
Honey Bee Colonies (Apis mellifera L.) Perform Orientation Defensiveness That Varies among Bred Lines


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Breeding and Placement of Honey Bee Colonies at the Experimental Apiary
2.2. Chemical, Physical, and Visual Stimuli Application (Assays)
2.3. Assessment of Defensiveness and Orientation in Honey Bee Colonies
2.4. Data Analysis
3. Results
3.1. Experiment 1: Evaluating the Defensive Behavior of Honey Bees among Bred Lines
3.1.1. Effects of Assay (Alarm Pheromone, Isopentyl Acetate/Paraffin, and Empty Suedes) on Time and Intensity of Recruitment of Honey Bees on the Suede
3.1.2. Behavioral Responses of Honey Bee Colonies to Different Assays
3.1.3. Variation in the Aggressive Behavior among Honey Bee Bred Lines
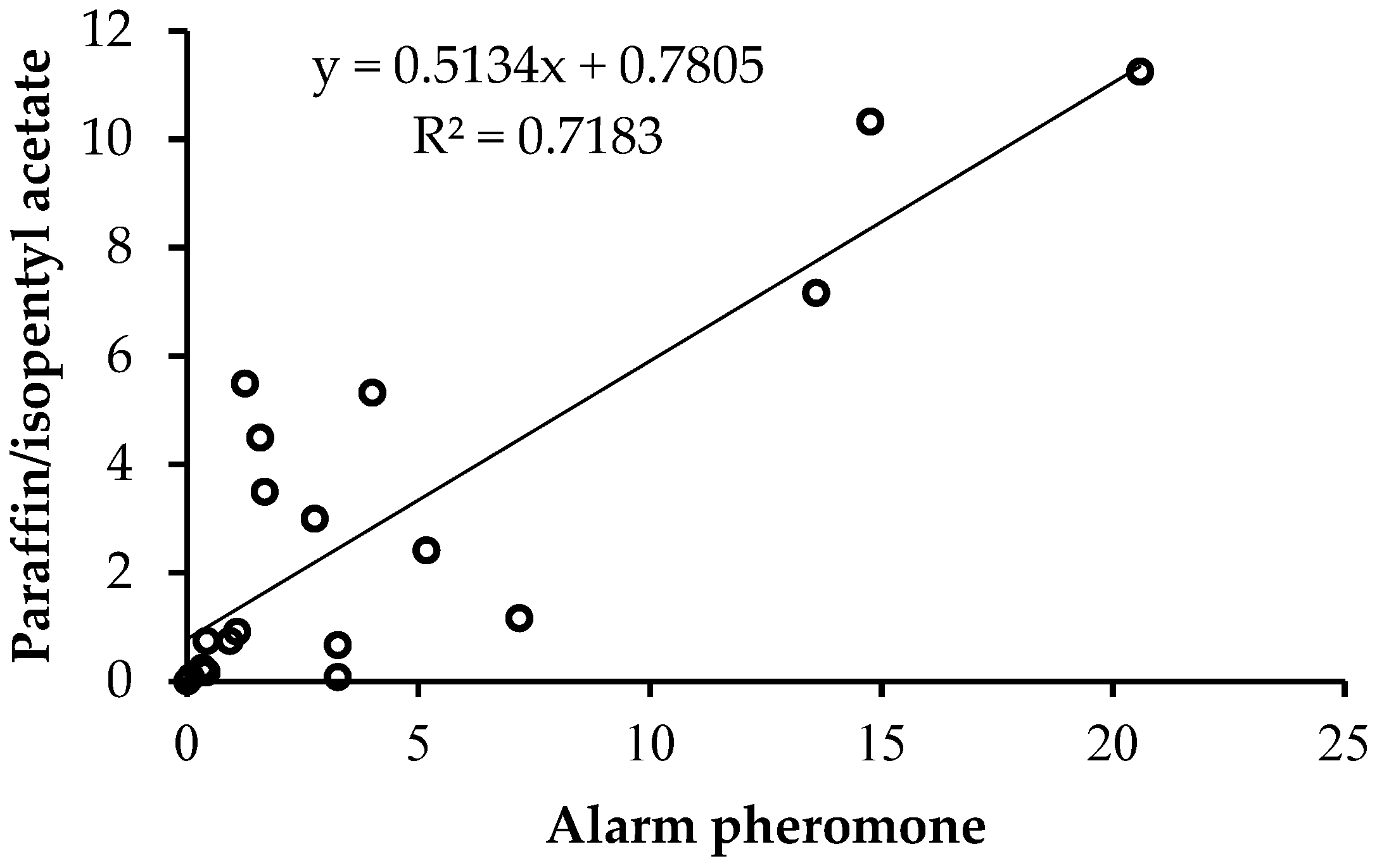
3.2. Experiment 2: Evaluating Orientation Defensiveness among Honey Bee Bred Lines
Orientation Aggressiveness among Bred Lines of Honey Bee Colonies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alcock, J. Animal Behavior: An Evolutionary Approach, 9th ed.; Sinauer Associates: Sunderland, MA, USA, 2009; p. 546. [Google Scholar]
- Collins, A.M.; Rinderer, T.E. Effect of empty comb on defensive behavior of honeybees. J. Chem. Ecol. 1985, 11, 333–338. [Google Scholar] [CrossRef]
- Breed, M.D.; Guzmán-Novoa, E.; Hunt, G.J. Defensive behavior of honey bees: Organization, genetics, and comparisons with other bees. Annu. Rev. Entomol. 2004, 49, 271–298. [Google Scholar] [CrossRef]
- Collins, A.M.; Rinderer, T.E.; Tucker, K.W. Colony defense of two honeybee types and their hybrid 1naturally mated queens. J. Apic. Res. 1988, 27, 137–140. [Google Scholar] [CrossRef]
- Villa, J.D. Defensive behavior of Africanized and European honeybees at two elevations in Colombia. J. Apic. Res. 1988, 27, 141–145. [Google Scholar] [CrossRef]
- Page, R.E.; Robinson, G.E.; Fondryk, M.K.; Nasr, M.E. Effects of worker genotypic diversity on honeybee colony development and behavior Apis mellifera L. Behav. Ecol. Sociobiol. 1995, 36, 387–396. [Google Scholar] [CrossRef]
- Page, R.E.; Rueppell, O.; Amdam, G.V. Genetics of reproduction and regulation of honeybee (Apis mellifera L.) social behavior. Annu. Rev. Genet. 2012, 46, 97–119. [Google Scholar] [CrossRef]
- Avalos, A.; Fang, M.; Pan, H.; Lluch, A.R.; Lipka, A.E.; Zhao, S.D.; Hudson, M.E. Genomic regions influencing aggressive behavior in honey bees are defined by colony allele frequencies. Proc. Natl. Acad. Sci. USA 2020, 117, 17135–17141. [Google Scholar] [CrossRef] [PubMed]
- Spivak, M.; Gilliam, M. Hygienic behavior of honey bees and its application for control of brood diseases and varroa. Bee World 1998, 79, 124–134. [Google Scholar] [CrossRef]
- Harrison, J.W. Bees with Varroa Sensitive Hygiene preferentially remove mite infested pupae aged ≤ five days post capping. J. Apic. Res. 2007, 46, 134–139. [Google Scholar] [CrossRef]
- Bogdanov, S. Beeswax: Production, Properties Composition and control. Bee Prod. Sci. 2016, 1, 1–18. [Google Scholar]
- Russell, R. Beekeeping, poverty alleviation and conservation in Imadiala, Madagascar. Bees Dev. J. 2008, 84, 6–7. [Google Scholar]
- Ngama, S.; Korte, L.; Bindelle, J.; Vermeulen, C.; Poulsen, J.R. How bees deter elephants: Beehive trials with forest elephants (Loxodonta africanacyclotis) in Gabon. PLoS ONE 2016, 11, e0155690. [Google Scholar] [CrossRef]
- Cook, R.M.; Parrini, F.; King, L.E.; Witowski, E.T.F.; Henley, M.D. African honeybees as a mitigation method for elephant impact on trees. Biol. Conserv. 2017, 217, 329–336. [Google Scholar] [CrossRef]
- King, L.E.; Lala, F.; Nzumu, H.; Mwambingu, E.; Douglashamilton, I. Beehive fences as a multidimensional conflict-mitigation tool for farmers coexisting with elephants. Conserv. Biol. 2017, 31, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Isaac, B.D.; Robert, B.W.; Patrick, P. Diurnality in the defensive behavior of African honeybees Apis mellifera adansonii and implications for their potential efficacy in beehive fences. Oryx 2022, 2022, 1–7. [Google Scholar] [CrossRef]
- Mmbaga, N.E.; Munishi, L.K.; Treydte, A.C. How dynamics and drivers of land use/land cover change impact elephant conservation and agricultural livelihood development in Rombo, Tanzania. J. Land Use Sci. 2017, 12, 168–181. [Google Scholar] [CrossRef]
- Wright, M.G.; Spencer, C.; Cook, R.M.; Henley, M.D.; North, W.; Mafra-Neto, A. African bush elephants respond to a honeybee alarm pheromone blend. Curr. Biol. 2018, 28, R778–R780. [Google Scholar] [CrossRef] [PubMed]
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1987; p. 294. [Google Scholar]
- Rittschof, C.C.; Coombs, C.B.; Frazier, M.; Grozinger, C.M.; Robinson, G.E. Early life experience affects honey bee aggression and resilience to immune challenge. Sci. Rep. 2015, 5, 15572. [Google Scholar] [CrossRef] [PubMed]
- Alaux, C.; Sinha, S.; Hasadsri, L.; Hunt, G.J.; Guzmán-Novoa, E.; DeGrandi-Hoffman, G.; Uribe-rubio, J.L.; Southey, B.R.; Rodriguez-Zas, S.; Robinson, G.E. Honey bee aggression supports a link between gene regulation and behavioral evolution. Proc. Natl. Acad. Sci. USA 2009, 106, 15400–15405. [Google Scholar] [CrossRef]
- Guzmán-Novoa, E.; Page, R.E. Genetic dominance and worker interactions affect honey bee colony defense. Behav. Ecol. 1994, 5, 91–97. [Google Scholar] [CrossRef]
- Hunt, G.J.; Guzmán-Novoa, E.; Uribe-rubio, J.L.; Prieto-Merlos, D. Genotype-environment interactions in honeybee guarding behavior. Anim. Bahav. 2003, 66, 459–467. [Google Scholar] [CrossRef]
- Hunt, G.J. Flight and fight: A comparative view of the neurophysiology and genetics of honey bee defensive behavior. J. Insect Physiol. 2007, 53, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Burrell, D.; Smith, B.H. Age-related but not caste-related regulation of abdominal mechanisms underlying the sting reflex of the honey bee, Apis mellifera. J. Comp. Physiol. A 1994, 174, 581–592. [Google Scholar] [CrossRef]
- Collins, A.M.; Rinderer, T.E.; Harbo, J.R.; Bolten, A.B. Colony defense by Africanized and European honey bees. Science 1982, 218, 72–74. [Google Scholar] [CrossRef]
- Guzmán-Novoa, E.; Page, R.E. Backcrossing Africanized honey bee (Apis mellifera L.) queens to European drones reduces colony defensive behavior. Ann. Entomol. Soc. Am. 1993, 86, 352–355. [Google Scholar] [CrossRef]
- Guzmán-Novoa, E.; Page, R.E. The impact of Africanized bees on Mexican beekeeping. Am. Bee J. 1994, 134, 101–106. [Google Scholar]
- Guzmán-Novoa, E.; Hunt, G.J.; Page, R.E.; Fondrk, M.K. Genetic correlations among honey bee (Hymenoptera: Apidae) behavioral characteristics and wing length. Ann. Entomol. Soc. Am. 2002, 95, 402–406. [Google Scholar] [CrossRef]
- Ilyasov, R.A.; Poskryakov, A.V.; Petukhov, A.V.; Nikolenko, A.G. Molecular genetic analysis of five extant reserves of black honeybee Apismelliferamellifera in the Urals and the Volga region. Russ. J. Genet. 2016, 52, 828–839. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Collins, A.M.; Martin, J.H.; Schmidt, J.O.; Spangler, H.G. Nest defense behavior in colonies from crosses between Africanized and European honeybees (Apismelli(era L.) (Hymenoptera: Apidae). J. Insect Behav. 1998, 11, 37–45. [Google Scholar] [CrossRef]
- Guzmán-Novoa, E.; Hunt, G.J.; Uribe, J.L.; Smith, C.; Arechavaleta-Velasco, M.E. Confirmation of QTL effects and evidence of genetic dominance of honey bee defensive behavior: Results of colony and individual behavioral assays. Behav. Genet. 2002, 32, 95–102. [Google Scholar] [CrossRef]
- Winston, M.L. Killer Bees: The Africanized Honey Bee in the Americas; Harvard University Press: Cambridge, MA, USA, 1992; p. 162. [Google Scholar]
- Collins, A.M.; Kubasek, K.J. Field test of honey bee (Hymenoptera: Apidae) colony defensive behavior. Ann. Entomol. Soc. Am. 1982, 75, 383–387. [Google Scholar] [CrossRef]
- Plate, M.; Bernstein, R.; Hppe, A.; Bienefelt, K. The importance of controlled mating in honey bee breeding. Genet. Sel. Evol. 2019, 51, 74. [Google Scholar] [CrossRef] [PubMed]
- Doolittle, G.M. Scientific Queen-Rearing as Practically Applied; Being a Method by which the Best of Queen-Bees are Reared in Perfect Accord with Nature’s Ways: For the Amateur and Veteran in Beekeeping, 6th ed.; American Bee Journal: Hamilton, IL, USA, 1915; p. 126. [Google Scholar]
- Collins, A.M.; Rinderer, T.E.; Tucker, K.W.; Sylvester, H.A.; Lackett, J.J. A model of honeybee defensive behavior. J. Apic. Res. 1980, 19, 224–231. [Google Scholar] [CrossRef]
- Blum, M.S.; Fales, H.M.; Tucker, K.W.; Collins, A.M. Chemistry of the sting apparatus of the worker honeybee. J. Apic. Res. 1978, 17, 218–221. [Google Scholar] [CrossRef]
- Johnson, L.K.; Haynes, L.W.; Carlson, M.A.; Fortnum, H.A.; Gorgas, D.L. Alarm substances of the stingless bee, Trigona silvestriana. J. Chem. Ecol. 1985, 11, 409–416. [Google Scholar] [CrossRef]
- Wilson-Rich, N.; Spivak, M.; Fefferman, N.H.; Starks, P.T. Genetic, individual and group facilitation of disease resistance insect societies. Annu. Rev. Entomol. 2009, 54, 405–423. [Google Scholar] [CrossRef] [PubMed]
- Wager, B.R.; Breed, M.D. Does honey bee sting alarm pheromone gives orientation information to defensive bees? Behavior 2000, 93, 1329–1332. [Google Scholar] [CrossRef]
- Boch, R.; Shearer, D.A.; Stone, B.C. Identification of iso-amyl acetate as an active component in the sting pheromone of the honey bee. Nature 1962, 195, 1018–1020. [Google Scholar] [CrossRef]
- Collins, A.M.; Rinderer, T.E.; Daly, H.V. Alarm pheromone production by two honey bee (Apis mellifera) types. J. Chem. Ecol. 1989, 15, 1747–1756. [Google Scholar] [CrossRef] [PubMed]
- Lensky, Y.; Cassier, P.; Tel-Zur, D. The setaceous membrane of honey bee (Apis mellifera) workers sting apparatus: Structure and alarm pheromone distribution. J. Insect. Physiol. 1995, 41, 589–595. [Google Scholar] [CrossRef]
- Lopez-Incera, A.; Nouvian, M.; Ried, K.; Muller, T.; Briegel, H.J. Honey bee communication during collective defense is shaped by predation. BMC Biol. 2021, 19, 106. [Google Scholar] [CrossRef] [PubMed]
- Petrov, T.; Hajnal, M.; Klein, J.; Šafránek, D.; Nouvian, M. Extracting individual characteristics from population data reveals a negative social effect during honey beedefence. PLoS. Comput. Biol. 2022, 18, e1010305. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, E.; Agra, M.N.; Garcia, C.; Gennari, G.; Maldonado, L.; Rodriguez, G.A.; Palacio, M.A.; Scannapieco, A.C.; Lanzavecchia, S.B. Defensive behavior and morphometric variation in Apis mellifera colonies from two different agro-ecological zones of North-Western Argentina. Front. Ecol. Evol. 2021, 9, 590225. [Google Scholar] [CrossRef]
- Harrison, J.W.; Palmer, J.H.; Rittschof, C.C. Altering social cue perception impacts honey bee aggression with minimal impacts on aggression-related brain gene expression. Sci. Rep. 2019, 9, 14642. [Google Scholar] [CrossRef] [PubMed]
- Mangier, B.; Ekszterowicz, G.; Laurent, J.; Rival, M.; Pfister, F. Bee hive traffic monitoring by tracking bee flight paths. In Proceedings of the 13th Internal Joint Conference on Computer Vision, Imaging and Computer Graphics Theory and Applications, Funchal, Madeira, Portugal, 27–29 January 2018; pp. 563–571. [Google Scholar] [CrossRef]
- Scheiner, R.; Abramson, C.I.; Brodschneider, R.; Crailsheim, K.; Farina, W.M.; Fuchs, S.; Grünewald, B.; Hahshold, S.; Karrer, M.; Koeniger, G.; et al. Standard methods for behavioral studies of Apis mellifera. J. Apic. Res. 2013, 52, 1–58. [Google Scholar] [CrossRef]
- Boch, R.; Shearer, D.A.; Petrasovits, A. Efficacies of two alarm substances of the honeybee. J. Insect Physiol. 1970, 16, 17–24. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Type | Volume (mL) | Volume (µL) | Percentage Composition (%) |
|---|---|---|---|
| n-Butyl acetate | 0.15 | 150 | 1.5 |
| Isopentyl acetate | 3.2 | 3200 | 32 |
| Isopentyl alcohol | 1.5 | 1500 | 15 |
| n-Hexyl acetate | 0.4 | 400 | 4 |
| n-Octyl acetate | 1.7 | 1700 | 17 |
| 2-Nonanol | 1.0 | 1000 | 10 |
| Benzyl acetate | 1.6 | 1600 | 16 |
| Benzyl alcohol | 0.45 | 450 | 4.5 |
| Paraffin oil | 90 | 90,000 | 900 |
| Total = 100 mL | 100,000 (µL) | 1000 |
| Bred Lines | Chemical Assay | Control | |
|---|---|---|---|
| Alarm Pheromone (Time/s) (Mean ± SE) | Paraffin/Isopentyl Acetate (Time/s) (Mean ± SE) | Empty Suedes (Time/s) (Mean ± SE) | |
| A | 6.48 ± 0.23 Aa | 7.79 ± 0.21 Aa | 21.43 ± 2.26 Ab |
| B | 6.77 ± 0.26 Aa | 7.87 ± 0.26 Aa | 21.06 ± 2.2 4Ab |
| C | 6.1 ± 0.24 Aa | 7.23 ± 0.24 ABa | 19.56 2.28 Ab |
| D | 7.02 ± 0.27 Aa | 6.58 ± 0.22 BCa | 21.5 ± 3.78 Ab |
| E | 6.21 ± 0.24 Aa | 5.94 ± 0.25 Ca | 20.81 ± 2.5 Ab |
| Recruitment intensity | High | Average | Very low |
| Bred Lines | Alarm Pheromone | Paraffin/Isopentyl Acetate | Control (No Chemical) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| No. of Stings (Mean ± SE) | F-Value | p-Value | No. of Stings (Mean ± SE) | F-Value | p-Value | No. of Stings (Mean ± SE) | F-Value | p-Value | |
| A | 2.21 ± 0.77 ab | 0.462 | 0.633 | 2.62 ± 0.84 a | 0.136 | 0.873 | 0.12 ± 0.07 abc | 1.898 | 0.162 |
| B | 1.08± 0.56 b | 0.586 | 0.561 | 1.04 ± 0.37 a | 1.23 | 0.302 | 0.02 ± 0.02 c | 1 | 0.376 |
| C | 5.69± 1.75 ab | 1.152 | 0.325 | 3 ± 1.14 a | 0.86 | 0.43 | 0.46 ± 0.25 a | 0.451 | 0.64 |
| D | 4.69± 1.91 ab | 0.469 | 0.629 | 2.85 ± 1.43 a | 0.317 | 0.73 | 0.04 ± 0.03 bc | 0.5 | 0.61 |
| E | 6.98± 2.12 a | 3.655 | 0.034 | 4.98 ± 1.07 a | 3.368 | 0.043 | 0.4 ± 0.15 ab | 0.07 | 0.932 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akongte, P.N.; Park, B.-S.; Kim, D.-W.; Choi, Y.-S. Honey Bee Colonies (Apis mellifera L.) Perform Orientation Defensiveness That Varies among Bred Lines. Insects 2023, 14, 546. https://doi.org/10.3390/insects14060546
Akongte PN, Park B-S, Kim D-W, Choi Y-S. Honey Bee Colonies (Apis mellifera L.) Perform Orientation Defensiveness That Varies among Bred Lines. Insects. 2023; 14(6):546. https://doi.org/10.3390/insects14060546
Chicago/Turabian StyleAkongte, Peter Njukang, Bo-Sun Park, Dong-Won Kim, and Yong-Soo Choi. 2023. "Honey Bee Colonies (Apis mellifera L.) Perform Orientation Defensiveness That Varies among Bred Lines" Insects 14, no. 6: 546. https://doi.org/10.3390/insects14060546
APA StyleAkongte, P. N., Park, B.-S., Kim, D.-W., & Choi, Y.-S. (2023). Honey Bee Colonies (Apis mellifera L.) Perform Orientation Defensiveness That Varies among Bred Lines. Insects, 14(6), 546. https://doi.org/10.3390/insects14060546