
iORandLigandDB: A Website for Three-Dimensional Structure Prediction of Insect Odorant Receptors and Docking with Odorants


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Data Processing
2.3. Prediction of Secondary Structures and Three-Dimensional Structures
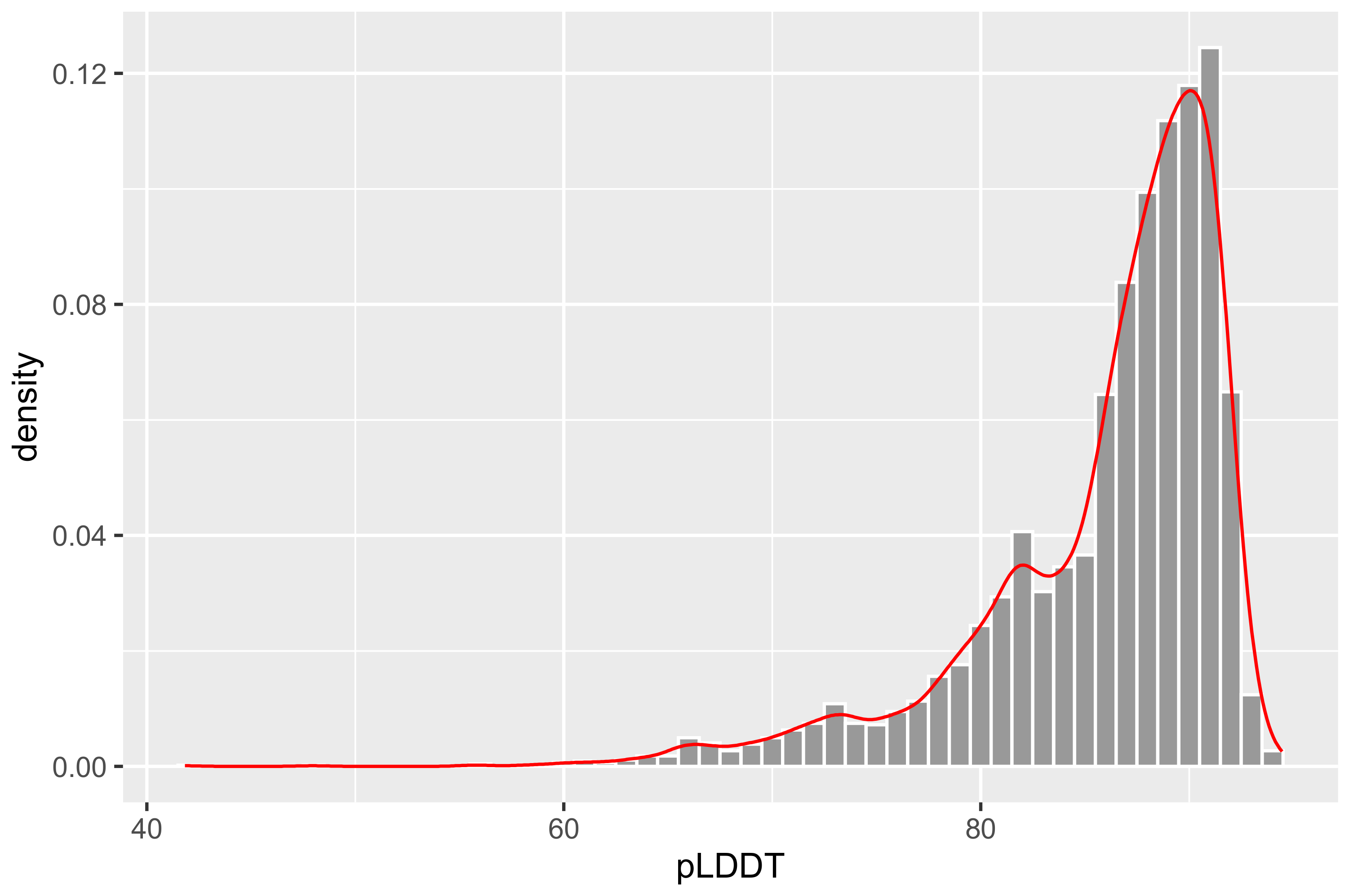
- The model of pLDDT > 90 is considered highly reliable. Due to its high reliability, it should be suitable for any application. It is very helpful for the analysis of protein structure and function.
- The model of pLDDT between 70 and 90 is considered to have high reliability in backbone prediction.
- The model of pLDDT < 70 is considered to have very low reliability or even considered to be unreliable. It should be applied with caution. The lower the pLDDT value, the lower the reliability.
2.4. Virtual Screening of Ligands
2.5. Prediction of Binding Regions and Transmembrane Domains
2.6. Verification of Docking Posture
2.7. Structure Prediction and Ligand Virtual Screening Services
2.8. Database Implementation
3. Results
3.1. Insect Odorant Receptor Sequences
3.2. Three-Dimensional Structure of Odorant Receptor Sequences
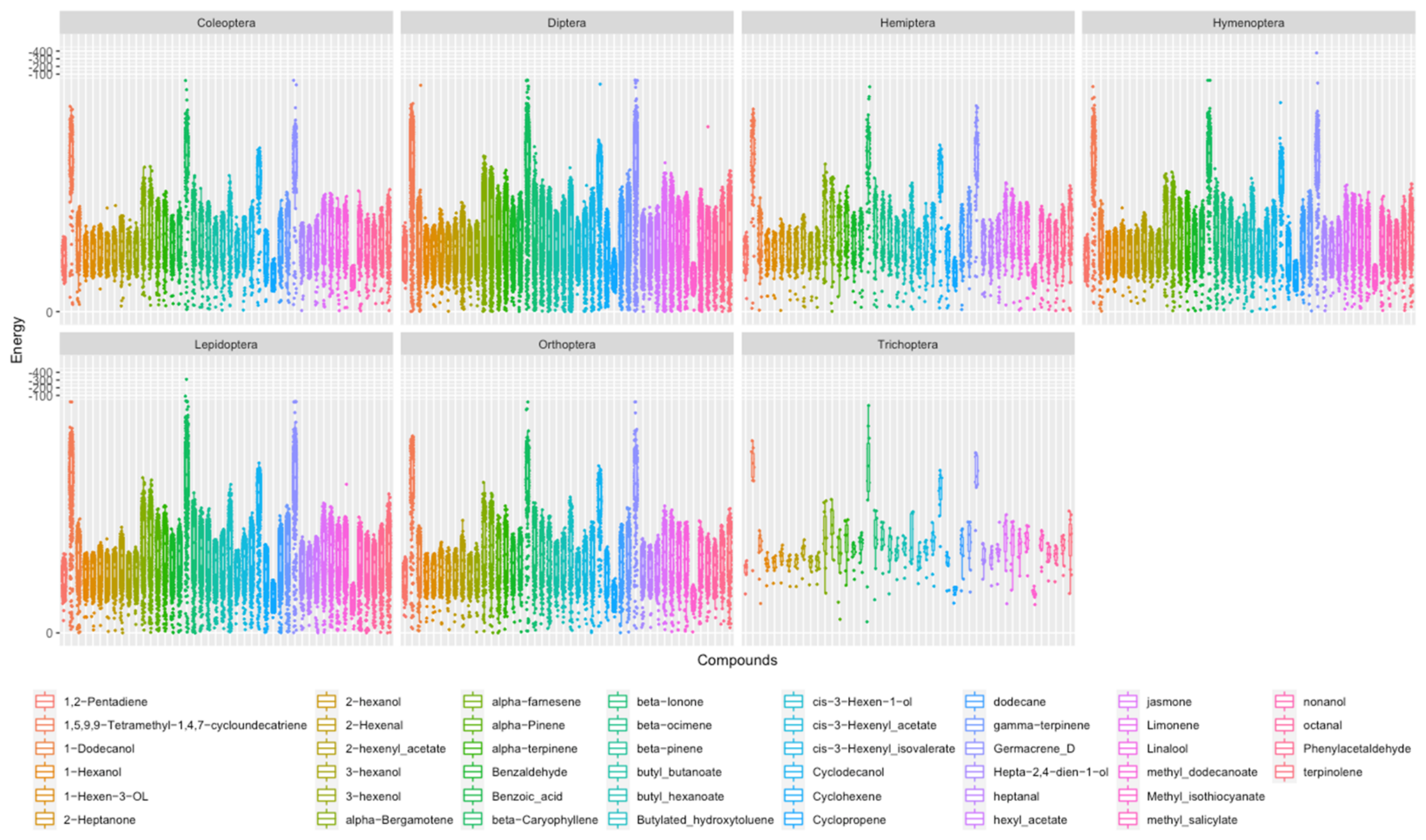
3.3. Virtual Screening of Ligands
3.4. Binding Regions and Transmembrane Domains
3.5. Verification of Docking Posture
3.6. Web Interface and Usage
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, R.T.; Huang, L.Q.; Dong, J.F.; Wang, C.Z. A moth odorant receptor highly expressed in the ovipositor is involved in detecting host-plant volatiles. eLife 2020, 9, e53706. [Google Scholar] [CrossRef] [PubMed]
- Haverkamp, A.; Hansson, B.S.; Knaden, M. Combinatorial codes and labeled lines: How insects use olfactory cues to find and judge food, mates, and oviposition sites in complex environments. Front. Physiol. 2018, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Kannan, K.; Galizia, C.G.; Nouvian, M. Olfactory strategies in the defensive behaviour of insects. Insects 2022, 13, 470. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-J.; Field, L.M.; He, X.L. Insect odorant-binding proteins: Do they offer an alternative pest control strategy? Outlooks Pest Manag. 2010, 21, 31–34. [Google Scholar] [CrossRef]
- Andersson, M.N.; Newcomb, R.D. Pest control compounds targeting insect chemoreceptors: Another silent spring? Front. Ecol. Evol. 2017, 5, 5. [Google Scholar] [CrossRef]
- Konopka, J.K.; Task, D.; Afify, A.; Raji, J.; Deibel, K.; Maguire, S.; Lawrence, R.; Potter, C.J. Olfaction in Anopheles mosquitoes. Chem. Senses 2021, 46, bjab021. [Google Scholar] [CrossRef]
- Mika, K.; Benton, R. Olfactory Receptor Gene Regulation in Insects: Multiple Mechanisms for Singular Expression. Front. Neurosci. 2021, 15, 738088. [Google Scholar] [CrossRef]
- Wicher, D.; Miazzi, F. Functional properties of insect olfactory receptors: Ionotropic receptors and odorant receptors. Cell Tissue Res. 2021, 383, 7–19. [Google Scholar] [CrossRef]
- Fleischer, J.; Pregitzer, P.; Breer, H.; Krieger, J. Access to the odor world: Olfactory receptors and their role for signal transduction in insects. Cell. Mol. Life Sci. 2018, 75, 485–508. [Google Scholar] [CrossRef]
- Zhu, J.; Arena, S.; Spinelli, S.; Liu, D.; Zhang, G.; Wei, R.; Cambillau, C.; Scaloni, A.; Wang, G.; Pelosi, P. Reverse chemical ecology: Olfactory proteins from the giant panda and their interactions with putative pheromones and bamboo volatiles. Proc. Natl. Acad. Sci. USA 2017, 114, E9802–E9810. [Google Scholar] [CrossRef]
- Butterwick, J.A.; Del Mármol, J.; Kim, K.H.; Kahlson, M.A.; Rogow, J.A.; Walz, T.; Ruta, V. Cryo-EM structure of the insect olfactory receptor Orco. Nature 2018, 560, 447–452. [Google Scholar] [CrossRef]
- Kamala Jayanthi, P.D.; Kempraj, V.; Aurade, R.M.; Kumar Roy, T.; Shivashankara, K.S.; Verghese, A. Computational reverse chemical ecology: Virtual screening and predicting behaviorally active semiochemicals for Bactrocera dorsalis. BMC Genom. 2014, 15, 209. [Google Scholar] [CrossRef]
- Manzoni, C.; Kia, D.A.; Vandrovcova, J.; Hardy, J.; Wood, N.W.; Lewis, P.A.; Ferrari, R. Genome, transcriptome and proteome: The rise of omics data and their integration in biomedical sciences. Brief. Bioinform. 2018, 19, 286–302. [Google Scholar] [CrossRef]
- Pulgar-Sánchez, M.; Marrero-Ponce, Y.; Hernández-Lambraño, R.; García-Jacas, C.R.; Martinez-Rios, F. Machine Learning Models and New Computational Tool for the Discovery of Insect Repellents that Interfere with Olfaction; Research Square: Durham, UK, 2022. [Google Scholar]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Baek, M.; Di Maio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef]
- Hopf, T.A.; Morinaga, S.; Ihara, S.; Touhara, K.; Marks, D.S.; Benton, R. Amino acid coevolution reveals three-dimensional structure and functional domains of insect odorant receptors. Nat. Commun. 2015, 6, 6077. [Google Scholar] [CrossRef]
- Dai, J.Q.; Zhen-Yu, L.I.; Chen, H.Y.; Feng, X.; Han, S.C. Isolation and identification of plant volatiles from 3 kinds of cruciferous vegetables. Guangdong Agric. Ences 2011, 26, 816–822. [Google Scholar]
- Zhang, Q.Q.; Chen, H.Y.; Qin, Y.C. Damage from Bemisia tabaci Affects Its Host Plant’s Volatiles and the Parasitism of Encarsia formosa. Chin. J. Biol. Control 2011, 27, 22–26. [Google Scholar]
- Bruce, T.J.A.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects—Finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef]
- Bukovinszky, T.; Gols, R.; Posthumus, M.A.; Vet, L.E.; Van Lenteren, J.C. Variation in plant volatiles and attraction of the parasitoid Diadegma semiclausum (Hellén). J. Chem. Ecol. 2005, 31, 461–480. [Google Scholar] [CrossRef]
- Poelman, E.H.; Bruinsma, M.; Zhu, F.; Weldegergis, B.T.; Boursault, A.E.; Jongema, Y.; van Loon, J.J.; Vet, L.E.; Harvey, J.A.; Dicke, M. Hyperparasitoids use herbivore-induced plant volatiles to locate their parasitoid host. PLoS Biol. 2012, 10, e1001435. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Uda, Y.; Ono, Y.; Nakagawa, T.; Suwa, M.; Yamaoka, R.; Touhara, K. Highly selective tuning of a silkworm olfactory receptor to a key mulberry leaf volatile. Curr. Biol. 2009, 19, 881–890. [Google Scholar] [CrossRef] [PubMed]
- SOLÉ, J.; SANS, A.; RIBA, M.; GUERRERO, A. Behavioural and electrophysiological responses of the European corn borer Ostrinia nubilalis to host-plant volatiles and related chemicals. Physiol. Entomol. 2010, 35, 354–363. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- States, D.J.; Gish, W. Combined use of sequence similarity and codon bias for coding region identification. J. Comput. Biol. 1994, 1, 39–50. [Google Scholar] [CrossRef]
- Madden, T.L.; Tatusov, R.L.; Zhang, J. Applications of network BLAST server. Methods Enzymol. 1996, 266, 131–141. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Zhang, J.; Madden, T.L. PowerBLAST: A new network BLAST application for interactive or automated sequence analysis and annotation. Genome Res. 1997, 7, 649–656. [Google Scholar] [CrossRef]
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Schäffer, A.A.; Agarwala, R.; Altschul, S.F.; Lipman, D.J.; Madden, T.L. Domain enhanced lookup time accelerated BLAST. Biol. Direct 2012, 7, 12. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Thierry-Mieg, J.; Thierry-Mieg, D.; Busby, B.; Madden, T.L. Magic-BLAST, an accurate RNA-seq aligner for long and short reads. BMC Bioinform. 2019, 20, 405. [Google Scholar] [CrossRef]
- Sehnal, D.; Bittrich, S.; Deshpande, M.; Svobodová, R.; Berka, K.; Bazgier, V.; Velankar, S.; Burley, S.K.; Koča, J.; Rose, A.S. Mol* Viewer: Modern web app for 3D visualization and analysis of large biomolecular structures. Nucleic Acids Res. 2021, 49, W431–w437. [Google Scholar] [CrossRef]
- Touw, W.G.; Baakman, C.; Black, J.; te Beek, T.A.; Krieger, E.; Joosten, R.P.; Vriend, G. A series of PDB-related databanks for everyday needs. Nucleic Acids Res. 2015, 43, D364–D368. [Google Scholar] [CrossRef]
- Kabsch, W.; Sander, C. Dictionary of protein secondary structure: Pattern recognition of hydrogen-bonded and geometrical features. Biopolymers 1983, 22, 2577–2637. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Santos-Martins, D.; Eberhardt, J.; Bianco, G.; Solis-Vasquez, L.; Ambrosio, F.A.; Koch, A.; Forli, S. D3R Grand Challenge 4: Prospective pose prediction of BACE1 ligands with AutoDock-GPU. J. Comput. Aided Mol. Des. 2019, 33, 1071–1081. [Google Scholar] [CrossRef]
- Forli, S.; Olson, A.J. A force field with discrete displaceable waters and desolvation entropy for hydrated ligand docking. J. Med. Chem. 2012, 55, 623–638. [Google Scholar] [CrossRef]
- Rose, A.S.; Bradley, A.R.; Valasatava, Y.; Duarte, J.M.; Prlic, A.; Rose, P.W. NGL viewer: Web-based molecular graphics for large complexes. Bioinformatics 2018, 34, 3755–3758. [Google Scholar] [CrossRef]
- Rose, A.S.; Hildebrand, P.W. NGL Viewer: A web application for molecular visualization. Nucleic Acids Res. 2015, 43, W576–W579. [Google Scholar] [CrossRef] [PubMed]
- Ravindranath, P.A.; Forli, S.; Goodsell, D.S.; Olson, A.J.; Sanner, M.F. AutoDockFR: Advances in Protein-Ligand Docking with Explicitly Specified Binding Site Flexibility. PLoS Comput. Biol. 2015, 11, e1004586. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Stoffler, D.; Sanner, M. Hierarchical and multi-resolution representation of protein flexibility. Bioinformatics 2006, 22, 2768–2774. [Google Scholar] [CrossRef] [PubMed]
- Schmidtke, P.; Bidon-Chanal, A.; Luque, F.J.; Barril, X. MDpocket: Open-source cavity detection and characterization on molecular dynamics trajectories. Bioinformatics 2011, 27, 3276–3285. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Hallgren, J.; Tsirigos, K.D.; Pedersen, M.D.; Almagro Armenteros, J.J.; Marcatili, P.; Nielsen, H.; Krogh, A.; Winther, O. DeepTMHMM predicts alpha and beta transmembrane proteins using deep neural networks. bioRxiv, 2022. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 27–38. [Google Scholar] [CrossRef]
- Guruprasad, K.; Reddy, B.V.; Pandit, M.W. Correlation between stability of a protein and its dipeptide composition: A novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. 1990, 4, 155–161. [Google Scholar] [CrossRef]
- Smoluch, M.; Mielczarek, P.; Drabik, A.; Silberring, J. 5—Online and Offline Sample Fractionation. In Proteomic Profiling and Analytical Chemistry, 2nd ed.; Ciborowski, P., Silberring, J., Eds.; Elsevier: Boston, MA, USA, 2016; pp. 63–99. [Google Scholar]
- Mariani, V.; Biasini, M.; Barbato, A.; Schwede, T. lDDT: A local superposition-free score for comparing protein structures and models using distance difference tests. Bioinformatics 2013, 29, 2722–2728. [Google Scholar] [CrossRef]
- Tunyasuvunakool, K.; Adler, J.; Wu, Z.; Green, T.; Zielinski, M.; Žídek, A.; Bridgland, A.; Cowie, A.; Meyer, C.; Laydon, A.; et al. Highly accurate protein structure prediction for the human proteome. Nature 2021, 596, 590–596. [Google Scholar] [CrossRef]
- Del Mármol, J.; Yedlin, M.A.; Ruta, V. The structural basis of odorant recognition in insect olfactory receptors. Nature 2021, 597, 126–131. [Google Scholar] [CrossRef]
- Revadi, S.V.; Giannuzzi, V.A.; Rossi, V.; Hunger, G.M.; Conchou, L.; Rondoni, G.; Conti, E.; Anderson, P.; Walker, W.B.; Jacquin-Joly, E.; et al. Stage-specific expression of an odorant receptor underlies olfactory behavioral plasticity in Spodoptera littoralis larvae. BMC Biol. 2021, 19, 231. [Google Scholar] [CrossRef]
- Liu, N.Y.; Zhu, J.Y.; Zhang, T.; Dong, S.L. Characterization of two odorant binding proteins in Spodoptera exigua reveals functional conservation and difference. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2017, 213, 20–27. [Google Scholar] [CrossRef]
- Liu, H.; Xiu, C.; Zhang, T.; Lu, Y. Odor Perception in the Cotton Bollworm, Helicoverpa armigera, Exposed to Juglans regia, a Marginal Host Plant. J. Chem. Ecol. 2022, 48, 618–627. [Google Scholar] [CrossRef]
- Hou, X.; Zhang, D.D.; Yuvaraj, J.K.; Corcoran, J.A.; Andersson, M.N.; Löfstedt, C. Functional characterization of odorant receptors from the moth Eriocrania semipurpurella: A comparison of results in the Xenopus oocyte and HEK cell systems. Insect Biochem. Mol. Biol. 2020, 117, 103289. [Google Scholar] [CrossRef]
- Xu, L.; Jiang, H.B.; Tang, K.Y.; Yan, Y.; Schetelig, M.F.; Wang, J.J. CRISPR-mediated mutagenesis of the odorant receptor co-receptor (Orco) gene disrupts olfaction-mediated behaviors in Bactrocera dorsalis. Insect Sci. 2022, 29, 1275–1286. [Google Scholar] [CrossRef]
- Bezerra-Silva, P.C.; Dutra, K.A.; Santos, G.K.; Silva, R.C.; Iulek, J.; Milet-Pinheiro, P.; Navarro, D.M. Evaluation of the Activity of the Essential Oil from an Ornamental Flower against Aedes aegypti: Electrophysiology, Molecular Dynamics and Behavioral Assays. PLoS ONE 2016, 11, e0150008. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Q.; Guo, J.; Li, J.; Wang, J.; Wen, M.; Zhao, H.; Ren, B. Molecular basis of peripheral olfactory sensing during oviposition in the behavior of the parasitic wasp Anastatus japonicus. Insect Biochem. Mol. Biol. 2017, 89, 58–70. [Google Scholar] [CrossRef]
- Chen, Z.L.; Huang, C.; Li, X.S.; Li, G.C.; Yu, T.H.; Fu, G.J.; Zhang, X.; Song, C.; Bai, P.H.; Cao, L.; et al. Behavioural regulator and molecular reception of a double-edge-sword hunter beetle. Pest Manag. Sci. 2022, 78, 2693–2703. [Google Scholar] [CrossRef]
- Bichão, H.; Borg-Karlson, A.K.; Araújo, J.; Mustaparta, H. Five types of olfactory receptor neurons in the strawberry blossom weevil Anthonomus rubi: Selective responses to inducible host-plant volatiles. Chem. Senses 2005, 30, 153–170. [Google Scholar] [CrossRef]
- Gu, S.H.; Wang, W.X.; Wang, G.R.; Zhang, X.Y.; Guo, Y.Y.; Zhang, Z.; Zhou, J.J.; Zhang, Y.J. Functional characterization and immunolocalization of odorant binding protein 1 in the lucerne plant bug, Adelphocoris lineolatus (GOEZE). Arch. Insect Biochem. Physiol. 2011, 77, 81–99. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Duan, H.; Wang, Q.; Xiao, Y.; Wang, Q.; Xiao, Q.; Sun, L.; Zhang, Y. Key Amino Residues Determining Binding Activities of the Odorant Binding Protein AlucOBP22 to Two Host Plant Terpenoids of Apolygus lucorum. J. Agric. Food Chem. 2019, 67, 5949–5956. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, H.; Wang, Z.; Liu, J.; Zhang, X.; Li, C.; Zeng, X. Molecular Identification, Expression, and Functional Analysis of a General Odorant-Binding Protein 1 of Asian Citrus Psyllid. Environ. Entomol. 2019, 48, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Matthiessen, J.N.; Shackleton, M.A. Advantageous attributes of larval whitefringed weevil, Naupactus leucoloma (Coleoptera: Curculionidae) for bioassaying soil fumigants, and responses to pure and plant-derived isothiocyanates. Bull. Entomol. Res. 2000, 90, 349–355. [Google Scholar] [CrossRef]
- Scharf, M.E.; Nguyen, S.N.; Song, C. Evaluation of volatile low molecular weight insecticides using Drosophila melanogaster as a model. Pest Manag. Sci. 2006, 62, 655–663. [Google Scholar] [CrossRef]
- Batra, S.; Corcoran, J.; Zhang, D.D.; Pal, P.; K P, U.; Kulkarni, R.; Löfstedt, C.; Sowdhamini, R.; Olsson, S.B. A Functional Agonist of Insect Olfactory Receptors: Behavior, Physiology and Structure. Front. Cell. Neurosci. 2019, 13, 134. [Google Scholar] [CrossRef]
- Wang, C.F.; Yang, K.; You, C.X.; Zhang, W.J.; Guo, S.S.; Geng, Z.F.; Du, S.S.; Wang, Y.Y. Chemical Composition and Insecticidal Activity of Essential Oils from Zanthoxylum dissitum Leaves and Roots against Three Species of Storage Pests. Molecules 2015, 20, 7990–7999. [Google Scholar] [CrossRef]
- Suckling, D.M.; Twidle, A.M.; Gibb, A.R.; Manning, L.M.; Mitchell, V.J.; Sullivan, T.E.; Wee, S.L.; El-Sayed, A.M. Volatiles from apple trees infested with light brown apple moth larvae attract the parasitoid Dolichogenidia tasmanica. J. Agric. Food Chem. 2012, 60, 9562–9566. [Google Scholar] [CrossRef]
- Mozuraitis, R.; Stranden, M.; Ramirez, M.I.; Borg-Karlson, A.K.; Mustaparta, H. (-)-Germacrene D increases attraction and oviposition by the tobacco budworm moth Heliothis virescens. Chem. Senses 2002, 27, 505–509. [Google Scholar] [CrossRef]
- Stranden, M.; Liblikas, I.; König, W.A.; Almaas, T.J.; Borg-Karlson, A.K.; Mustaparta, H. (-)-Germacrene D receptor neurones in three species of heliothine moths: Structure-activity relationships. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 2003, 189, 563–577. [Google Scholar] [CrossRef]
- Basciu, A.; Koukos, P.I.; Malloci, G.; Bonvin, A.; Vargiu, A.V. Coupling enhanced sampling of the apo-receptor with template-based ligand conformers selection: Performance in pose prediction in the D3R Grand Challenge 4. J. Comput. Aided Mol. Des. 2020, 34, 149–162. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Nguyen, T.H.; Pham, T.N.H.; Huy, N.T.; Bay, M.V.; Pham, M.Q.; Nam, P.C.; Vu, V.V.; Ngo, S.T. Autodock Vina Adopts More Accurate Binding Poses but Autodock4 Forms Better Binding Affinity. J. Chem. Inf. Model. 2020, 60, 204–211. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Id | Name | Abbreviation |
|---|---|---|
| 1 | 1,2-Pentadiene | PEN |
| 2 | 2-Hexenal | HX2 |
| 3 | beta-Caryophyllene | CAY |
| 4 | Linalool | LIN |
| 5 | 1,5,9,9-Tetramethyl-1,4,7-cycloundecatriene | TEC |
| 6 | alpha-Bergamotene | BER |
| 7 | beta-Ionone | ION |
| 8 | Cyclohexene | CYC |
| 9 | Methyl isothiocyanate | MEI |
| 10 | 1-Dodecanol | DOD |
| 11 | alpha-Farnesene | FAR |
| 12 | Butylated hydroxytoluene | BUH |
| 13 | Cyclopropene | CYL |
| 14 | Phenylacetaldehyde | PHE |
| 15 | 1-Hexanol | HE1 |
| 16 | alpha-Pinene | PIN |
| 17 | cis-3-Hexen-1-ol | CHO |
| 18 | Germacrene D | GED |
| 19 | 1-Hexen-3-OL | HEO |
| 20 | Benzaldehyde | BEN |
| 21 | cis-3-Hexenyl acetate | CHA |
| 22 | Hepta-2,4-dien-1-ol | HDO |
| 23 | 2-Heptanone | HEP |
| 24 | Benzoic acid | BEA |
| 25 | cis-3-Hexenyl isovalerate | CHI |
| 26 | Limonene | LIM |
| 27 | Cyclodecanol | CYO |
| 28 | alpha-terpinene | TER |
| 29 | jasmone | JAS |
| 30 | methyl_dodecanoate | MDO |
| 31 | 2-hexenyl_acetate | HEX |
| 32 | dodecane | DOE |
| 33 | methyl_salicylate | MES |
| 34 | 2-hexanol | HEA |
| 35 | hexyl_acetate | HET |
| 36 | 3-hexenol | HX3 |
| 37 | terpinolene | TEP |
| 38 | 3-hexanol | HEN |
| 39 | octanal | OCT |
| 40 | nonanol | NOA |
| 41 | heptanal | HEL |
| 42 | butyl_butanoate | BUB |
| 43 | beta-pinene | BPI |
| 44 | butyl_hexanoate | BUE |
| 45 | gamma-terpinene | GAM |
| 46 | beta-ocimene | OCI |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, S.; Qian, K.; He, L.; Zhang, Z. iORandLigandDB: A Website for Three-Dimensional Structure Prediction of Insect Odorant Receptors and Docking with Odorants. Insects 2023, 14, 560. https://doi.org/10.3390/insects14060560
Jin S, Qian K, He L, Zhang Z. iORandLigandDB: A Website for Three-Dimensional Structure Prediction of Insect Odorant Receptors and Docking with Odorants. Insects. 2023; 14(6):560. https://doi.org/10.3390/insects14060560
Chicago/Turabian StyleJin, Shuo, Kun Qian, Lin He, and Zan Zhang. 2023. "iORandLigandDB: A Website for Three-Dimensional Structure Prediction of Insect Odorant Receptors and Docking with Odorants" Insects 14, no. 6: 560. https://doi.org/10.3390/insects14060560
APA StyleJin, S., Qian, K., He, L., & Zhang, Z. (2023). iORandLigandDB: A Website for Three-Dimensional Structure Prediction of Insect Odorant Receptors and Docking with Odorants. Insects, 14(6), 560. https://doi.org/10.3390/insects14060560