
The Role of BmTMED6 in Female Reproduction in Silkworm, Bombyx mori

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silkworm
2.2. RNA Extraction, cDNA Synthesis, RT-PCR, and qRT-PCR Analysis
2.3. Construction of Prokaryotic Expression Vector and Purification of Recombinant Protein
2.4. Synthesis of Double-Stranded RNA (dsRNA) and Injection
2.5. Western Blotting Assay
2.6. Construction of Overexpression Vector, Cell Culture, and Transfection
2.7. Statistical Analysis and Data Presentation
3. Results
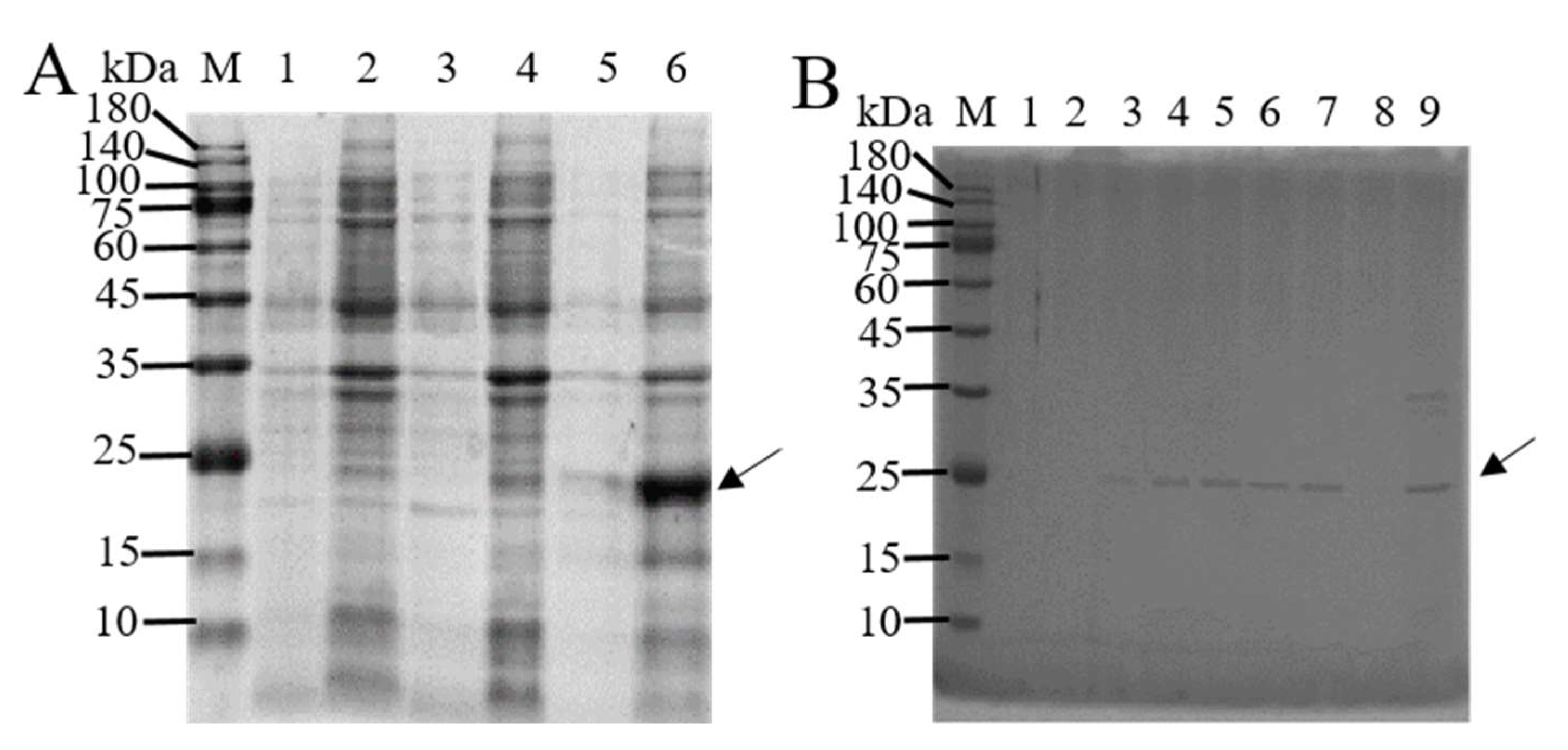
3.1. In Vitro Expression and Purification of BmTMED6
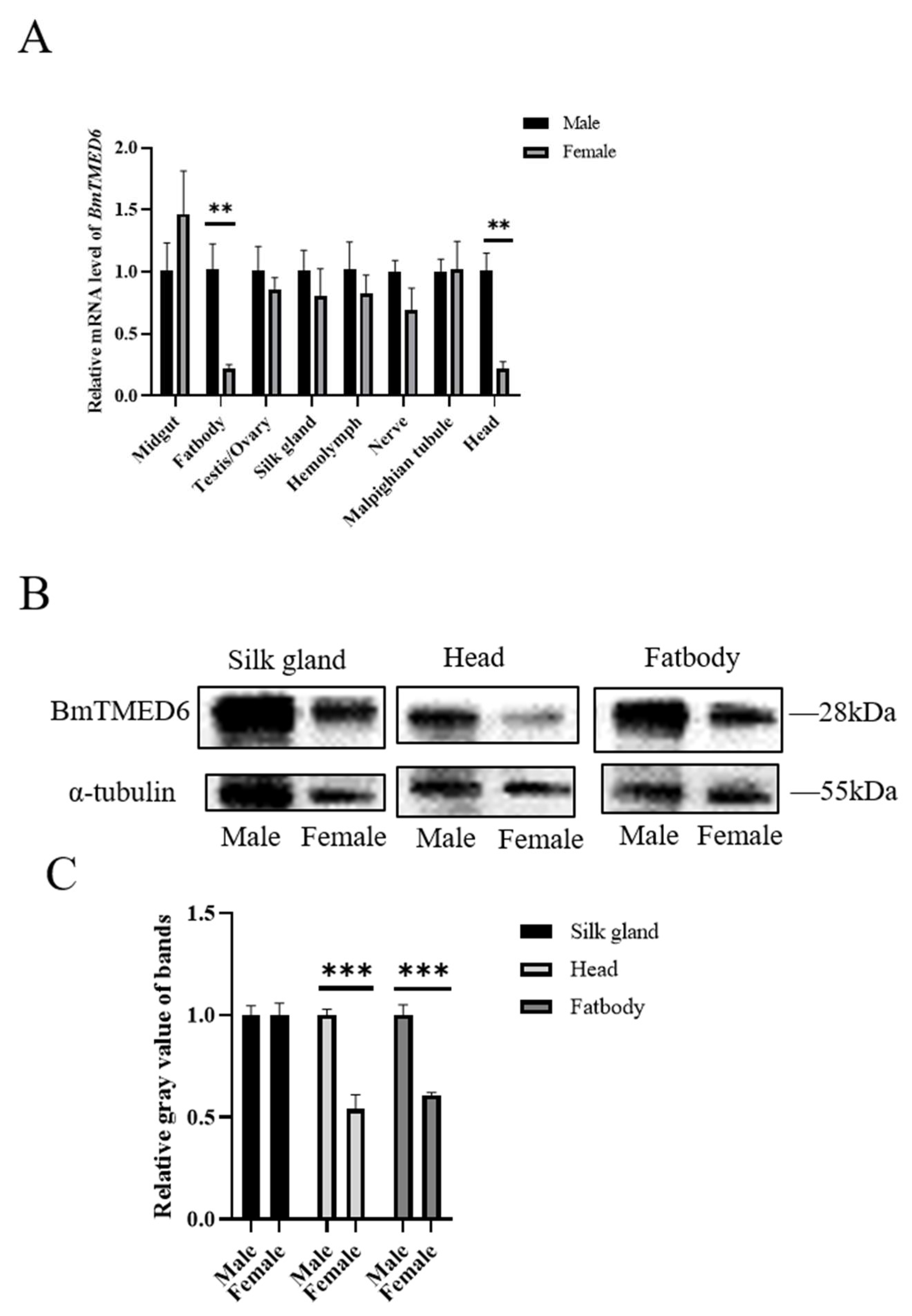
3.2. Expression of BmTMED6 in Larval Tissues of Females and Males
3.3. Expression of BmTMED6 in the Ovary during Pupal Development
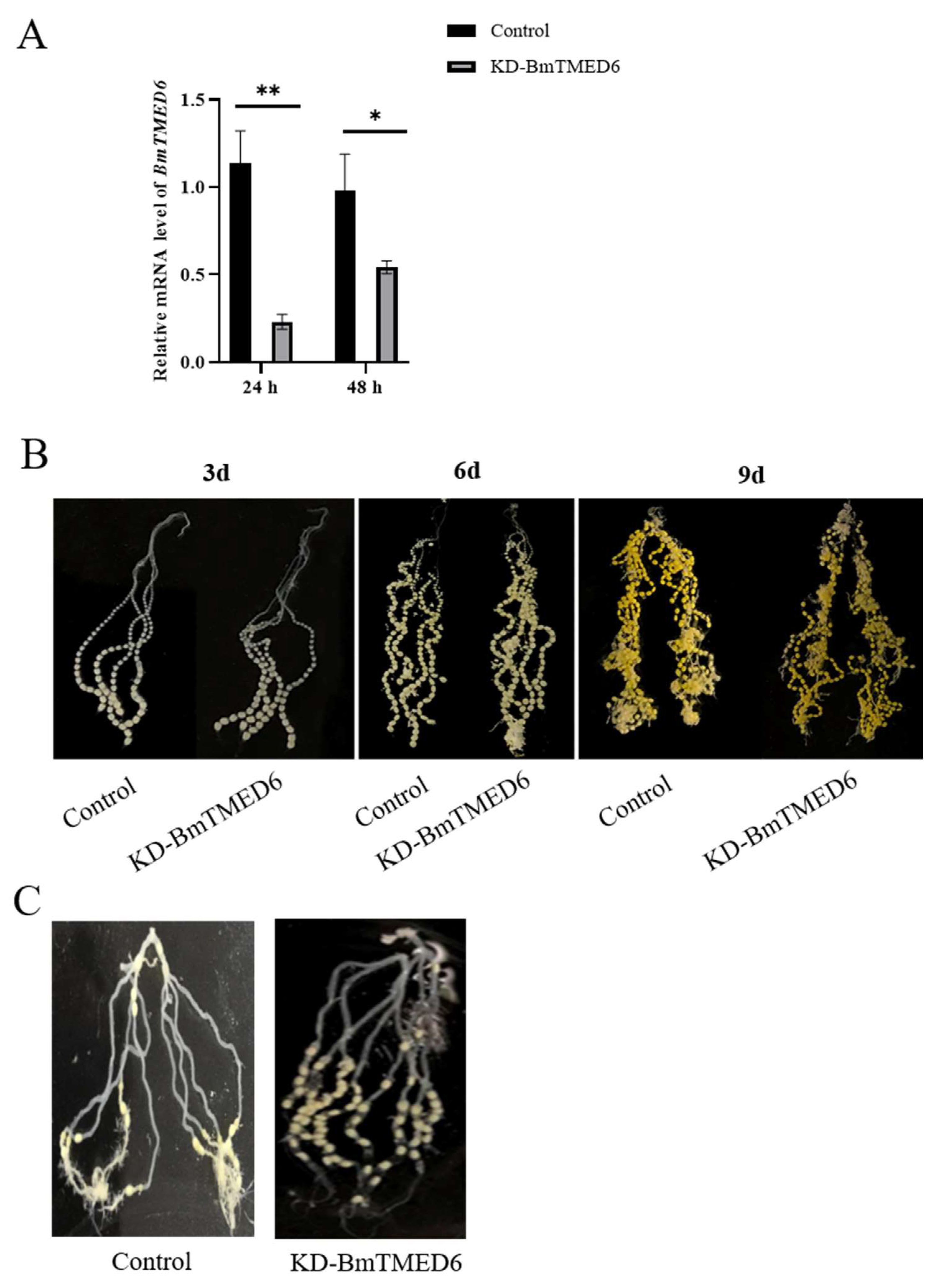
3.4. Effect of BmTMED6 Knockdown in Female Pupae
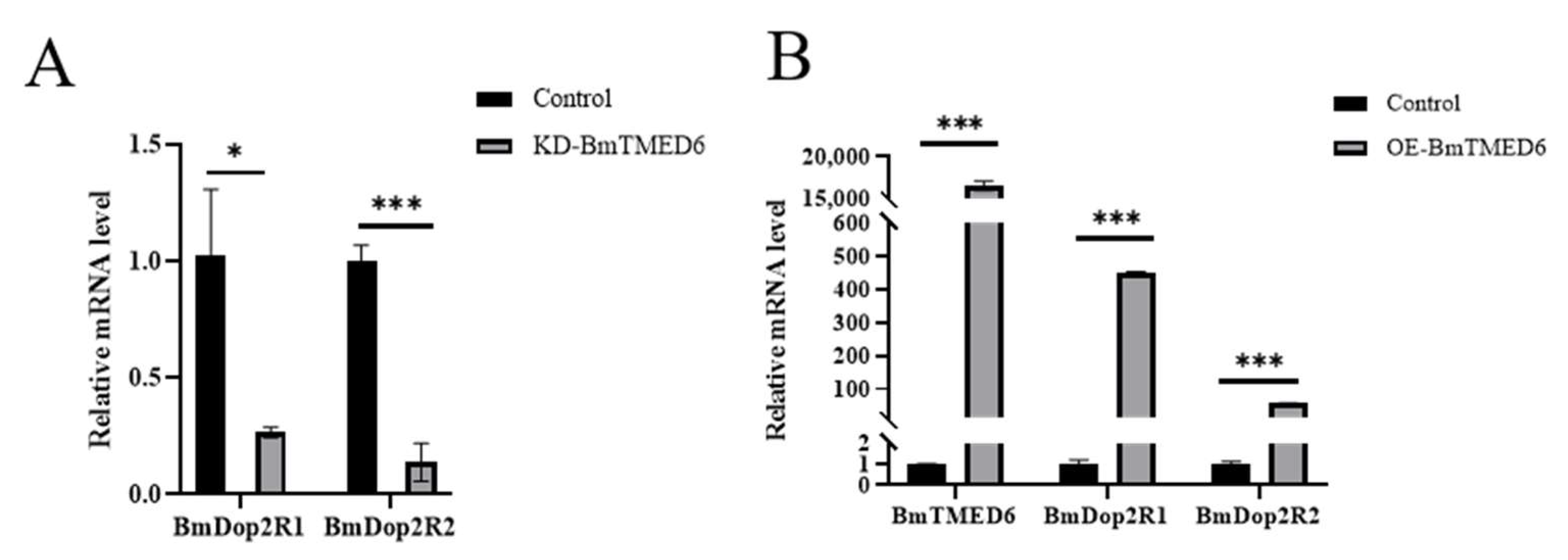
3.5. The Impact of BmTMED6 on BmDop2R Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Schuiki, I.; Volchuk, A. Diverse roles for the p24 family of proteins in eukaryotic cells. Biomol. Concepts 2012, 3, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, J.C.; Langhans, M.; Sturm, S.; Hillmer, S.; Aniento, F.; Robinson, D.G.; Marcote, M.J. Putative p24 complexes in Arabidopsis contain members of the delta and beta subfamilies and cycle in the early secretory pathway. J. Exp. Bot. 2013, 64, 3147–3167. [Google Scholar] [CrossRef] [PubMed]
- Strating, J.; Martens, G.J.M. The p24 family and selective transport processes at the ER-Golgi interface. Biol. Cell 2009, 101, 495–509. [Google Scholar] [CrossRef] [PubMed]
- Aber, R.; Chan, W.; Mugisha, S.; Jerome-Majewska, L.A. Transmembrane emp24 domain proteins in development and disease. Genet. Res. 2019, 101, e14. [Google Scholar] [CrossRef] [PubMed]
- Strating, J.R.P.M.; van Bakel, N.H.M.; Leunissen, J.A.M.; Martens, G.J.M. A comprehensive overview of the vertebrate p24 family: Identification of a novel tissue-specifically expressed member. Mol. Biol. Evol. 2009, 26, 1707–1714. [Google Scholar] [CrossRef]
- WANG, C.; GUO, Y.; LI, H.; Chen, P. Analyzing the evolution of insect TMED gene and the expression pattern of silkworm TMED gene. Chin. J. Biotechnol. 2023, 39, 4996–5013. [Google Scholar] [CrossRef]
- Connolly, D.J.; O’Neill, L.A.J.; McGettrick, A.F. The GOLD domain-containing protein TMED1 is involved in interleukin-33 signaling. J. Biol. Chem. 2013, 288, 5616–5623. [Google Scholar] [CrossRef]
- Doyle, S.L.; Husebye, H.; Connolly, D.J.; Espevik, T.; O’Neill, L.A.J.; McGettrick, A.F. The GOLD domain-containing protein TMED7 inhibits TLR4 signalling from the endosome upon LPS stimulation. Nat. Commun. 2012, 3, 707. [Google Scholar] [CrossRef]
- Zhang, X.M.; Luo, Y.L.; Li, Q.C. TMED3 promotes proliferation and migration in breast cancer cells by activating Wnt/beta-catenin signaling. Oncotargets Ther. 2020, 13, 5819–5830. [Google Scholar] [CrossRef]
- Cheng, X.Y.; Deng, X.L.; Zeng, H.P.; Zhou, T.; Li, D.Z.; Zheng, W.V. Silencing of TMED5 inhibits proliferation, migration and invasion, and enhances apoptosis of hepatocellular carcinoma cells. Adv. Clin. Exp. Med. 2023, 32, 677–688. [Google Scholar] [CrossRef]
- Wang, X.C.; Yang, R.Z.; Jadhao, S.B.; Yu, D.Z.; Hu, H.; Glynn-Cunningham, N.; Sztalryd, C.; Silver, K.D.; Gong, D.W. Transmembrane emp24 protein transport domain 6 is selectively expressed in pancreatic islets and implicated in insulin secretion and diabetes. Pancreas 2012, 41, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Carney, G.E.; Taylor, B.J. logjam encodes a predicted EMP24/GP25 protein that is required for Drosophila oviposition behavior. Genetics 2003, 164, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S.; Schwedes, C.C.; Ellis, L.L.; Grady, S.T.; Adams, R.L.; Johnson, N.; Whittington, J.R.; Carney, G.E. Drosophila melanogaster p24 trafficking proteins have vital roles in development and reproduction. Mech. Dev. 2012, 129, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Boltz, K.A.; Carney, G.E. Loss of p24 function in Drosophila melanogaster causes a stress response and increased levels of NF-kappa B-regulated gene products. Bmc Genom. 2008, 9, 212. [Google Scholar] [CrossRef] [PubMed]
- The International Silkworm Genome Consortium. The genome of a lepidopteran model insect, the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.Z.; Jiang, L.; Xia, Q.Y. Selection of reference genes for analysis of stress-responsive genes after challenge with viruses and temperature changes in the silkworm Bombyx mori. Mol. Genet. Genom. 2016, 291, 999–1004. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-delta delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pan, M.-H.; Cai, X.-J.; Liu, M.; Lv, J.; Tang, H.; Tan, J.; Lu, C. Establishment and characterization of an ovarian cell line of the silkworm, Bombyx mori. Tissue Cell 2010, 42, 42–46. [Google Scholar] [CrossRef]
- Chen, P.; Chen, P.; Li, T.; Shen, Q.; Yan, D.-F.; Zhang, L.; Chen, X.; Li, Y.; Zhao, W. Two dopamine D2-like receptor genes from the silkworm (Bombyx mori) and their evolutionary history in metazoan. Sci. Rep. 2017, 7, 6848. [Google Scholar] [CrossRef]
- Navarro, K.G.; Chamberlin, H.M. Genetic characterization of C. elegans TMED genes. Dev. Dyn. 2023, 252, 1149–1161. [Google Scholar] [CrossRef]
- Wen, C.H.; Greenwald, I. p24 proteins and quality control of LIN-12 and GLP-1 trafficking in Caenorhabditis elegans. J. Cell Biol. 1999, 145, 1165–1175. [Google Scholar] [CrossRef] [PubMed]
- Boltz, K.A.; Ellis, L.L.; Carney, G.E. Drosophila melanogaster p24 genes have developmental, tissue-specific, and sex-specific expression patterns and functions. Dev. Dyn. 2007, 236, 544–555. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.O.; Boswell, S.A.; Seo, J.S.; Lee, S.W. Novel oxidative stress-responsive gene ERS25 functions as a regulator of the heat-shock and cell death response. J. Biol. Chem. 2008, 283, 13063–13069. [Google Scholar] [CrossRef] [PubMed]
- Sasako, T.; Ohsugi, M.; Kubota, N.; Itoh, S.; Okazaki, Y.; Terai, A.; Kubota, T.; Yamashita, S.; Nakatsukasa, K.; Kamura, T.; et al. Hepatic Sdf2l1 controls feeding-induced ER stress and regulates metabolism. Nat. Commun. 2019, 10, 947. [Google Scholar] [CrossRef]
- Park, H.; Seo, S.K.; Sim, J.R.; Hwang, S.J.; Kim, Y.J.; Shin, D.H.; Jang, D.G.; Noh, S.H.; Park, P.G.; Ko, S.H.; et al. TMED3 complex mediates ER stress-associated secretion of CFTR, pendrin, and SARS-CoV-2 spike. Adv. Sci. 2022, 9, 2105320. [Google Scholar] [CrossRef] [PubMed]
- Tashima, Y.; Hirata, T.; Maeda, Y.; Murakami, Y.; Kinoshita, T. Differential use of p24 family members as cargo receptors for the transport of glycosylphosphatidylinositol-anchored proteins and Wnt1. J. Biochem. 2022, 171, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Middleton, C.A.; Nongthomba, U.; Parry, K.; Sweeney, S.T.; Sparrow, J.C.; Elliott, C.J.H. Neuromuscular organization and aminergic modulation of contractions in the Drosophila ovary. BMC Biol. 2006, 4, 17. [Google Scholar] [CrossRef]
- Brewer, J.C.; Olson, A.C.; Collins, K.M.; Koelle, M.R. Serotonin and neuropeptides are both released by the HSN command neuron to initiate C. elegans egg laying. PLoS Genet. 2019, 15, 1007896. [Google Scholar] [CrossRef]
- Blenau, W.; Baumann, A. Molecular and pharmacological properties of insect biogenic amine receptors: Lessons from Drosophila melanogaster and Apis mellifera. Arch. Insect Biochem. 2001, 48, 13–38. [Google Scholar] [CrossRef]
- Vallone, D.; Picetti, R.; Borrelli, E. Structure and function of dopamine receptors. Neurosci. Biobehav. Rev. 2000, 24, 125–132. [Google Scholar] [CrossRef]
- Bai, H.; Palli, S.R. Identification of G protein-coupled receptors required for vitellogenin uptake into the oocytes of the red flour beetle, Tribolium castaneum. Sci. Rep. 2016, 6, 27648. [Google Scholar] [CrossRef] [PubMed]
- Azanchi, R.; Kaun, K.R.; Heberlein, U. Competing dopamine neurons drive oviposition choice for ethanol in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, 21153–21158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.X.; Rogulja, D.; Crickmore, M.A. Dopaminergic circuitry underlying mating drive. Neuron 2016, 91, 168–181. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.B.; Wang, Y.F.; Reiser, G. P24A, a type I transmembrane protein, controls ARF1-dependent resensitization of protease-activated receptor-2 by influence on receptor trafficking. J. Biol. Chem. 2007, 282, 30246–30255. [Google Scholar] [CrossRef] [PubMed]
- Fadista, J.; Vikman, P.; Laakso, E.O.; Mollet, I.G.; Esguerra, J.L.; Taneera, J.; Storm, P.; Osmark, P.; Ladenvall, C.; Prasad, R.B.; et al. Global genomic and transcriptomic analysis of human pancreatic islets reveals novel genes influencing glucose metabolism. Proc. Natl. Acad. Sci. USA 2014, 111, 13924–13929. [Google Scholar] [CrossRef] [PubMed]
- Rubí, B.; Ljubicic, S.; Pournourmohammadi, S.; Carobbio, S.; Armanet, M.; Bartley, C.; Maechler, P. Dopamine D2-like receptors are expressed in pancreatic beta cells and mediate inhibition of insulin secretion. J. Biol. Chem. 2005, 280, 36824–36832. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.M.; Ren, L.L.; Zhong, X.Q.; Ding, Y.Q.; Liu, T.; Liu, Z.H.; Yang, X.H.; Cui, L.J.; Yang, L.J.; Fan, Y.Y.; et al. D2-like receptors mediate dopamine-inhibited insulin secretion via ion channels in rat pancreatic beta-cells. Front. Endocrinol. 2020, 11, 152. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Development Time (d) | Survival Rate | Mating Success Rate | Proportion of Female Moths That Ovulate |
|---|---|---|---|---|
| Control | 9.5 | 100% | 100% | 100% |
| KD-BmTMED6 | 9.5 | 100% | 100% | 100% |
| Group | Number of Laid Eggs | Number of Unlaid Eggs | Total Egg Production |
|---|---|---|---|
| Control | 226.38 ± 32.513 a | 14.92 ± 7.205 d | 241.31 ± 32.778 a |
| KD-BmTMED6 | 141.00 ± 49.346 b | 70.85 ± 21.385 c | 211.85 ± 36.792 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Hu, Z.; Guo, Y.; Xiao, W.; Zhang, Y.; Zhou, A.; Chen, P. The Role of BmTMED6 in Female Reproduction in Silkworm, Bombyx mori. Insects 2024, 15, 103. https://doi.org/10.3390/insects15020103
Wang C, Hu Z, Guo Y, Xiao W, Zhang Y, Zhou A, Chen P. The Role of BmTMED6 in Female Reproduction in Silkworm, Bombyx mori. Insects. 2024; 15(2):103. https://doi.org/10.3390/insects15020103
Chicago/Turabian StyleWang, Chunyang, Zunmei Hu, Yu Guo, Wenfu Xiao, Youhong Zhang, Anlian Zhou, and Ping Chen. 2024. "The Role of BmTMED6 in Female Reproduction in Silkworm, Bombyx mori" Insects 15, no. 2: 103. https://doi.org/10.3390/insects15020103
APA StyleWang, C., Hu, Z., Guo, Y., Xiao, W., Zhang, Y., Zhou, A., & Chen, P. (2024). The Role of BmTMED6 in Female Reproduction in Silkworm, Bombyx mori. Insects, 15(2), 103. https://doi.org/10.3390/insects15020103