsi (only slightly darker very distal apices of hind tibiae and yellow tarsi in N. bengalensis). From N. chakrabartii sp. nov., N. xitianmushanus apterae differ in the lack of dark pigment in the distal part of the hind femora (femora with dark distal patches in N. chakrabartii sp. nov.). Apterae of N. xitianmushanus differ from N. micromeli in having the following: (1) pigmented distal part of ANT IV, V, and pigmented ANT VI (only distal end of ANT VI pigmented in N. micromeli); (2) BASE with 25–27 setae (20–22 in N. micromeli); (3) HT I basal length/HT I ventral length 0.50–0.53 (0.35–0.46 in N. micromeli). The species can be distinguished from N. piri by the following: (1) pigmented distal part of ANT IV, V, and pigmented ANT VI (only distal part of ANT VI pigmented in N. piri); (2) HT I with 11–12 setae (8–9 setae in N. piri); (3) ANT VI/ANT III 0.61–0.66 (0.52–0.60 in N. piri). Alate viviparous females of N. xitianmushanus are most similar to N. piri and N. sinensis sp. nov. The three species differ from the other species in abdominal sclerotisation (poorly developed in N. bengalensis and N. chakrabartii sp. nov. and with extra sclerotisation on ABD VI and VII in N. micromeli). Alatae of N. xitianmushanus differ from those of N. sinensis sp. nov. by having much more developed spino-pleural sclerotisation on ABD III–V, and ABD IV with marginal plates which are fused with SIPH sclerites (ABD III–V mostly with spinal sclerotisation and ABD IV without marginal sclerites in N. sinensis sp. nov.). Alatae of N. xitianmushanus differ from N. piri in having the following: (1) well-developed marginal sclerites on ABD VI (poorly developed in N. piri); (2) 21–25 setae on ABD VIII (more than 30 setae in N. piri); (3) URS/ANT III 0.58–0.63 (0.48–0.57 in N. piri).
Host plants. This species has been described from Eriobotrya japonica.
Distribution. So far, the species is known from its type locality in Xitianmu Shan, Zhejang in China.
Remarks: Probably due to its short description in Chinese,
Nippolachnus xitianmushanus has been treated as a possible synonym of
N. piri by Blackman and Eastop [
10,
11]. This status was also upheld by Remaudière and Remaudière [
19]. Comparative analyses of the type material of this and other species and the listed differences clearly show the independent status of this species, which was also corroborated in the results of the phylogenetic analyses.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












