
Predation Risk Effects of Lady Beetle Menochilus sexmaculatus (Fabricius) on the Melon Aphid, Aphis gossypii Glover

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Aphids for Laboratory Experiments
2.2. Menochilus sexmaculatus for Laboratory Experiments
2.3. Behavioral Responses of Aphis gossypii to Menochilus sexmaculatus Risk
2.4. The Effect of Menochilus sexmaculatus Risk on Aphid Biology
2.5. Oxidative Stress Level and Oxidative Damage of Aphids under Menochilus sexmaculatus Risk
2.6. Identification of Compounds of Menochilus sexmaculatus
2.7. Behavioral Responses of Aphis gossypii to Compounds of Menochilus sexmaculatus
2.8. Statistics
3. Results
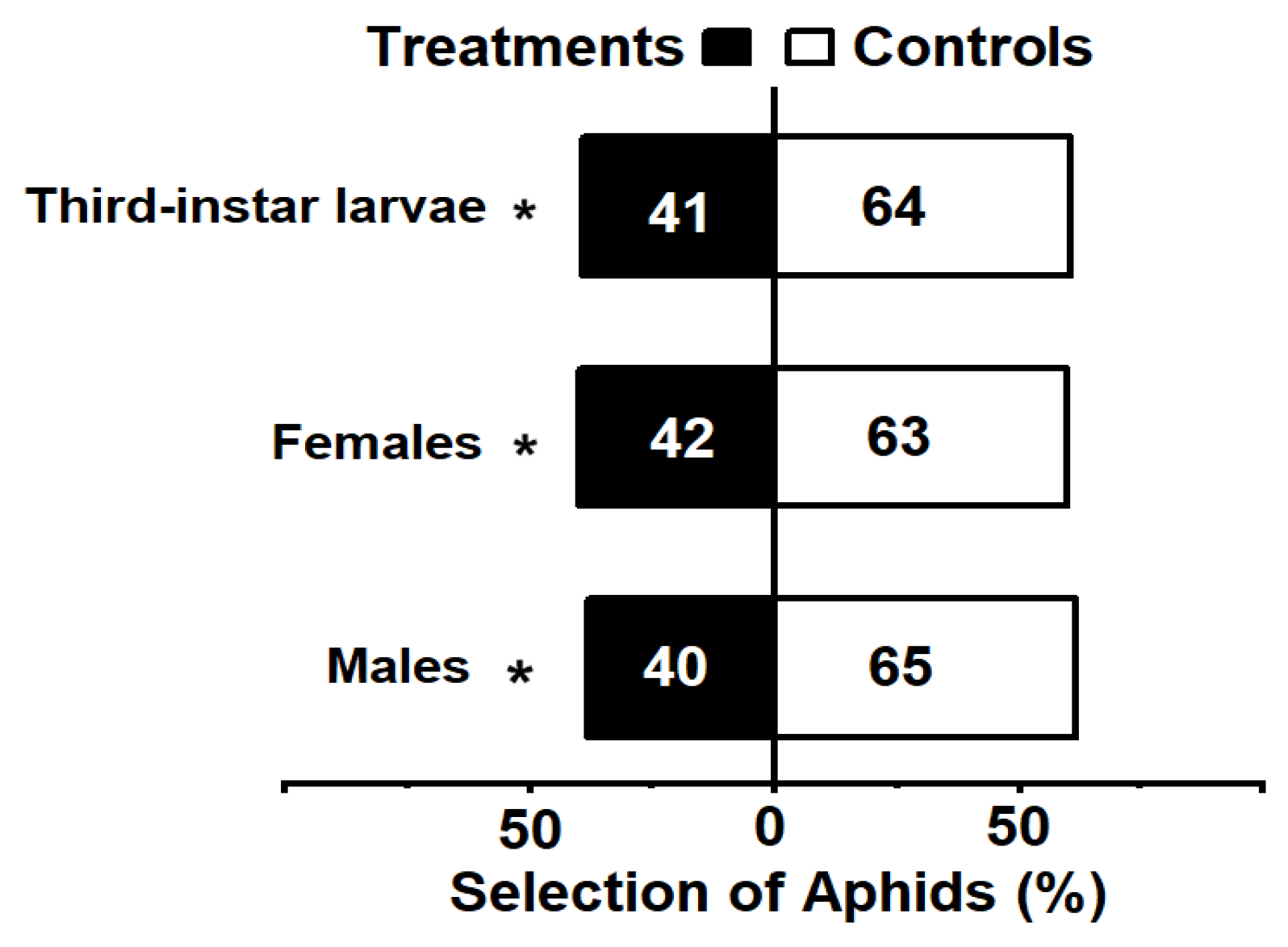
3.1. The Behavioral Response of Aphis gossypii to Menochilus sexmaculatus
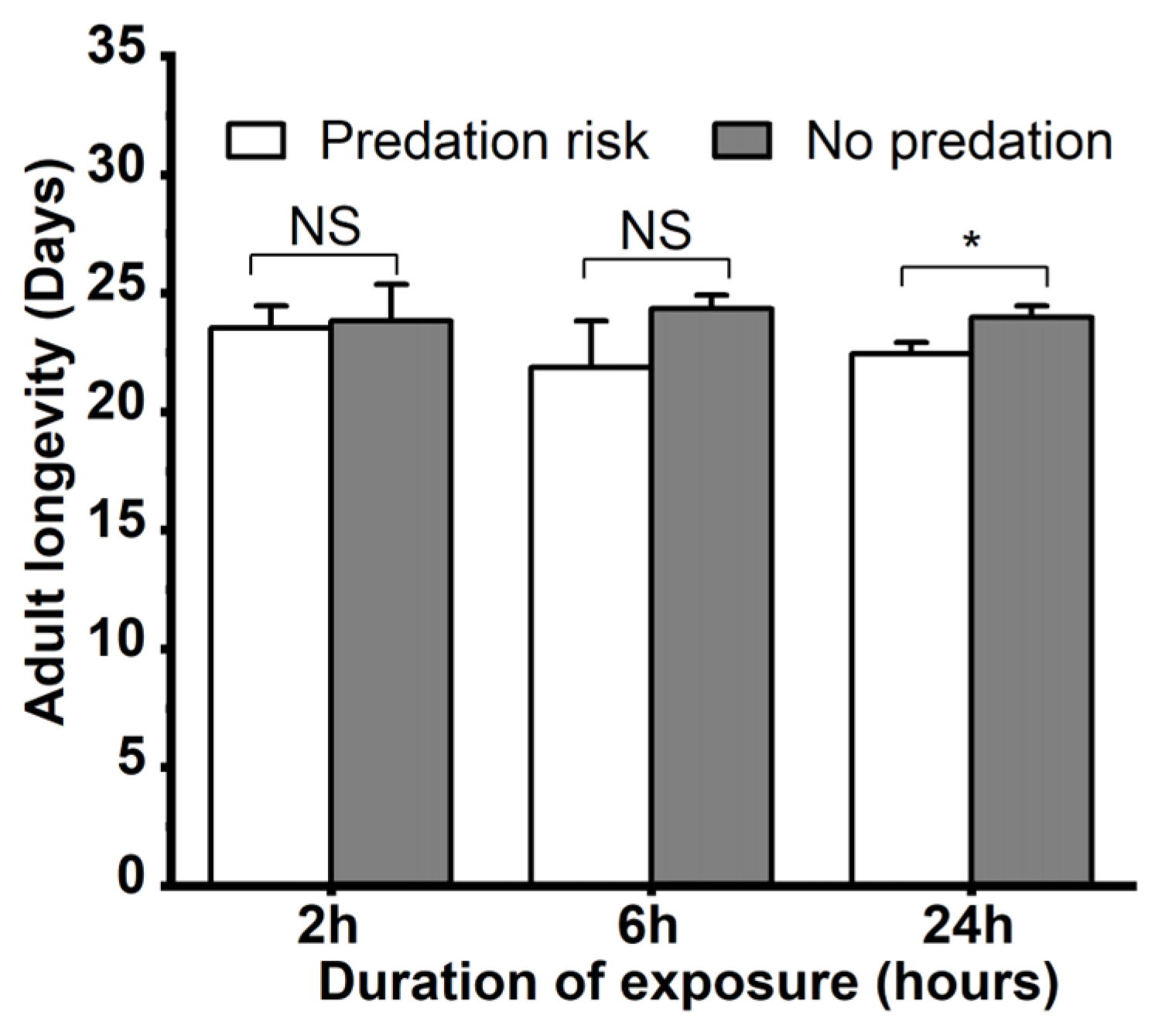
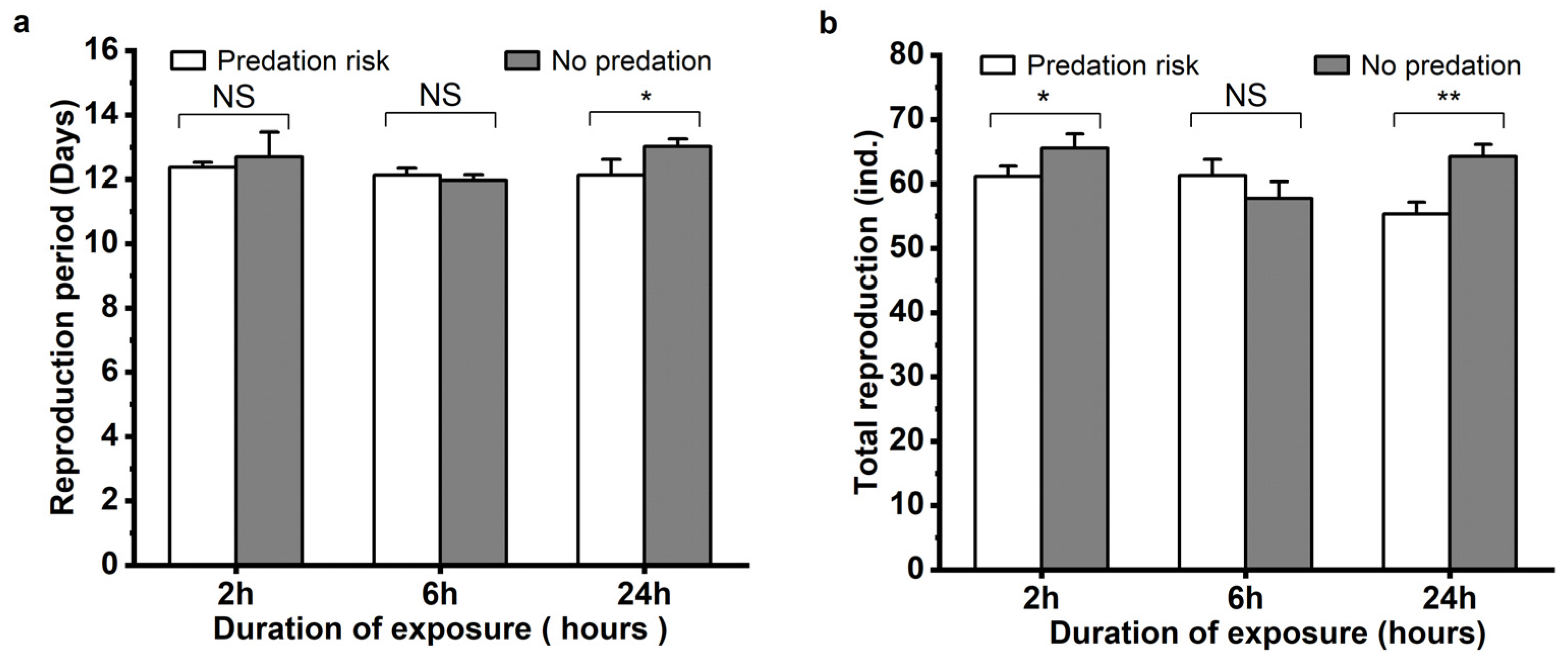
3.2. The Influence of Menochilus sexmaculatus Risk on Growth and Fecundity of Aphis gossypii
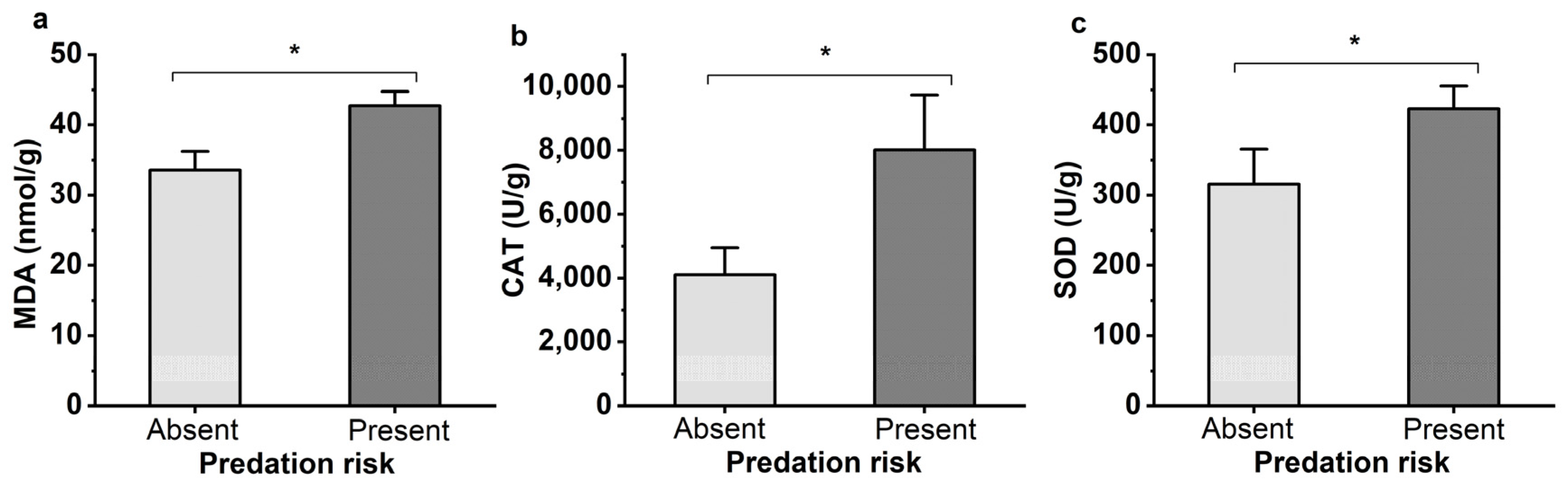
3.3. Oxidative Stress Level and Oxidative Damage of Aphis gossypii under Predation Risk
3.4. GC-MS Analysis of the Compounds of Menochilus sexmaculatus
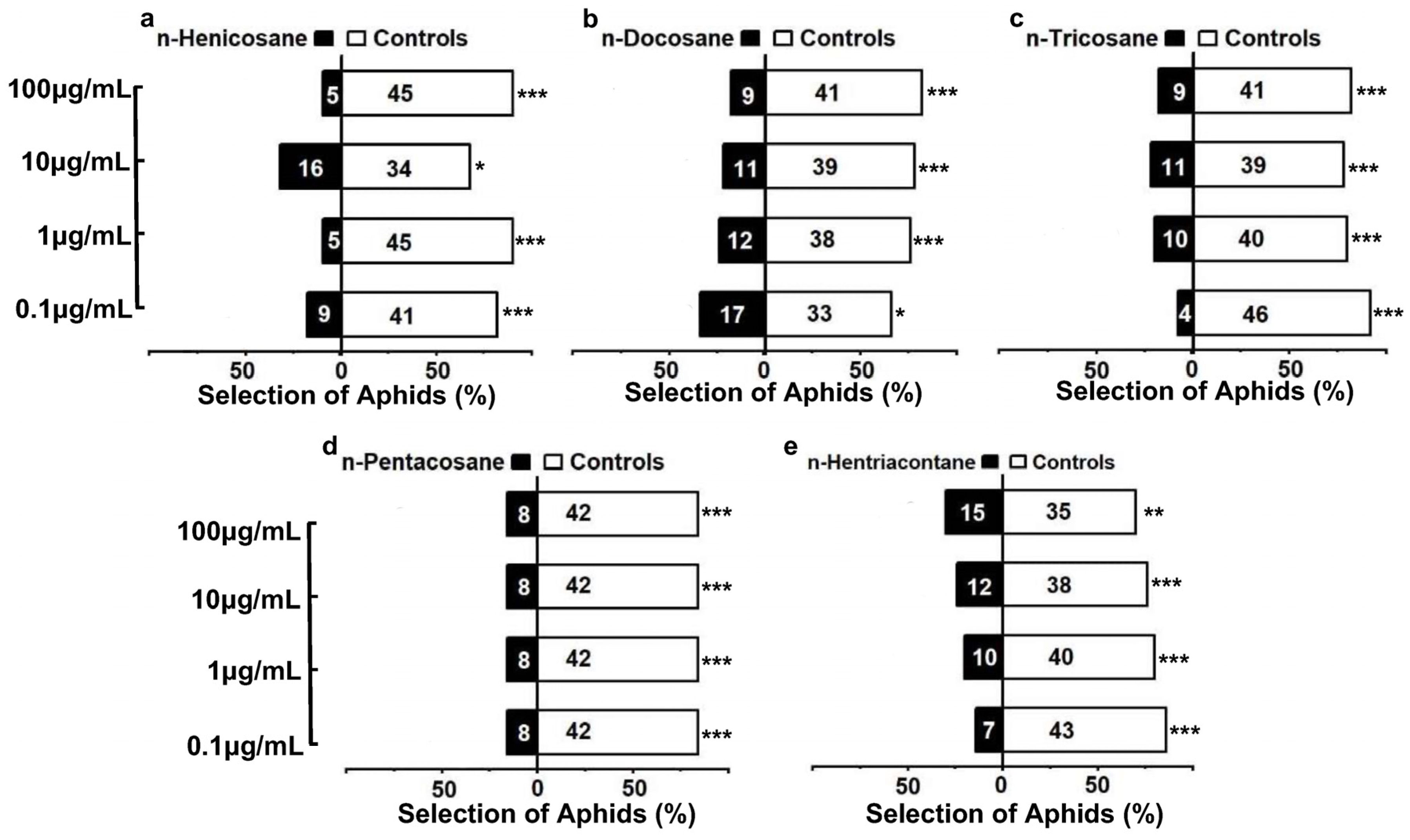
3.5. Behavioral Responses of Aphis gossypii to Alkane Compounds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hart, A.; Brown, C.D.; Lewis, K.A.; Tzilivakis, J. p-EMA (II): Evaluating ecological risks of pesticides for a farm-level risk assessment system. Agronomie 2003, 23, 75–84. [Google Scholar] [CrossRef][Green Version]
- Pimentel, D.; Burgess, M. Environmental and economic costs of the application of pesticides primarily in the United States. Environ. Dev. Sustain. 2005, 7, 229–252. [Google Scholar] [CrossRef]
- Werner, E.E.; Peacor, S.D. A review of trait-mediated indirect interactions in ecological communities. Ecology 2003, 84, 1083–1100. [Google Scholar] [CrossRef]
- Kunert, G.; Weisser, W.W. The interplay between density- and trait-mediated effects in predator-prey interactions: A case study in aphid wing polymorphism. Oecologia 2003, 135, 304–312. [Google Scholar] [CrossRef]
- Hermann, S.L.; Landis, D.A. Scaling up our understanding of non-consumptive effects in insect systems. Curr. Opin. Insect Sci. 2017, 20, 54–60. [Google Scholar] [CrossRef]
- Hermann, S.L.; Thaler, J.S. Prey perception of predation risk: Volatile chemical cues mediate non-consumptive effects of a predator on a herbivorous insect. Oecologia 2014, 176, 669–676. [Google Scholar] [CrossRef]
- Wen, J.; Ueno, T. Predator cue-induced plasticity of morphology and behavior in planthoppers facilitate the survival from predation. Sci. Rep. 2021, 11, 16760. [Google Scholar] [CrossRef]
- Silberbush, A.; Markman, S.; Lewinsohn, E.; Bar, E.; Cohen, J.E.; Blaustein, L. Predator-released hydrocarbons repel oviposition by a mosquito. Eco. Lett. 2010, 13, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Mollo, E.; Garson, M.J.; Polese, G.; Amodeo, P.; Ghiselin, M.T. Taste and smell in aquatic and terrestrial environments. Nat. Prod. Rep. 2017, 34, 496–513. [Google Scholar] [CrossRef] [PubMed]
- Hermann, S.L.; Bird, S.A.; Ellis, D.R.; Landis, D.A. Predation risk differentially affects aphid morphotypes: Impacts on prey behavior, fecundity and transgenerational dispersal morphology. Oecologia 2021, 197, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Ninkovic, V.; Feng, Y.; Olsson, U.; Pettersson, J. Ladybird footprints induce aphid avoidance behavior. Biol. Control. 2013, 65, 63–71. [Google Scholar] [CrossRef]
- Pervez, A.; Omkar. Functional responses of coccinellid predators: An illustration of a logistic approach. J. Insect Sci. 2005, 5, 5. [Google Scholar] [CrossRef]
- Patel, P.; Kumar, B.; Upadhyay, R.; Bhusal, D.; Kumar, D. Semiochemical tracks of predaceous Coccinellidae (Coleoptera) modulate feeding attributes and assimilation of nutrients in Zygogramma bicolorata (Coleoptera: Chrysomelidae). Can. Entomol. 2020, 152, 330–341. [Google Scholar] [CrossRef]
- Jeffrey, C. A review of the Cucurbitaceae. Bot. J. Linn. Soc. 1980, 81, 233–247. [Google Scholar] [CrossRef]
- Mliki, A.; Staub, J.E.; Sun, Z.Y.; Ghorbel, A. Genetic diversity in African cucumber (Cucumis sativus L.) provides potential for germplasm enhancement. Genet. Resour. Crop Ev. 2003, 50, 461–468. [Google Scholar] [CrossRef]
- Liang, D.; Liu, M.; Hu, Q.J.; He, M.; Qi, X.H.; Xu, Q.; Zhou, F.C.; Chen, X.H. Identification of differentially expressed genes related to aphid resistance in cucumber (Cucumis sativus L.). Sci. Rep. 2015, 5, 9645. [Google Scholar] [CrossRef]
- Satar, S.; Kersting, U.; Uygun, N. Effect of temperature on development and fecundity of Aphis gossypii Glover (Homoptera: Aphididae) on cucumber. J. Pest. Sci. 2005, 78, 133–137. [Google Scholar] [CrossRef]
- Boivin, G.; Hance, T.; Brodeur, J. Aphid parasitoids in biological control. Can. J. Plant Sci. 2012, 92, 1–12. [Google Scholar] [CrossRef]
- Dong, W.; Yang, H.; Wang, C.; Li, H.; Shang, J.; Chen, Z.; Zhang, H.; Liang, P.; Gao, X.; Shi, X. Cross-resistance and fitness costs of the cis-nitromethylene neonicotinoid cycloxaprid resistance in melon aphid, Aphis gossypii (Hemiptera: Aphididae). J. Econ. Entomol. 2022, 115, 1668–1675. [Google Scholar] [CrossRef] [PubMed]
- Nelson, E.H. Predator avoidance behavior in the pea aphid: Costs, frequency, and population consequences. Oecologia 2007, 151, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.L. Nonlethal effects in the ecology of predator-prey interactions: What are the ecological effects of anti-predator decision-making? Bioscience 1998, 48, 25–34. [Google Scholar] [CrossRef]
- Zhang, R.B.; Wang, B.; Grossi, G.; Falabella, P.; Liu, Y.; Yan, S.C.; Lu, J.; Xi, J.H.; Wang, G.R. Molecular basis of alarm pheromone detection in aphids. Curr. Biol. 2017, 27, 55–61. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kitchell, J.F.; Hodgson, J.R.; Cochran, P.A.; Elser, M.M.; Lodge, D.M.; Kretchmer, X.H. Regulation of lake primary productivity by food web structure. Ecology 1987, 68, 1863–1876. [Google Scholar] [CrossRef]
- Peacor, S.D.; Peckarsky, B.L.; Trussell, G.C.; Vonesh, J.R. Costs of predator-induced phenotypic plasticity: A graphical model for predicting the contribution of nonconsumptive and consumptive effects of predators on prey. Oecologia 2013, 171, 1–10. [Google Scholar] [CrossRef]
- Sheriff, M.J.; Thaler, J.S. Ecophysiological effects of predation risk; an integration across disciplines. Oecologia 2014, 176, 607–611. [Google Scholar] [CrossRef]
- Apfelbach, R.; Blanchard, C.D.; Blanchard, R.J.; Hayes, R.A.; McGregor, T.S. The effects of predator odors in mammalian prey species: A review of field and laboratory studies. Neurosci. Biobehav. Rev. 2005, 29, 1123–1144. [Google Scholar] [CrossRef]
- Amo, L.; López, P.; Martín, J. Can wall lizards combine chemical and visual cues to discriminate predatory from non-predatory snakes inside refuges? Ethology 2006, 112, 478–484. [Google Scholar] [CrossRef]
- Zhang, H. Recognition and responding of Acyrthosiphon pisum to odor and traces of harmonia axyridis. Master’s Dissertation, Northwest A&F University, Yangling, China, 2016. [Google Scholar]
- Thaler, J.S.; McArt, S.H.; Kaplan, I. Compensatory mechanisms for ameliorating the fundamental trade-off between predator avoidance and foraging. Proc. Natl. Acad. Sci. USA 2012, 109, 12075–12080. [Google Scholar] [CrossRef]
- Rypstra, A.L.; Buddle, C.M. Spider silk reduces insect herbivory. Biol. Lett. 2010, 9, 20120948. [Google Scholar] [CrossRef] [PubMed]
- Rendon, D.; Whitehouse, E.A.M.; Taylor, W.P. Consumptive and non-consumptive effects of wolf spiders on cotton bollworms. Entomol. Exp. Appl. 2016, 158, 170–183. [Google Scholar] [CrossRef]
- Sendoya, F.; Freitas, V.L.; Oliveira, P.S. Egg-laying butterflies distinguish predaceous ants by sight. Am. Nat. 2009, 174, 134–140. [Google Scholar] [CrossRef]
- Lee, D.H.; Nyrop, J.P.; Sanderson, J.P. Non-consumptive effects of the predatory beetle Delphastus catalinae (Coleoptera: Coccinellidae) on habitat use patterns of adult whitefly Bemisia argentifolii (Hemiptera: Aleyrodidae). Appl. Entomol. Zool. 2014, 49, 599–606. [Google Scholar] [CrossRef]
- Li, J.J.; Wang, Z.W.; Tan, K.; Qu, Y.F.; Nieh, J.C. Giant Asian honeybees use olfactory eavesdropping to detect and avoid ant predators. Anim. Behav. 2014, 97, 69–76. [Google Scholar] [CrossRef]
- Hermann, S.L.; Thaler, J.S. The effect of predator presence on the behavioral sequence from host selection to reproduction in an invulnerable stage of insect prey. Oecologia 2018, 188, 945–952. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J. Free radicals in biology and medicine (5th edn). Oxf. Academic. 2015, 22, 199–283. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Michaud, D.; Cloutier, C. A proteomic analysis of the aphid macrosiphum euphorbiae under heat and radiation stress. Insect Biochem. Mol. Biol. 2009, 39, 20–30. [Google Scholar] [CrossRef]
- Li, J.; Zhao, H.Y.; Zhao, X.D. Study on effect of ecological characteristics and enzyme activity of aphid radiated by different intensities of ultraviolet. J. Northwest A F Univ. (Nat. Sci. Ed.) 2005, 4, 61–64. [Google Scholar] [CrossRef]
- Janssens, L.; Stoks, R. Predation risk causes oxidative damage in prey. Biol. Lett. 2013, 9, 20130350. [Google Scholar] [CrossRef]
- Slos, S.; Meester, L.D.; Stoks, R. Food level and sex shape predator-induced physiological stress: Immune defence and antioxidant defence. Oecologia 2009, 161, 461–467. [Google Scholar] [CrossRef]
- Venkanna, Y.; Suroshe, S.; Dahuja, A. Non-consumptive effects of the zigzag ladybird beetle, Cheilomenes sexmaculata (Fab.) on its prey, the cotton aphid, Aphis gossypii Glover. Biocontrol Sci. Technol. 2021, 31, 1204–1219. [Google Scholar] [CrossRef]
- McGraw, K.J.; Cohen, A.A.; Constantini, D.; Hõrak, P. The ecological significance of antioxidants and oxidative stress: A marriage between mechanistic and functional perspectives. Funct. Ecol. 2010, 24, 947–949. [Google Scholar] [CrossRef]
- Monaghan, P.; Metcalfe, B.N.; Torres, R. Oxidative stress as a mediator of life history trade-offs: Mechanisms, measurements and interpretation. Ecol. Lett. 2008, 12, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Mappes, J.; Mappes, T.; Lappalainen, T. Unequal maternal investment in offspring quality in relation to predation risk. Evol. Ecol. 1997, 11, 237–243. [Google Scholar] [CrossRef]
- Sitvarin, M.I.; Breen, K.; Rypstra, A.L. Predator cues have contrasting effects on lifespan of Pardosa milvina (Araneae: Lycosidae). J. Arachnol. 2015, 43, 107–110. [Google Scholar] [CrossRef]
- Stoks, R. Food stress and predator-induced stress shape developmental performance in a damselfly. Oecologia 2001, 127, 222–229. [Google Scholar] [CrossRef]
- Elliott, K.H.; Betini, G.S.; Dworkin, I.; Norris, D.R. Experimental evidence for within- and cross-seasonal effects of fear on survival and reproduction. J. Anim. Ecol. 2016, 85, 507–515. [Google Scholar] [CrossRef]
- Wang, B. The influence of non-consumptive effects of Harmonia axyridis on fitness of Acythosiphon pisum. Master’s Dissertation, Northwest A&F University, Yangling, China, 2020. [Google Scholar]
- Li, J.; Long, D.B.; Xiao, T.G.; Ou, Y.F. The impacts of stress from Lysiphlebia Japonica Ashmead on the development and fecundity of the three successive generations of aphids. Chin. J. Appl. Entomol. 2013, 50, 951–958. [Google Scholar] [CrossRef]
- Nakashima, Y.; Birkett, M.A.; Pye, B.J.; Pickett, J.A. The role of semiochemicals in the avoidance of the seven-spot ladybird, Coccinella septempunctata, by the aphid parasitoid, Aphidius ervi. J. Chem. Ecol. 2004, 30, 1103–1116. [Google Scholar] [CrossRef]
- Kosaki, A.; Yamaoka, R. Chemical composition of footprints and cuticular lipids of three species of lady beetles. J. Appl. Entomol. 1996, 40, 47–53. [Google Scholar] [CrossRef]
- Abassi, S.A.; Birkett, M.A.; Pettersson, J.; Pickett, J.A. Ladybird beetle odour identified and found to be responsible for attraction between adults. Cell Mol. Life Sci. 1998, 54, 876–879. [Google Scholar] [CrossRef]
- Bernays, E.A. Insect defenses: Adaptive mechanisms and strategies of prey and predators. Entomol. Am. 1991, 37, 63–91. [Google Scholar] [CrossRef]
- Telang, A.; Li, Y.; Noriega, F.G.; Brown, M.R. Effects of larval nutrition on the endocrinology of mosquito egg development. J. Exp. Biol. 2006, 209, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.R. Responses of aphids to semiochemicals released from ladybirds. Master’s Dissertation, Swedish University of Agricultural Sciences Department of Ecology, Uppsala, Swedish, 2012. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, X.; Cui, X.; Tang, J.; Zhu, J.; Li, J. Predation Risk Effects of Lady Beetle Menochilus sexmaculatus (Fabricius) on the Melon Aphid, Aphis gossypii Glover. Insects 2024, 15, 13. https://doi.org/10.3390/insects15010013
Lin X, Cui X, Tang J, Zhu J, Li J. Predation Risk Effects of Lady Beetle Menochilus sexmaculatus (Fabricius) on the Melon Aphid, Aphis gossypii Glover. Insects. 2024; 15(1):13. https://doi.org/10.3390/insects15010013
Chicago/Turabian StyleLin, Xingming, Xiangxin Cui, Jihong Tang, Jiawei Zhu, and Jinhua Li. 2024. "Predation Risk Effects of Lady Beetle Menochilus sexmaculatus (Fabricius) on the Melon Aphid, Aphis gossypii Glover" Insects 15, no. 1: 13. https://doi.org/10.3390/insects15010013
APA StyleLin, X., Cui, X., Tang, J., Zhu, J., & Li, J. (2024). Predation Risk Effects of Lady Beetle Menochilus sexmaculatus (Fabricius) on the Melon Aphid, Aphis gossypii Glover. Insects, 15(1), 13. https://doi.org/10.3390/insects15010013