
Determination of Larval Instars of Dastarcus helophoroides (Coleoptera: Bothrideridae) Using Head Capsule Width Frequency Distribution

 , and
, and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Head Capsule Measurement
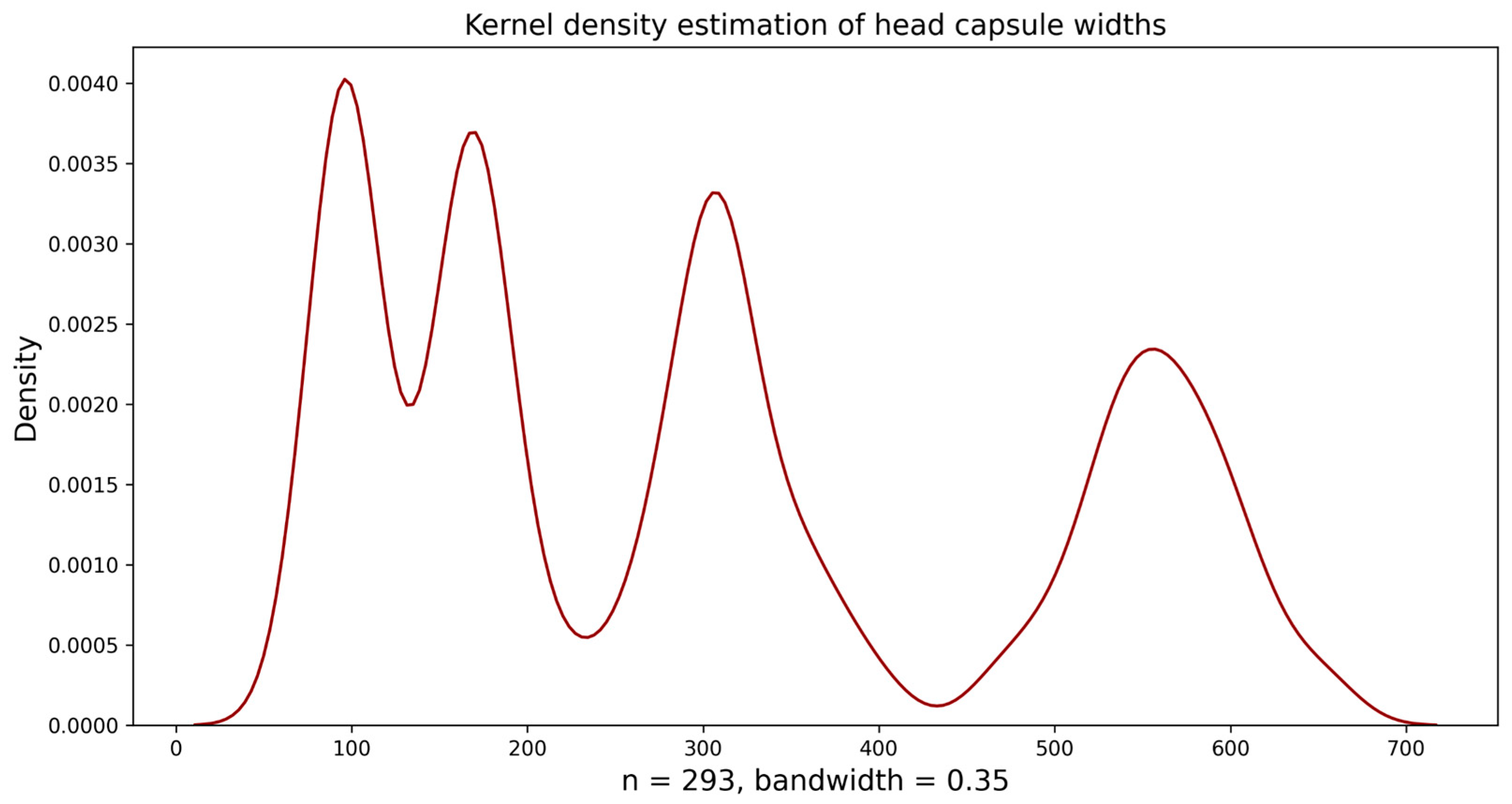
2.3. Analysis of Head Capsule Widths
3. Results
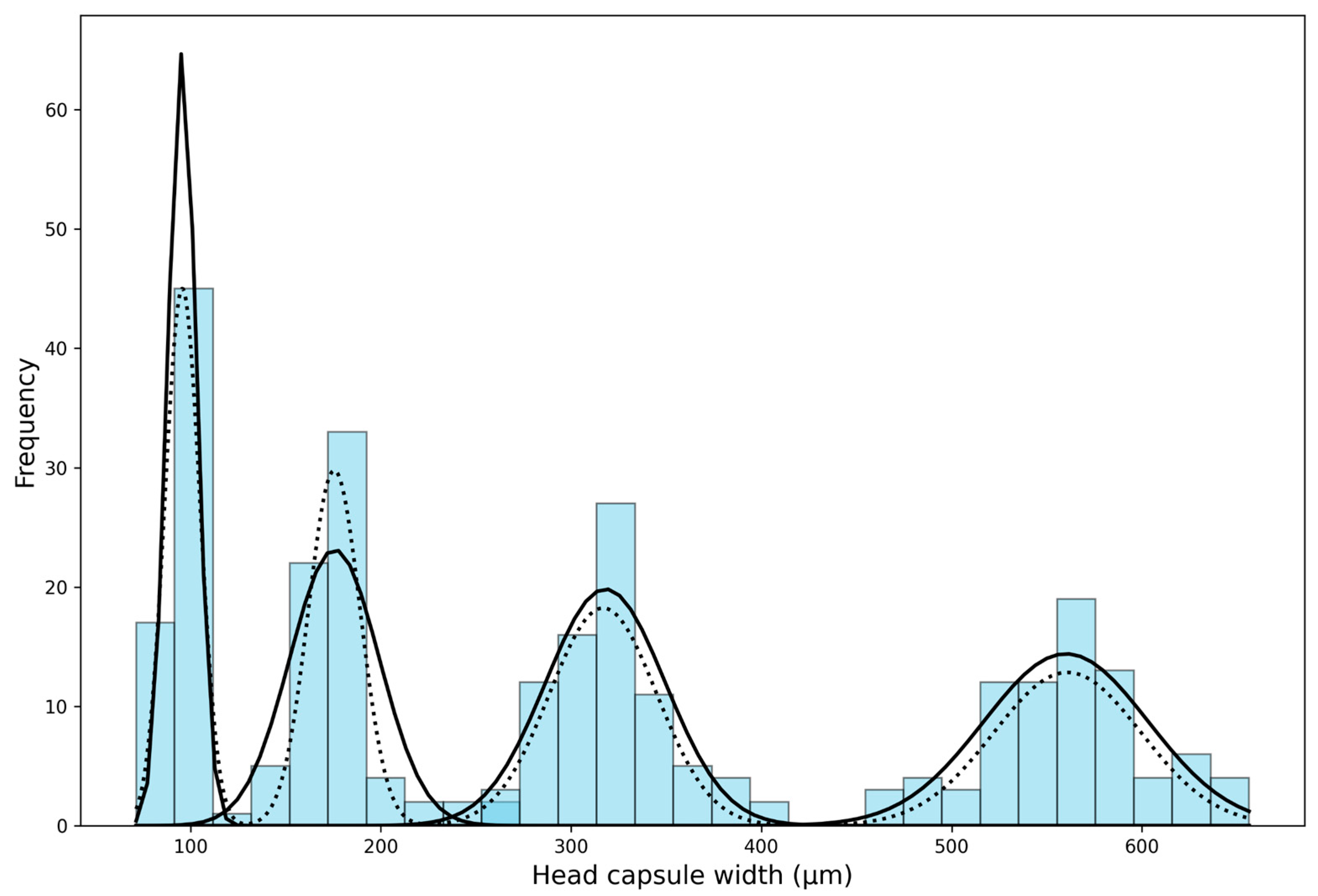
3.1. Instar-Wise Head Capsule Widths (Observed)
3.2. Instar-Wise Head Capsule Widths (Theoretical)
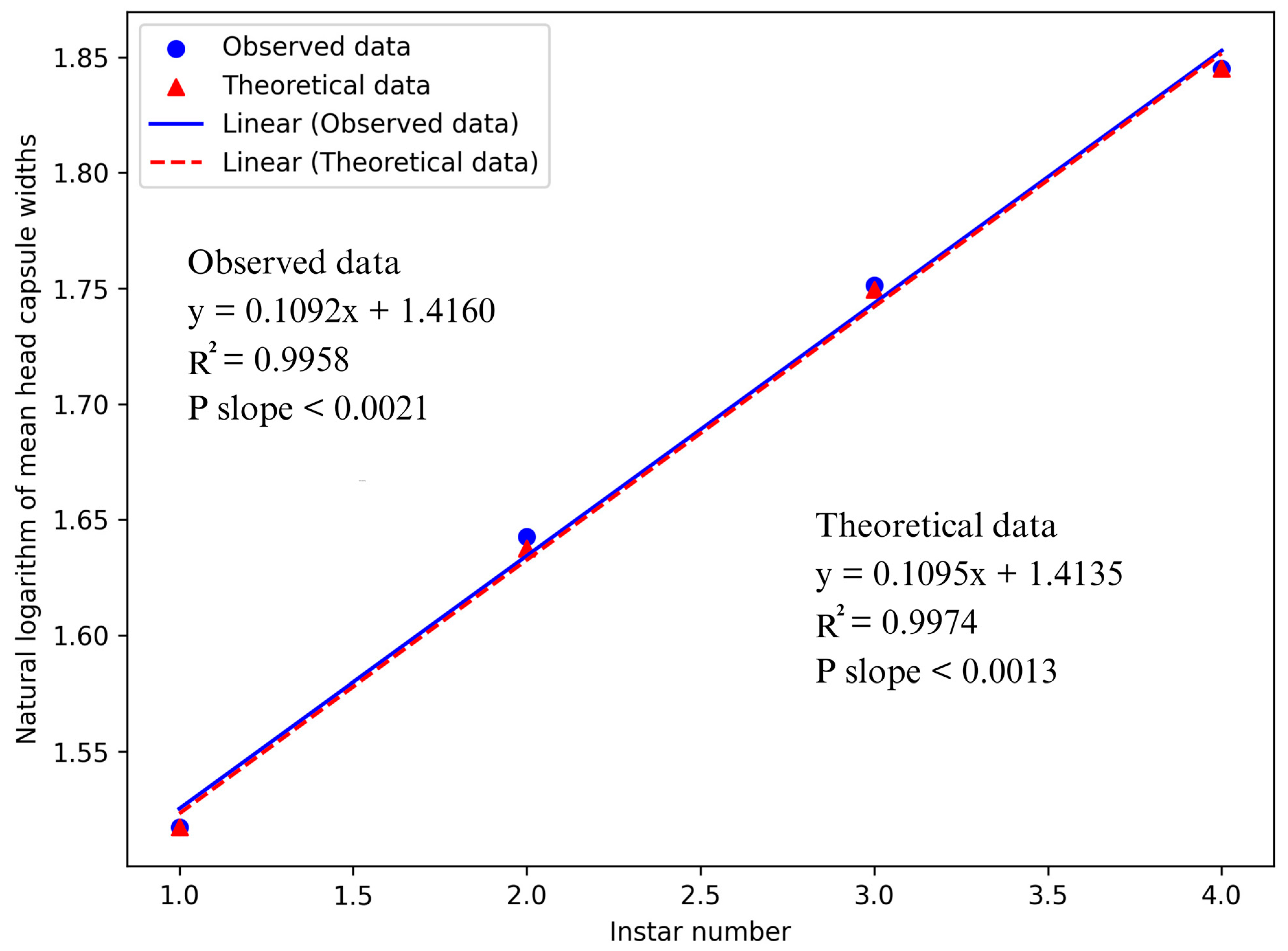
3.3. Regression Analysis of Head Capsule Widths
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rim, K.; Golec, J.R.; Duan, J.J. Host selection and potential non-target risk of Dastarcus helophoroides, a larval parasitoid of the Asian long-horned beetle, Anoplophora glabripennis. Biol. Control 2018, 123, 120–126. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, X.; Zhang, Y.; Zhang, Y. Research advances of Chinese major forest pests by integrated management based on biological control. Chin. J. Biol. Control 2018, 34, 163. [Google Scholar]
- Wang, Q. Cerambycid pests in agricultural and horticultural crops. In Cerambycidae of the World; CRC Press: Boca Raton, FL, USA, 2017; pp. 423–576. [Google Scholar]
- Mamiya, Y. Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus [Pinus densiflora, Pinus thunbergii, Pinus luchuensis, Japan]. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef]
- Robertson, L.; Cobacho Arcos, S.; Escuer, M.; Santiago Merino, R.; Esparrago, G.; Abelleira, A.; Navas, A. Incidence of the pinewood nematode Bursaphelenchus xylophlius Steiner & Buhrer, 1934 (Nickle, 1970) in Spain. Nematology 2011, 13, 755–757. [Google Scholar]
- Meng, J. Sustainability: A framework of typology based on efficiency and effectiveness. J. Macromark. 2015, 35, 84–98. [Google Scholar] [CrossRef]
- Siviter, H.; Muth, F. Do novel insecticides pose a threat to beneficial insects? Proc. R. Soc. B 2020, 287, 20201265. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, J.; du Plessis, H. Chemical control and insecticide resistance in Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2022, 115, 1761–1771. [Google Scholar] [CrossRef] [PubMed]
- Deveci, H.A.; Nur, G.; Deveci, A.; Kaya, I.; Kaya, M.M.; Kükürt, A.; Gelen, V.; Başer, Ö.F.; Karapehlivan, M. An Overview of the Biochemical and Histopathological Effects of Insecticides; IntechOpen: London, UK, 2021. [Google Scholar]
- Wang, G.; Xu, X.; Cheng, Q.; Hu, J.; Xu, X.; Zhang, Y.; Guo, S.; Ji, Y.; Zhou, C.; Gao, F.; et al. Preparation of sustainable release mesoporous silica nano-pesticide for control of Monochamus alternatus. Sustain. Mater. Technol. 2023, 35, e00538. [Google Scholar] [CrossRef]
- Saddam, B.; Idrees, M.A.; Kumar, P.; Mahamood, M. Biopesticides: Uses and importance in insect pest control: A review. Int. J. Trop. Insect Sci. 2024, 44, 1013–1020. [Google Scholar] [CrossRef]
- Tadahisa, U. Preliminary release experiments in laboratory and outdoor cages of Dastarcus helophoroides (Fairmaire) (Coleoptera: Bothrideridae) for biological control of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Bull. For. For. Prod. Res. Inst. 2003, 2, 255–262. [Google Scholar]
- Li, M.; Li, Y.; Lei, Q.; Yang, Z. Biocontrol of Asian long-horned beetle larva by releasing eggs of Dastarcus helophoroides (Coleoptera: Bothrideridae). Sci. Silvae Sin. 2009, 45, 78–82. [Google Scholar]
- Tang, Y.; Gao, S.; Zhang, Y.; Yang, Z.; Lu, J.; Zhan, M.; Wang, J. Key Environmental Factors Affecting Parasitism of Monochamus alternatus Hope by Dastarcus helophoroides Fairmaire. Chin. J. Biol. Control 2015, 31, 830. [Google Scholar]
- Castoreña, M.M.V.; Valencia, E.A.C. Determinación de estadios larvales de Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) para la construcción de un modelo de predicción. Folia Entomol. Mex. 2004, 43, 307–312. [Google Scholar]
- Gullan, P.; Cranston, P. The Insects: An Outline of Entomology. Nature 1994, 370, 261. [Google Scholar]
- Yamany, A.S.; Abdel-Gaber, R. Identification of fourth-instar larvae of Aedes albopictus (Skuse) (Diptera: Culicidae) employing scanning electron microscopic tool. Microsc. Res. Tech. 2024, 87, 933–947. [Google Scholar] [CrossRef]
- Lambiase, S.; Corotti, S.; Sacchi, R. Morphometric analysis for determination of larval instars in Dermestes frischii Kugelann and Dermestes undulatus Brahm (Coleoptera: Dermestidae). J. Forensic Sci. 2024, 69, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
- Merville, A.; Vallier, A.; Venner, S.; Siberchicot, A.; Fouchet, D.; Heddi, A.; Bel-Venner, M.-C. Determining the instar of a weevil larva (Coleoptera: Curculionidae) using a parsimonious method. Eur. J. Entomol. 2014, 111, 567–573. [Google Scholar] [CrossRef]
- Panzavolta, T. Instar determination for Pissodes castaneus (Coleoptera: Curculionidae) using head capsule widths and lengths. Environ. Entomol. 2014, 36, 1054–1058. [Google Scholar] [CrossRef]
- Cazado, L.E.; Van Nieuwenhove, G.A.; O’brien, C.; Gastaminza, G.A.; Murúa, M.G. Determination of number of instars of Rhyssomatus subtilis (Coleoptera: Curculionidae) based on head capsule widths. Fla. Entomol. 2014, 97, 639–643. [Google Scholar] [CrossRef]
- Dyar, H.G. The number of molts of lepidopterous larvae. Psyche 1890, 5, 420–422. [Google Scholar] [CrossRef]
- Gaines, J.; Campbell, F. Dyar’s rule as related to the number of instars of the corn ear worm, Heliothis obsoleta (Fab.), collected in the field. Ann. Entomol. Soc. Am. 1935, 28, 445–461. [Google Scholar] [CrossRef]
- Chen, Y.; Seybold, S. Application of a frequency distribution method for determining instars of the beet armyworm (Lepidoptera: Noctuidae) from widths of cast head capsules. J. Econ. Entomol. 2013, 106, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Su, J.M.; Hao, C.F.; Li, M.L. Female reproductive system of Dastarcus helophoroides (Fairmaire) (Coleoptera: Bothrideridae). J. Northwest AF Univ. 2016, 44, 100–108. [Google Scholar]
- Tang, H.; Yang, Z.; Zhang, Y.; Li, G. Technical researches on distinguishing female and male alive adults of the main parasite of longhorn beetles, Dastarcus helophoroides (Coleoptera: Bothrideridae) without injuring. Acta Zoot. Sin 2007, 32, 649–654. [Google Scholar]
- Han, X.; Duan, Y.L.; Li, X.F.; Wang, Z.Y.; Zhang, Y.N.; Qiao, L.Q. Sensilla Ultrastructure of Antennae and Mouthparts of the First Instar Larvae of Dastarcus helophoroides (Coleoptera: Bothrideridae). For. Sci. Res. 2021, 34, 180–184. [Google Scholar]
- Pei, P. Dastarcus helophoroides P450 Participates in the Molecular Mechanism of Cypermethrin Stress Response. Master’s Thesis, Northwest A&F University, Yangling, China, 2021. [Google Scholar]
- Wang, X.; Ni, X.; Duan, C.; Li, R.; Jiang, X.; Xu, M.; Yu, R.J. The effect of ultrasound treatment on the structural and functional properties of Tenebrio molitor Myofibrillar protein. Foods 2024, 13, 2817. [Google Scholar] [CrossRef]
- Sun, C.W. Flying Ability and It’s Influencing Factors of Dastarcus helophoroides Adults. Master’s Thesis, Shandong Agricultural University, Jinan, China, 2019. [Google Scholar]
- Zhang, X. Effects of Dastarcus helophoroides Larval Density on the Degeneration of Subsitute Host Nutrition and Metabolism. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2018. [Google Scholar]
- Delbac, L.; Lecharpentier, P.; Thiery, D. Larval instars determination for the European Grapevine Moth (Lepidoptera: Tortricidae) based on the frequency distribution of head-capsule widths. Crop Prot. 2010, 29, 623–630. [Google Scholar] [CrossRef]
- Sukovata, L. A comparison of three approaches for larval instar separation in insects—A case study of Dendrolimus pini. Insects 2019, 10, 384. [Google Scholar] [CrossRef]
- Li, N.; Wu, L.; Geng, Y.; Wei, D.; Chen, M. Determination of larval instars of Semanotus bifasciatus (Coleoptera: Cerambycidae) based on frequency distributions of morphological variables. J. Entomol. Sci. 2020, 55, 405–415. [Google Scholar] [CrossRef]
- Ramasubramanian, T.; Rajan, T.S.; Sudhanan, E.M. Instar determination for sugarcane internode borer Chilo sacchariphagus indicus (Kapur) (Lepidoptera: Crambidae). J. Asia-Pac. Entomol. 2021, 24, 461–469. [Google Scholar] [CrossRef]
- Brockelsby, W.D.; Miskelly, C.M.; Glare, T.R.; Minor, M.A. The number of larval instars in the flax weevil (Anagotus fairburni) (Coleoptera: Curculionidae). N. Z. J. Zool. 2023, 52, 1–11. [Google Scholar] [CrossRef]
- McClellan, Q.; Logan, J. Instar determination for the gypsy moth (Lepidoptera: Lymantriidae) based on the frequency distribution of head capsule widths. Environ. Entomol. 1994, 23, 248–253. [Google Scholar] [CrossRef]
- Logan, J.; Bentz, B.; Vandygriff, J.; Turner, D. General program for determining instar distributions from headcapsule widths: Example analysis of mountain pine beetle (Coleoptera: Scolytide) data. Environ. Entomol. 1998, 27, 555–563. [Google Scholar] [CrossRef]
- Brooks, W.K. Report on the Stomatopoda collected by HMS Challenger during the years 1873–76. In Report on the Scientific Results of the Voyage of HMS; Challenger Reports; 1886; Volume 16, pp. 1–16. [Google Scholar] [CrossRef]
- Crosby, T. Dyar’s rule predated by Brooks’ rule. N. Z. Entomol. 1973, 5, 175–176. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, C.; Li, S.; Zhu, H.; Fan, B.; Wang, Y.; Su, P.; Han, Y.; Hao, D. Biological traits and life history of Pagiophloeus tsushimanus (Coleoptera: Curculionidae), a weevil pest on camphor trees in China. J. For. Res. 2021, 32, 1979–1988. [Google Scholar] [CrossRef]
- Yu, Y.; Cen, G.; Wei, D.; Zeng, X.; Zeng, T. Division of larval instars of Dorysthenes granulosus based on Crosby growth rule. J. South. Agric. 2012, 43, 1485–1489. [Google Scholar]
- Castañeda-Vildózola, Á.; González-Hernández, H.; Equihua-Martínez, A.; Valdez-Carrasco, J.; Peña, J.E.; Cazado, L.E.; Franco-Mora, O. Head capsule width is useful for determining larval instar in Heilipus lauri (Coleoptera: Curculionidae). Fla. Entomol. 2016, 99, 822–825. [Google Scholar] [CrossRef]
- Luo, W.; Ji, Y.-C.; Wen, J.-B. Application of a frequency distribution method for determining instars of Eucryptorrhynchus brandti (Coleoptera: Curculionidae) from several morphological variables. Biocontrol Sci. Technol. 2016, 26, 1329–1336. [Google Scholar] [CrossRef]
- Velásquez, Y.; Viloria, Á.L. Instar determination of the neotropical beetle Oxelytrum discicolle (Coleoptera: Silphidae). J. Med. Entomol. 2010, 47, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Fauteux, A.; Gonzalez, N.; Soares, A.O.; Lucas, É. Morphological determination of the larval instars of Eupeodes americanus (Diptera: Syrphidae). Phytoprotection 2022, 102, 30–34. [Google Scholar] [CrossRef]
- Shi, Y.; Wang, L.; Dou, W.; Jiang, H.-B.; Wei, D.-D.; Wei, D.; Niu, J.-Z.; Wang, J.-J. Determination of instars of Bactrocera dorsalis (Diptera: Tephritidae). Fla. Entomol. 2017, 100, 270–275. [Google Scholar] [CrossRef]
- Peterson, M.K.; Appel, A.G.; Hu, X.P. Instar determination of Blattella asahinai (Blattodea: Ectobiidae) from digital measurements of the pronotum using Gaussian mixture modeling and the number of cercal annuli. J. Insect Sci. 2019, 19, 5. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.; Haeussler, G. Some observations on the number of larval instars of the oriental peach moth, Laspeyresia molesta Busck. J. Econ. Entomol. 1928, 21, 843–852. [Google Scholar] [CrossRef]
- Chen, Y.; Dallara, P.L.; Nelson, L.J.; Coleman, T.W.; Hishinuma, S.M.; Carrillo, D.; Seybold, S.J. Comparative morphometric and chemical analyses of phenotypes of two invasive ambrosia beetles (Euwallacea spp.) in the United States. Insect Sci. 2017, 24, 647–662. [Google Scholar] [CrossRef]
- Preto, C.R.; Bellamy, D.E.; Walse, S.S.; Zalom, F.G. Predicting larval stage distribution of Lobesia botrana (Lepidoptera: Tortricidae) at three constant temperatures. J. Econ. Entomol. 2019, 112, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Calvo, D.; Molina, J.M. Head capsule width and instar determination for larvae of Streblote panda (Lepidoptera: Lasiocampidae). Ann. Entomol. Soc. Am. 2008, 101, 881–886. [Google Scholar] [CrossRef]
- Hansen, J.D.; Owens, J.C.; Huddleston, E.W. Relation of head capsule width to instar development in larvae of the range caterpillar, Hemileuca oliviae Cockerell (Lepidoptera: Saturniidae). J. Kans. Entomol. Soc. 1981, 54, 1–7. [Google Scholar]
- Cave, G.; Smith, C. Number of instars of the rice water weevil, Lissorhoptrus oryzophilus (Coleoptera: Curculionidae). Ann. Entomol. Soc. Am. 1983, 76, 293–294. [Google Scholar] [CrossRef]
- Zawadneak, M.A.; Goncalves, R.B.; Poltronieri, A.S.; Santos, B.; Bischoff, A.M.; Borba, A.M.; Pimentel, I.C. Biological parameters of Duponchelia fovealis (Lepidoptera: Crambidae) reared in the laboratory on two diets. Eur. J. Entomol. 2017, 114, 291–294. [Google Scholar] [CrossRef]
- Gousul, N.; Buhroo, A.A. Bionomics of slender burnished brass (Thysanoplusia orichalcea [Fabricius, 1775], Lepidoptera: Noctuidae) on potato (Solanum tuberosum L.) in Kashmir. Acta Agric. Slov. 2021, 117, 1–8. [Google Scholar]
- Cen, G.; Yu, Y.; Zeng, X.; Long, X.; Wei, D.; Gao, X.; Zeng, T. An adaptive kernel smoothing method for classifying Austrosimulium tillyardianum (Diptera: Simuliidae) larval instars. J. Insect Sci. 2015, 15, 159. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Wang, J.; Wang, W.; Zheng, Y. Larval Instars and Adult Flight Period of Monochamus saltuarius (Coleoptera: Cerambycidae). Forests 2022, 13, 910. [Google Scholar] [CrossRef]
- Yang, R.; Qiu, P.; Gu, Y.; Ni, M.; Xue, Z.; Han, J.; Jiang, Y.; Jin, Y.; Wang, Y.; Zhou, X. Biology of Rhynchaenus maculosus provides insights and implications for integrated management of this emerging pest. Sci. Rep. 2022, 12, 14650. [Google Scholar] [CrossRef]
- Esperk, T.; Tammaru, T.; Nylin, S. Intraspecific variability in number of larval instars in insects. J. Econ. Entomol. 2007, 100, 627–645. [Google Scholar] [CrossRef]
- Langor, D.W.; Spence, J.R.; Pohl, G.R. Host effects on fertility and reproductive success of Dendroctonus ponderosae Hopkins (Coleoptera: Scolytidae). Evolution 1990, 44, 609–618. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Kay, S.; Rojas, M.G.; Shapiro-Ilan, D.I.; Tedders, W.L. Morphometric analysis of instar variation in Tenebrio molitor (Coleoptera: Tenebrionidae). Ann. Entomol. Soc. Am. 2015, 108, 146–159. [Google Scholar] [CrossRef]
- Guo, W.; Gao, P.; Wang, F.; Hu, Z. Occurrence and control techniques of Anoplophora glabripennis and Anoplophora chinensis. Mod. Gard. 2016, 8, 53. [Google Scholar]
- Yang, C.; Wei, Y.; Li, C.; Huang, S.; Li, L.; Ma, Z. Division of larval instars of the coconut black-headed caterpillar, Opisina arenosella. Plant Prot. 2015, 41, 70–74. [Google Scholar]
- Xu, J.H.; Huang, X.F.; Xu, H.C.; Cheng, J.M.; Jiang, Y.; Fang, W.J. Rearing and biological properties of Monochamus alternatus. J. Zhejiang For. Sci. 2009, 29, 86–88. [Google Scholar]
- Huang, D.; Wang, R.J.; Tang, P.; Li, G.Z.; Peng, J.Y.; Wang, S.F.; Zhu, L.X. Study on the age division of Monochamus alternatus in Panzhihua City and the regularity of each age. Sichuan Agric. Sci. Tech. 2018, 373, 31–33. [Google Scholar]
- Bleiker, K.; Régnière, J. Determining the instar of mountain pine beetle (Coleoptera: Curculionidae) larvae by the width of their head capsules. Can. Entomol. 2015, 147, 635–640. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Instar | n | Head Capsule Width | CV (%) | Brooks–Dyar’s Ratio | Crosby’s Growth Ratio | |
|---|---|---|---|---|---|---|
| Mean ± SD (µm) | Range | |||||
| 1 | 63 | 95.52 ± 7.62 | 71.41–112.13 | 4.60 | ||
| 2 | 70 | 175.64 ± 23.69 | 142.83–255.15 | 7.78 | 1.83 | |
| 3 | 80 | 318.01 ± 31.57 | 270.18–406 | 5.73 | 1.81 | −0.01 |
| 4 | 80 | 560.06 ± 43.46 | 466.6–649.93 | 4.48 | 1.76 | −0.02 |
| Mean growth rate | 1.80 |
| Instar | n | Head Capsule Width | CV (%) | Brooks–Dyar’s Ratio | |
|---|---|---|---|---|---|
| Mean ± SD (µm) | Range | ||||
| 1 | 63 | 95.68 ± 9.21 | ˂116.14 | 4.60 | |
| 2 | 66 | 175.52 ± 13.64 | 116.14–235.42 | 5.37 | 1.83 |
| 3 | 84 | 316.80 ± 28.17 | 235.42–429 | 6.30 | 1.80 |
| 4 | 80 | 560.83 ± 38.78 | >429 | 4.48 | 1.77 |
| Mean growth rate | 1.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaheen, T.; Guo, J.; Wang, Y.; Zhou, J.; Tang, G.; Zhang, Z. Determination of Larval Instars of Dastarcus helophoroides (Coleoptera: Bothrideridae) Using Head Capsule Width Frequency Distribution. Insects 2024, 15, 1013. https://doi.org/10.3390/insects15121013
Shaheen T, Guo J, Wang Y, Zhou J, Tang G, Zhang Z. Determination of Larval Instars of Dastarcus helophoroides (Coleoptera: Bothrideridae) Using Head Capsule Width Frequency Distribution. Insects. 2024; 15(12):1013. https://doi.org/10.3390/insects15121013
Chicago/Turabian StyleShaheen, Tayyab, Jiali Guo, Yun Wang, Jiaojiao Zhou, Guanghui Tang, and Zhengqing Zhang. 2024. "Determination of Larval Instars of Dastarcus helophoroides (Coleoptera: Bothrideridae) Using Head Capsule Width Frequency Distribution" Insects 15, no. 12: 1013. https://doi.org/10.3390/insects15121013
APA StyleShaheen, T., Guo, J., Wang, Y., Zhou, J., Tang, G., & Zhang, Z. (2024). Determination of Larval Instars of Dastarcus helophoroides (Coleoptera: Bothrideridae) Using Head Capsule Width Frequency Distribution. Insects, 15(12), 1013. https://doi.org/10.3390/insects15121013