Oviposition Preference and Developmental Performance of Drosophila suzukii on Different Cherry Cultivars

 ,
,  and
and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Colony
2.2. Test Fruits
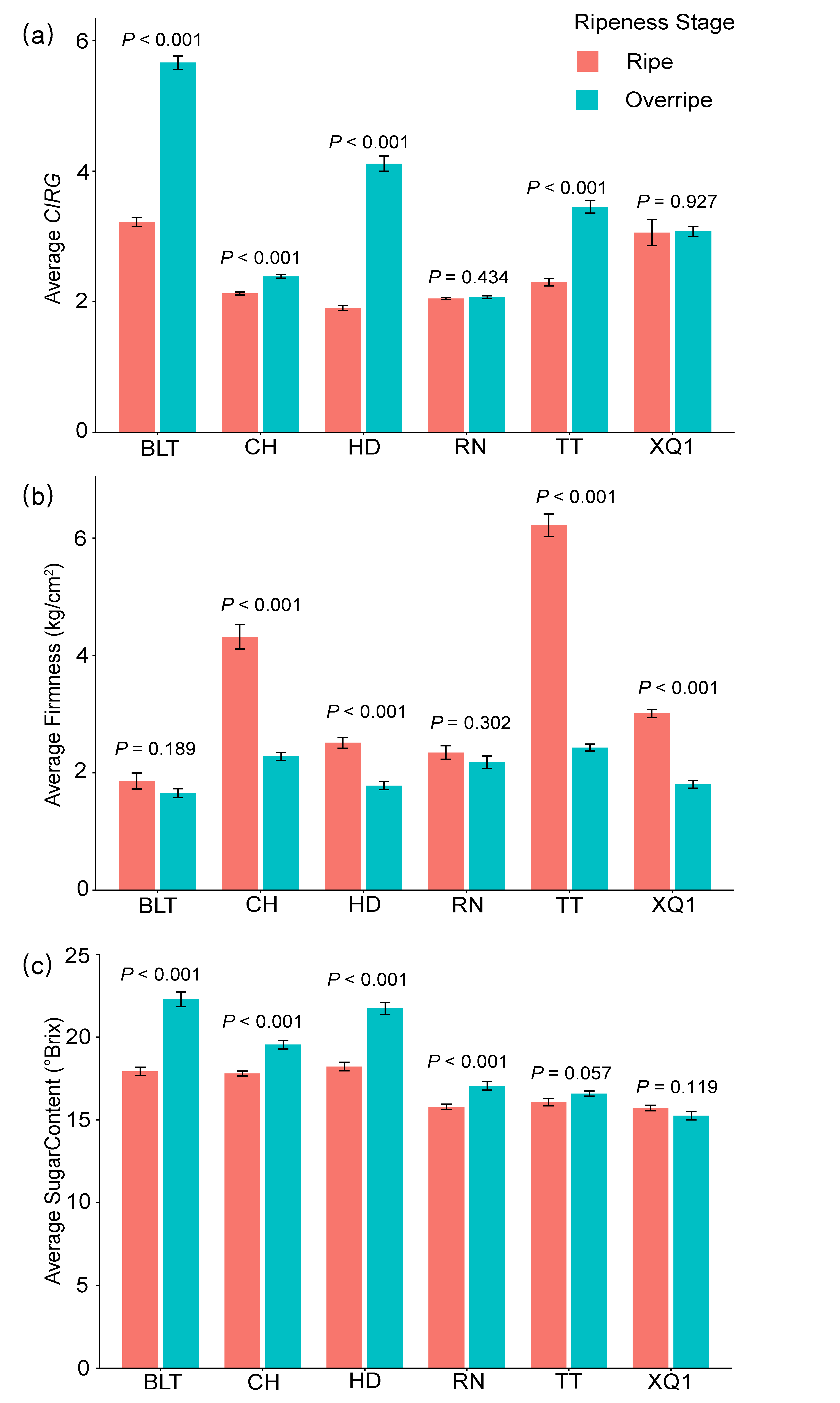
2.3. Measurement of Cherry Fruit Color, Firmness, and Sugar Content
2.3.1. Fruit Color
2.3.2. Fruit Firmness
2.3.3. Fruit Sugar Content
2.4. Oviposition Preference Test of Drosophila suzukii on Fruits of Different Cherry Cultivars
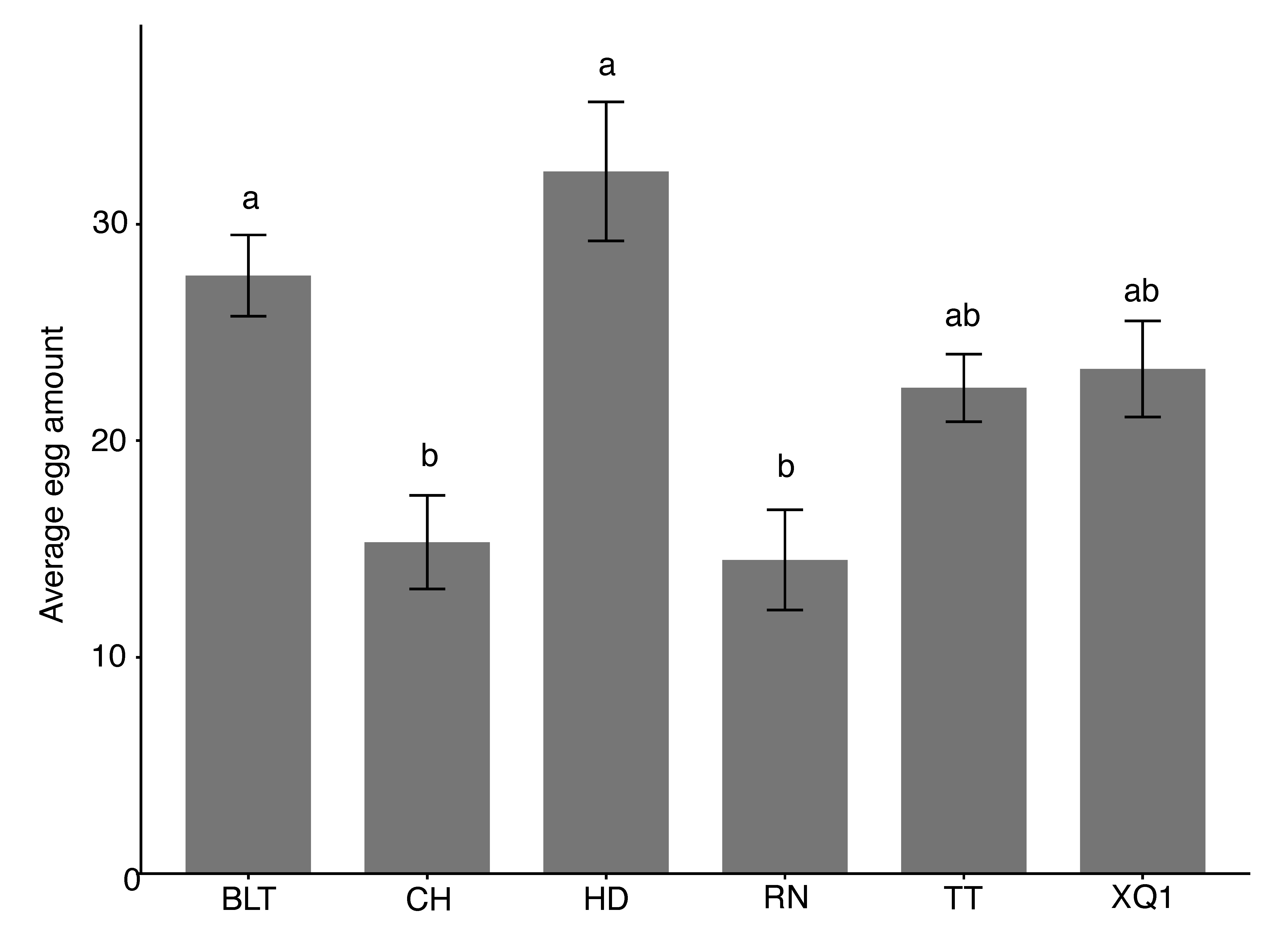
2.4.1. Non-Choice Oviposition
2.4.2. Choice Oviposition
2.5. Measurement of Growth and Development Parameters of Drosophila suzukii on Different Cherry Cultivars
2.5.1. Development Periods
2.5.2. Other Growth and Development Parameters
2.6. Data Analysis
3. Results
3.1. Physiological Parameters of Different Cherry Cultivars
3.2. Oviposition of Drosophila suzukii on Different Cherry Cultivars
3.2.1. Non-Choice Oviposition
3.2.2. Choice Oviposition
3.3. Growth and Development Parameters of Drosophila suzukii on Fruits of Different Cherry Cultivars
3.4. Correlation Analysis of Choice Oviposition, Developmental Parameters, and Fruit Physiological Parameters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asplen, M.K.; Anfora, G.; Biondi, A.; Choi, D.-S.; Chu, D.; Daane, K.M.; Gibert, P.; Gutierrez, A.P.; Hoelmer, K.A.; Hutchison, W.D.; et al. Invasion biology of spotted wing drosophila (Drosophila suzukii): A global perspective and future priorities. J. Pest Sci. 2015, 88, 469–494. [Google Scholar] [CrossRef]
- Rota-Stabelli, O.; Blaxter, M.; Anfora, G. Drosophila suzukii. Curr. Biol. 2013, 23, R8–R9. [Google Scholar] [CrossRef]
- Knapp, L.; Mazzi, D.; Finger, R. The economic impact of Drosophila suzukii: Perceived costs and revenue losses of Swiss cherry, plum and grape growers. Pest Manag. Scie. 2021, 77, 978–1000. [Google Scholar] [CrossRef]
- Mazzi, D.; Bravin, E.; Meraner, M.; Finger, R.; Kuske, S. Economic impact of the introduction and establishment of Drosophila suzukii on sweet cherry production in Switzerland. Insects 2017, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- De Ros, G.; Grassi, A.; Pantezzi, T. Recent trends in the economic impact of Drosophila suzukii. In Drosophila suzukii Management; Garcia, F.R.M., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 11–27. ISBN 978-3-030-62691-4. [Google Scholar]
- Zhao, X.; Yan, M.; Ding, Y.; Huo, Y.; Yuan, Z. Characterization and comparative analysis of the complete chloroplast genome sequence from Prunus avium “Summit”. PeerJ 2019, 7, e8210. [Google Scholar] [CrossRef]
- Ceccarelli, D.; Antonucci, F.; Costa, C.; Talento, C.; Ciccoritti, R. An artificial class modelling approach to identify the most largely diffused cultivars of sweet cherry (Prunus avium L.) in Italy. Food Chem. 2020, 333, 127515. [Google Scholar] [CrossRef]
- Kappel, F.; Granger, A.; Hrotkó, K.; Schuster, M. Cherry. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer US: Boston, MA, USA, 2012; pp. 459–504. ISBN 978-1-4419-0762-2. [Google Scholar]
- FAS Beijing; USDA Foreign Agriculture Service. China: Stone Fruit Annual. 2024. Available online: https://fas.usda.gov/data/china-stone-fruit-annual-8 (accessed on 4 December 2024).
- Overview of the development of the cherry industry in China. In Proceedings of the 9th International Cherry Symposium, Beijing, China, 21–25 May 2023.
- Beers, E.H.; Van Steenwyk, R.A.; Shearer, P.W.; Coates, W.W.; Grant, J.A. Developing Drosophila suzukii Management Programs for Sweet Cherry in the Western United States. Pest Manag. Sci. 2011, 67, 1386–1395. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wang, X.; Wang, J.; Zhu, D. Occurrence, damage, and control strategies of spotted wing drosophila on sweet cherry, blueberry, and other fruit Trees. Deciduous Fruits 2014, 46, 1–3. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, K.; Yan, G.; Guo, X.; Wang, J.; Zhang, X.; Zhou, Y. Research review on spotted wing drosophila (Drosophila suzukii). J. Fruit Sci. 2014, 31, 717–721+750. (In Chinese) [Google Scholar] [CrossRef]
- Yang, F.; Wang, Z.; Sun, A.; Yu, G.; Wang, G. Occurrence dynamics of spotted wing drosophila and comparison of different bait solutions in a cherry orchard in Beijing. China Fruits 2022, 6, 66–70. (In Chinese) [Google Scholar] [CrossRef]
- Little, C.M.; Chapman, T.W.; Hillier, N.K. Effect of color and contrast of highbush blueberries to host-finding behavior by Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2018, 47, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Little, C.M.; Dixon, P.L.; Chapman, T.W.; Hillier, N.K. Role of fruit characters and colour on host selection of boreal fruits and berries by Drosophila suzukii (Diptera: Drosophilidae). Can. Entomol. 2020, 152, 546–562. [Google Scholar] [CrossRef]
- Bolton, L.G.; Piñero, J.C.; Barrett, B.A. Olfactory cues from host- and non-host plant odor influence the behavioral responses of adult Drosophila suzukii (Diptera: Drosophilidae) to visual cues. Environ. Entomol. 2021, 50, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Entling, W.; Anslinger, S.; Jarausch, B.; Michl, G.; Hoffmann, C. Berry Skin resistance explains oviposition preferences of Drosophila suzukii at the level of grape cultivars and single berries. J. Pest Sci. 2019, 92, 477–484. [Google Scholar] [CrossRef]
- Fragnière, A.-L.; Bacher, S.; Kehrli, P. Identifying candidate host plants for trap cropping against Drosophila suzukii in vineyards. J. Pest Sci. 2024, 97, 1975–1991. [Google Scholar] [CrossRef]
- Zhang, X.; Bi, Y.; Fang, Q.; Wang, F.; Ye, G. Tropism behavior analysis of two drosophila species to fruits and volatiles of strawberry at different mature stage. Chin. J. Biol. Control. 2023, 39, 813–823. (In Chinese) [Google Scholar] [CrossRef]
- Gao, H.; Wang, Y.; Ma, N.; Chen, H.; Zhai, Y.; Dang, H.; Yu, Y. Oviposition selection of Drosophila suzukii (Diptera: Drosophilidae) on four cherry cultivars and its relationship to the physiological characteristics of cherry fruits. Acta Entomologica Sinica 2017, 60, 328–334. (In Chinese) [Google Scholar] [CrossRef]
- Yan, G.; Zhang, K. Major Cherry Varieties in China, 1st ed.; China Agriculture Press: Beijing, China, 2022; ISBN 978-7-109-29285-7. (In Chinese) [Google Scholar]
- Zhang, X.; Jiu, S.; Xu, Y.; Lv, Z.; Liu, R.; Wang, S.; Zhang, C. Evaluation and study on fruit traits of sweet cherry cultivars (strains). J. Plant Genet. Resour. 2024, 25, 1803–1814. (In Chinese) [Google Scholar] [CrossRef]
- Liu, B.; Ren, L.; Zhan, G.; Li, B.; Chen, N.; Wang, X. An Artificial diet and rearing method for mass rearing of spotted wing drosophila. China Patent Application Number CN201910882663.3, 18 September 2019. [Google Scholar]
- CIE. Colorimetry—Part 4: CIE 1976 Lab Colour Space; CIE: Vienna, Austria, 1976. [Google Scholar]
- Markovic, I.; Ilic, J.; Markovic, D.; Simonovic, V.; Kosanic, N. Color measurement of food products using CIE L* a* b* and RGB color space. J. Hyg. Eng. Des. 2020, 7, 50–53. [Google Scholar]
- Wrolstad, R.E.; Durst, R.W.; Lee, J. Tracking color and pigment changes in anthocyanin products. Trends Food Sci. Tech. 2005, 16, 423–428. [Google Scholar] [CrossRef]
- Wu, L.; Wang, F.; Sha, R.; Li, X.; Yu, K.; Feng, J. The Effect of N and KH2PO4 on skin color, sugars, and organic acids of “Flame Seedless” grape. Agronomy 2023, 13, 902. [Google Scholar] [CrossRef]
- Carreño, J.; Martínez, A.; Almela, L.; Fernández-López, J.A. Proposal of an index for the objective evaluation of the colour of red table grapes. Food Res. Int. 1995, 28, 373–377. [Google Scholar] [CrossRef]
- Fernández-López, J.A.; Almela, L.; Muñoz, J.A.; Hidalgo, V.; Carreño, J. Dependence between colour and individual anthocyanin content in ripening grapes. Food Res. Int. 1998, 31, 667–672. [Google Scholar] [CrossRef]
- Jaywant, S.A.; Singh, H.; Arif, K.M. Sensors and instruments for Brix measurement: A review. Sensors 2022, 22, 2290. [Google Scholar] [CrossRef]
- Little, C.M.; Rizzato, A.R.; Charbonneau, L.; Chapman, T.; Hillier, N.K. Color preference of the spotted wing drosophila, Drosophila suzukii. Sci. Rep. 2019, 9, 16051. [Google Scholar] [CrossRef]
- Kinjo, H.; Kunimi, Y.; Ban, T.; Nakai, M. Oviposition efficacy of Drosophila suzukii (Diptera: Drosophilidae) on different cultivars of blueberry. J. Econ. Entom. 2013, 106, 1767–1771. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef]
- Hu, Q.; Gao, H. Oviposition selection of Drosophila suzukii (Diptera: Drosophilidae) on grape cultivars and the relationship to physiological characteristics of grape fruits. North. Hortic. 2021, 2, 42–46. (In Chinese) [Google Scholar]
- Kamiyama, M.T.; Guédot, C. Varietal and developmental susceptibility of tart cherry (Rosales: Rosaceae) to Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entom. 2019, 112, 1789–1797. [Google Scholar] [CrossRef]
- Cai, P.; Song, Y.; Yi, C.; Zhang, Q.; Xia, H.; Lin, J.; Zhang, H.; Yang, J.; Ji, Q.; Chen, J. Potential host fruits for Drosophila suzukii: Olfactory and oviposition preferences and suitability for development. Entomol. Exp. Appl. 2019, 167, 880–890. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Egg–Larva Period (d) | Pupal Period (d) | Female Ratio | Hatching Rate | Pupation Rate | Eclosion Rate | Survival Rate |
|---|---|---|---|---|---|---|---|
| HD | 4.98 ± 0.009 a | 4.81 ± 0.038 | 0.58 ± 0.035 | 0.87 ± 0.014 | 0.87 ± 0.014 | 0.81 ± 0.009 | 0.62 ± 0.019 |
| TT | 4.97 ± 0.012 ab | 4.83 ± 0.037 | 0.67 ± 0.025 | 0.86 ± 0.009 | 0.84 ± 0.011 | 0.74 ± 0.024 | 0.53 ± 0.022 |
| XQ1 | 4.95 ± 0.016 ab | 4.70 ± 0.033 | 0.64 ± 0.035 | 0.89 ± 0.013 | 0.82 ± 0.020 | 0.78 ± 0.017 | 0.57 ± 0.024 |
| CH | 4.89 ± 0.023 bc | 4.78 ± 0.034 | 0.64 ± 0.035 | 0.85 ± 0.012 | 0.80 ± 0.025 | 0.75 ± 0.026 | 0.51 ± 0.030 |
| BLT | 4.92 ± 0.020 ab | 4.81 ± 0.042 | 0.59 ± 0.028 | 0.88 ± 0.019 | 0.86 ± 0.014 | 0.76 ± 0.024 | 0.58 ± 0.032 |
| RN | 4.82 ± 0.028 c | 4.88 ± 0.026 | 0.62 ± 0.062 | 0.83 ± 0.017 | 0.81 ± 0.015 | 0.76 ± 0.022 | 0.51 ± 0.024 |
| Egg Count | Egg–Larva Period | Pupal Period | Female Ratio | Hatching Rate | Pupation Rate | Eclosion Rate | Survival Rate | L* | a* | b* | CIRG | Firmness | Sugar Content | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Egg Count | 0.071 | 0.700 | 0.204 | 0.081 | 0.007 | 0.119 | 0.002 | 0.030 | 0.559 | 0.019 | 0.055 | 0.126 | 0.283 | |
| Egg–Larva Period | 0.774 | 0.336 | 0.978 | 0.087 | 0.190 | 0.381 | 0.119 | 0.021 | 0.548 | 0.059 | 0.334 | 0.642 | 0.844 | |
| Pupal Period | −0.203 | −0.480 | 0.792 | 0.079 | 0.786 | 0.528 | 0.535 | 0.421 | 0.229 | 0.532 | 0.931 | 0.486 | 0.670 | |
| Female Ratio | −0.604 | −0.015 | −0.139 | 0.588 | 0.175 | 0.106 | 0.125 | 0.689 | 0.145 | 0.508 | 0.276 | 0.063 | 0.073 | |
| Hatching Rate | 0.758 | 0.749 | −0.761 | −0.282 | 0.312 | 0.217 | 0.056 | 0.047 | 0.895 | 0.055 | 0.196 | 0.097 | 0.777 | |
| Pupation Rate | 0.933 ** | 0.620 | 0.144 | −0.635 | 0.500 | 0.283 | 0.036 | 0.079 | 0.295 | 0.044 | 0.035 | 0.248 | 0.157 | |
| Eclosion Rate | 0.703 | 0.441 | −0.327 | −0.721 | 0.591 | 0.527 | 0.032 | 0.513 | 0.855 | 0.484 | 0.619 | 0.115 | 0.544 | |
| Survival Rate | 0.966 ** | 0.704 | −0.321 | −0.695 | 0.801 | 0.840 * | 0.850 * | 0.082 | 0.621 | 0.061 | 0.121 | 0.057 | 0.311 | |
| L* | −0.855 * | −0.880 * | 0.409 | 0.210 | −0.818* | −0.761 | −0.338 | −0.756 | 0.932 | <0.001 | 0.045 | 0.332 | 0.449 | |
| a* | −0.303 | 0.311 | −0.578 | 0.671 | 0.070 | −0.515 | −0.097 | −0.258 | 0.045 | 0.678 | 0.221 | 0.306 | 0.256 | |
| b* | −0.886 * | −0.795 | 0.323 | 0.341 | −0.801 | −0.824 * | −0.359 | −0.790 | 0.984 *** | 0.218 | 0.012 | 0.223 | 0.321 | |
| CIRG | 0.802 | 0.481 | −0.046 | −0.533 | 0.612 | 0.844 * | 0.260 | 0.700 | −0.821 * | −0.587 | −0.908 * | 0.141 | 0.147 | |
| Firmness | −0.694 | −0.243 | 0.358 | 0.788 | −0.733 | −0.560 | −0.709 | −0.798 | 0.483 | 0.506 | 0.585 | −0.675 | 0.332 | |
| Sugar Content | 0.527 | 0.104 | 0.223 | −0.771 | 0.150 | 0.657 | 0.315 | 0.501 | −0.387 | −0.552 | −0.492 | 0.668 | −0.483 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Sun, H.; Wang, Z.; Xie, J.; He, J.; Qiao, G.; Wang, J.; Wang, Y.; Wang, S. Oviposition Preference and Developmental Performance of Drosophila suzukii on Different Cherry Cultivars. Insects 2024, 15, 984. https://doi.org/10.3390/insects15120984
Yang F, Sun H, Wang Z, Xie J, He J, Qiao G, Wang J, Wang Y, Wang S. Oviposition Preference and Developmental Performance of Drosophila suzukii on Different Cherry Cultivars. Insects. 2024; 15(12):984. https://doi.org/10.3390/insects15120984
Chicago/Turabian StyleYang, Fan, Haikuan Sun, Zehua Wang, Jingxia Xie, Jingyan He, Guanghang Qiao, Jing Wang, Yuyu Wang, and Shanning Wang. 2024. "Oviposition Preference and Developmental Performance of Drosophila suzukii on Different Cherry Cultivars" Insects 15, no. 12: 984. https://doi.org/10.3390/insects15120984
APA StyleYang, F., Sun, H., Wang, Z., Xie, J., He, J., Qiao, G., Wang, J., Wang, Y., & Wang, S. (2024). Oviposition Preference and Developmental Performance of Drosophila suzukii on Different Cherry Cultivars. Insects, 15(12), 984. https://doi.org/10.3390/insects15120984