
Multiple Aspects of the Fight against the Red Palm Weevil in an Urban Area: Study Case, San Benedetto del Tronto (Central Italy)




Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
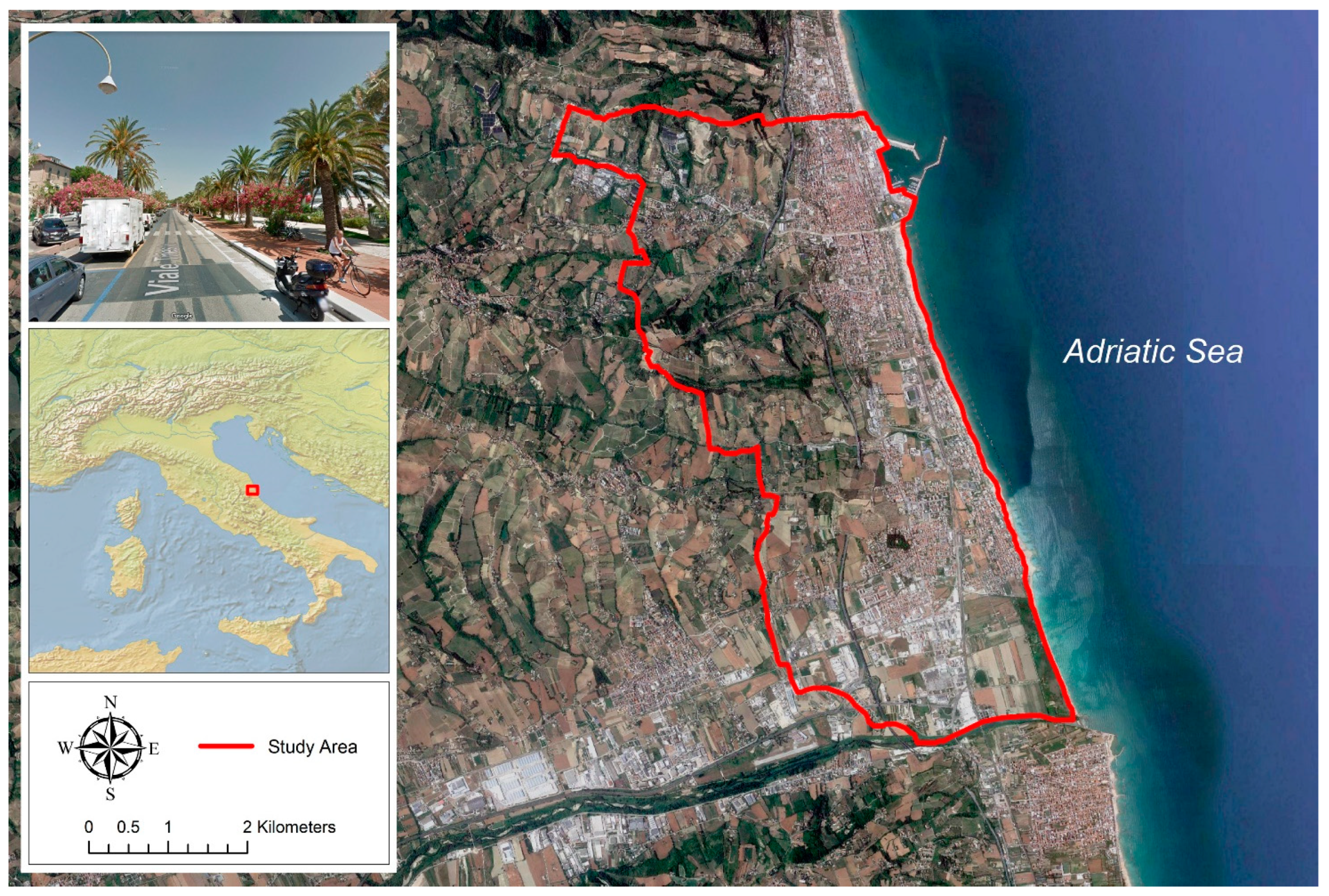
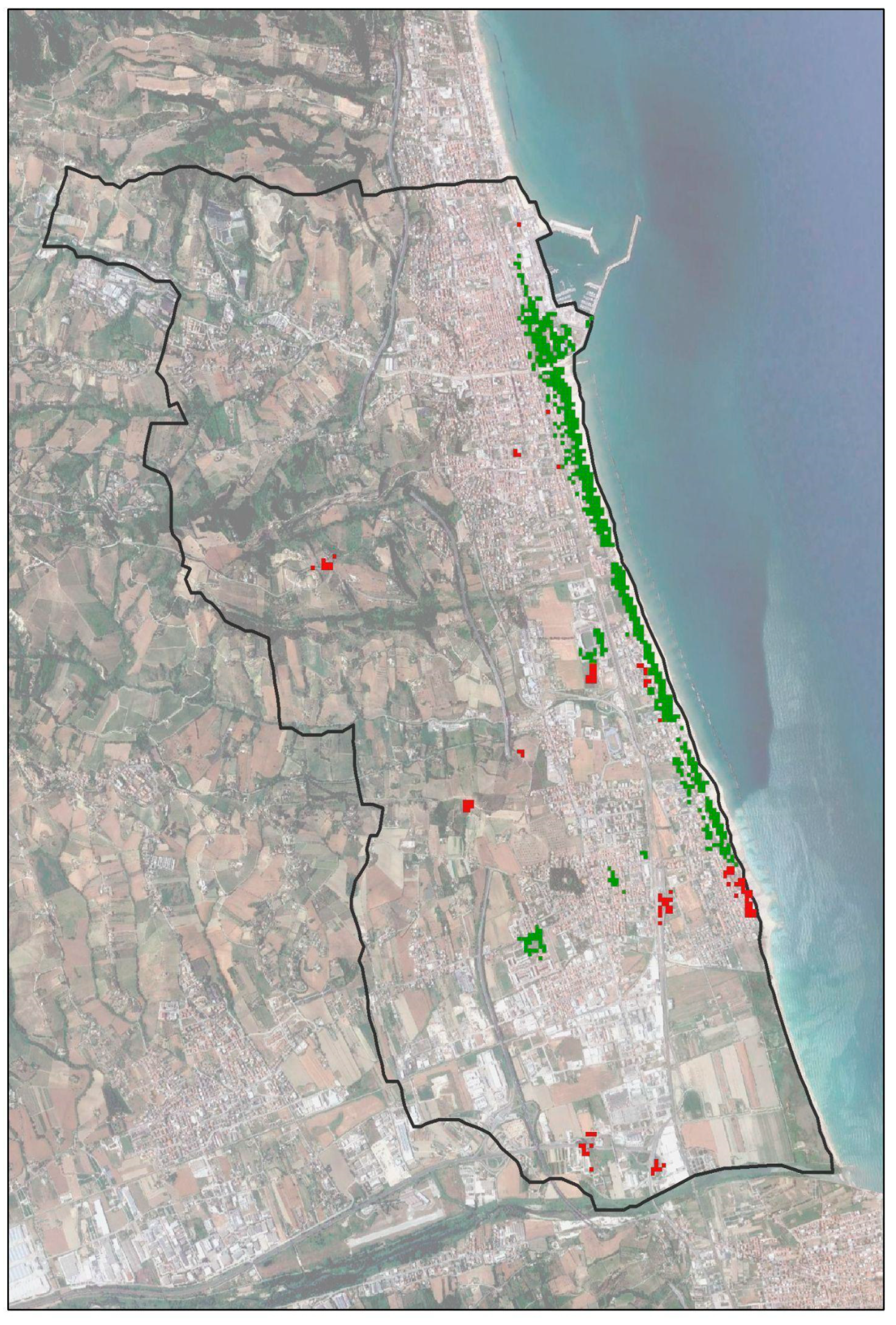
2.1. The Study Area and Collected Data
2.2. Geo-Statistical Analysis
2.3. In Silico Prediction of Insecticide Toxicity
3. Results and Discussion
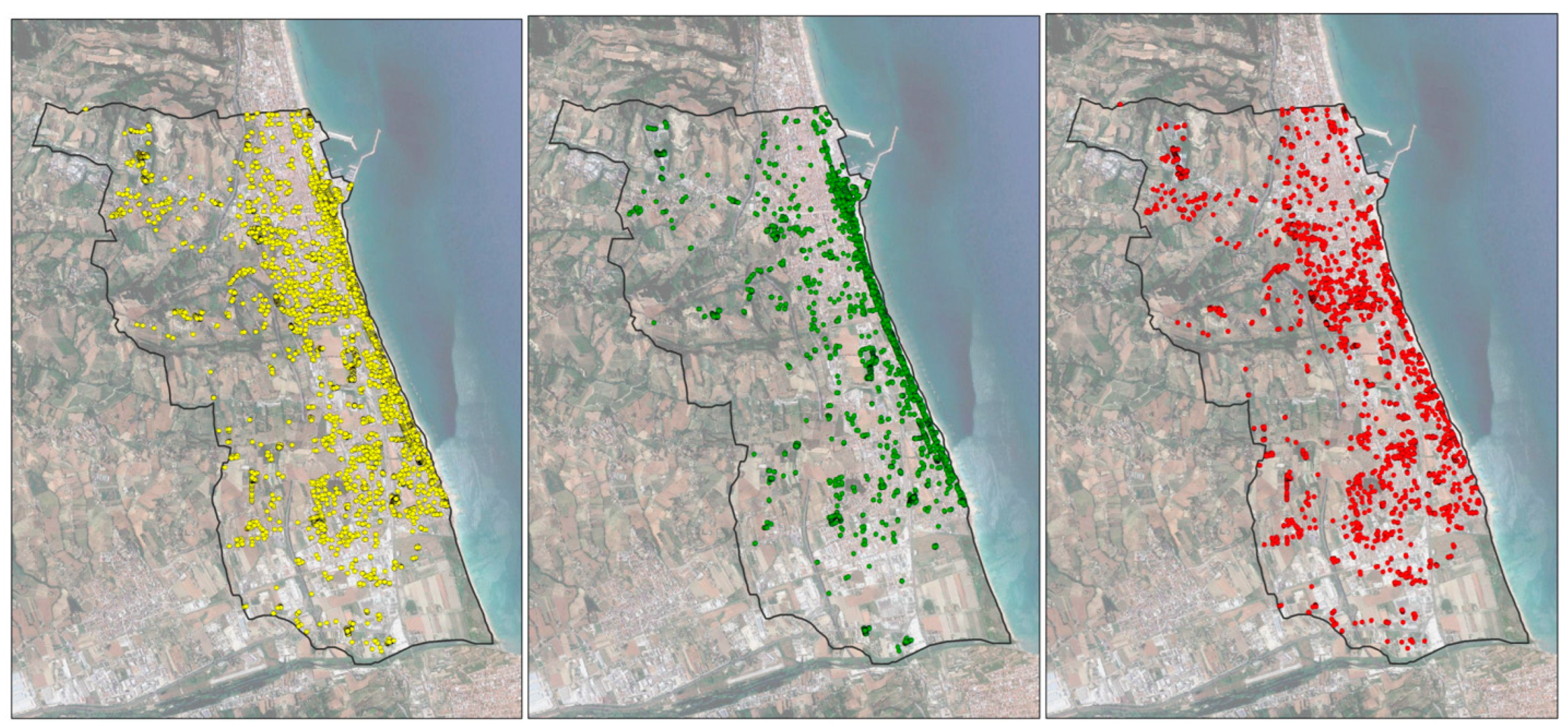
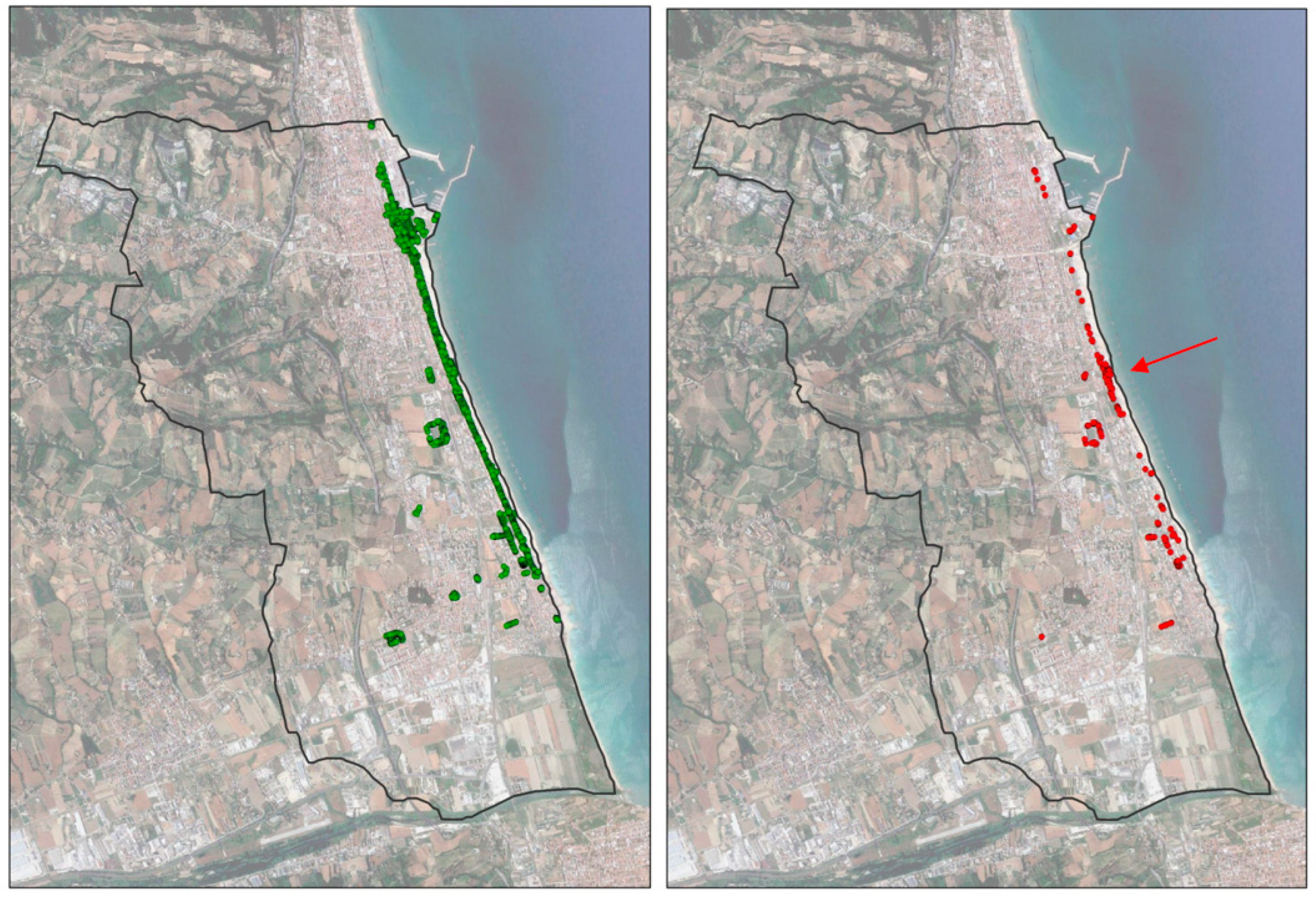
3.1. RPW Occurrence
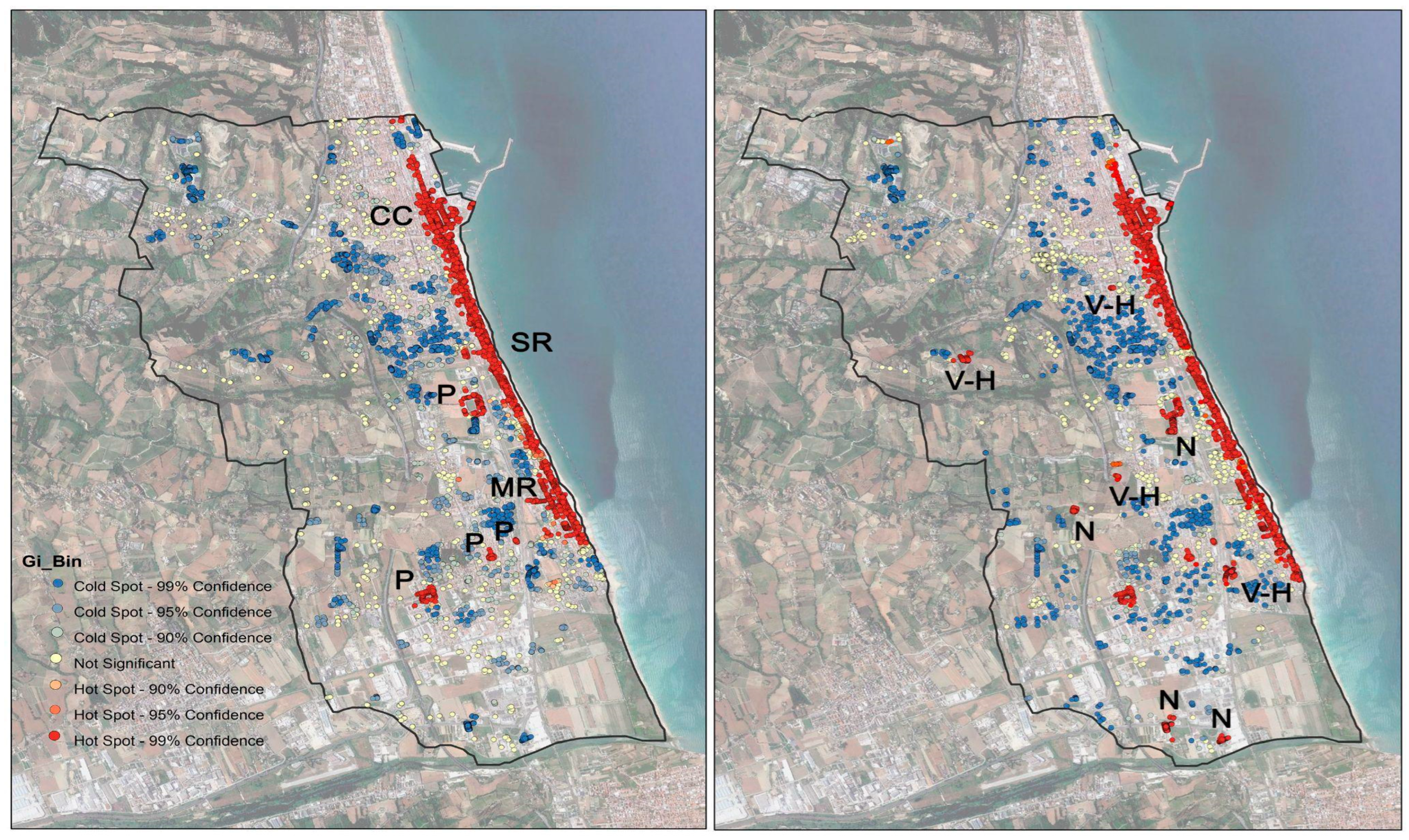
3.2. Geo-Statistical Analysis
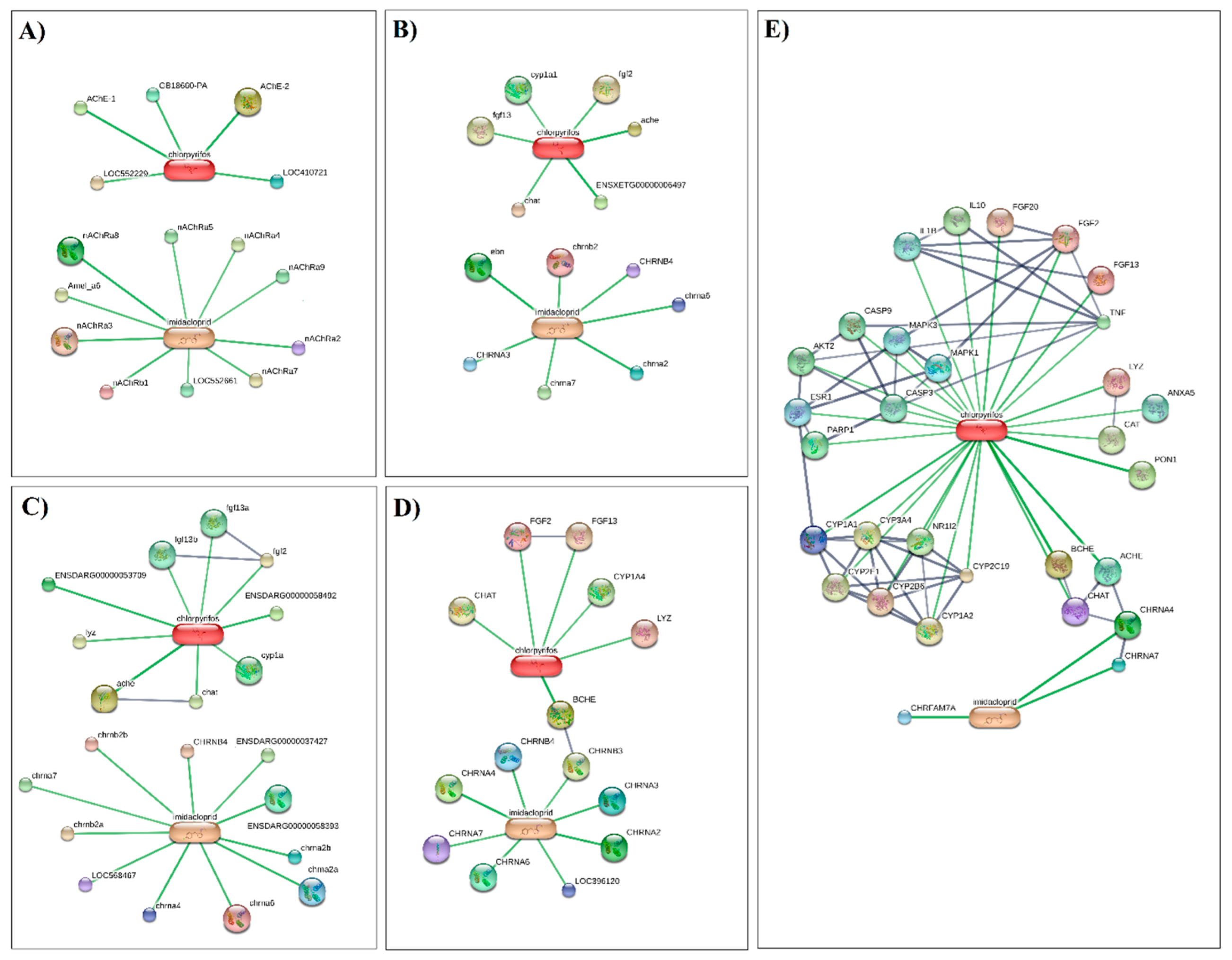
3.3. Computational Approach to Predict Imidacloprid- and Chlorpyrifos-Protein Interaction Networks
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cox, M.L. Red palm weevil, Rhynchophorus ferrugineus in Egypt. FAO Plant Prot. Bull. 1993, 41, 30–31. [Google Scholar]
- Milosavljević, I.; El-Shafie, H.A.F.; Faleiro, J.R.; Hoddle, C.D.; Leewis, M.; Hoddle, M.S. Palmageddon: The wasting of ornamental palms by invasive palm weevils, Rhynchophorus spp. J. Pest. Sci. 2019, 92, 143–156. [Google Scholar] [CrossRef]
- EPPO (European and Mediterranean Plant Prot. Org.). Global Database on Quarantine Pets. Available online: http://www.eppo.int (accessed on 10 September 2013).
- Kaakeh, W. Longevity, fecundity, and fertility of the red palm weevil, Rhynchophorus ferrugineus Olivier (Coleoptera: Curculionidae) on natural and artificial diets. Emir. J. Agric. Sci. 2015, 17, 23–33. [Google Scholar]
- Dembilio, Ó.; Jaques, J.A. Biology and management of Red palm weevil. In Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges; Wakil, W., Faleiro, J.R., Miller, T.A., Eds.; Springer: Basel, Switzerland, 2015; pp. 13–36. [Google Scholar]
- Faleiro, J.R. A review on the issues and management of red palm weevil Rhynchophorus ferrugineus (Coleoptera: Rhynchophoridae) in coconut and date palm during the last one hundred years. Int. J. Trop. Insect. Sci. 2006, 26, 135–154. [Google Scholar]
- Hagley, E.A.C. On the life history and habits of the palm weevil, Rhynchophorus palmarium. Ann. Entomol. Soc. Am. 1965, 58, 22–28. [Google Scholar] [CrossRef]
- Wattanpongsiri, A. A Revision of the Genera Rhynchophorus and Dynamis (Coleoptera: Curculionidae); Department of Agriculture Science Bulletin: Bangkok, Thailand, 1966; Volume 1, p. 328. [Google Scholar]
- Leefmans, S. De palmsnuitkever (Rhynchophorus ferrugineus Olivier). Meded. Inst. Plziekt. Buitenzorg 1920, 43, 1–90. [Google Scholar]
- Griffith, R. Red ring disease of coconut palm. Plant Dis. 1987, 71, 192–196. [Google Scholar]
- Weissling, T.J.; Giblin-Davis, R.M. Water loss dynamics and humidity preference of Rhynchophorus cruentatus (Coleoptera: Curculionidae) adults. Environ. Entomol. 1983, 22, 94–98. [Google Scholar] [CrossRef]
- Weissling, T.J.; Giblin-Davis, R.M.; Gries, G.; Gries, R.; Pérez, A.L.; Pierce, H.D.; Oehlschlager, A.C. Aggregation pheromone of palmetto weevil, Rhynchophorus cruentatus (F.) (Coleoptera: Curculionidae). J. Chem. Ecol. 1994, 20, 505–515. [Google Scholar] [CrossRef]
- Abbas, M.S.T.; Hanounik, S.B.; Shahdad, A.S.; Ai-Bagham, S.A. Aggregation pheromone traps, a major component of IPM strategy for the red palm weevil, Rhynchophorus ferrugineus in date palms (Coleoptera: Curculionidae). J. Pest. Sci. 2006, 79, 69–73. [Google Scholar] [CrossRef]
- Soroker, V.; Suma, P.; La Pergola, A.; Liopis, N.V.; Vacas, S.; Cohen, Y.; Alchanatis, V.; Milonas, P.; Golomb, O.; Goldshtein, E.; et al. Surveillance techniques and detection methods for Rhynchophorus ferrugineus and Paysandisia archon. In Handbook of Major Palm Pests; Biology and Management; John Wiley & Sons, Ltd.: Chichester, UK, 2017; Chapter 10; pp. 209–232. [Google Scholar]
- Wraight, S.P.; Jackson, M.A.; Kock, S.L. Production, Stabilization and Formulation of Fungal Biocontrol Agents. In Fungi as Biocontrol Agents: Progress, Problems and Potential; Butt, T.M., Jackson, C., Magan, N., Eds.; Centre for Agriculture and Bioscience International: Wallingford, UK, 2001; Chapter 10; pp. 253–287. [Google Scholar]
- Rochat, D.; Chapin, E.; Ferry, M.; Avand-Faghih, A.; Brun, L. Le charançon rouge du palmier dans le bassin méditerranéen. Phytoma-La Défense des Végetaux 2006, 595, 20–24. [Google Scholar]
- Faleiro, J.R.; Ben Abdallah, A.; El-Bellaj, M.; Al Ajlan, A.M.; Oihabi, A. Threat of the Red Palm Weevil, Rhynchophorus ferrugineus (Olivier) to Date Palm Plantations in North Africa. Arab. J. Plant Prot. 2012, 30, 274–280. [Google Scholar]
- Masilamany, D.; Wan Ali, W.K.A.; Baki, R. Biologi dan Pengurusan Kumbang Jalur Merah—Perosak Invasif Tanaman Kelapa (Biology and management of red palm weevil—An invasive pest of coconut palm). Bull. Teknol. MARDI 2012, 1, 151–157. [Google Scholar]
- Mazza, G.; Francardi, V.; Simoni, S.; Benvenuti, C.; Cervo, R.; Romeno, J.; Llácer, E.; Longo, S.; Nannelli, R.; Tarasco, E.; et al. An Overview on the Natural Enemies of Rhynchophorus Palm Weevils, with Focus on R. ferrugineus. Biol. Control 2014, 77, 83–92. [Google Scholar] [CrossRef]
- Giblin-Davis, R.M.; Faleiro, J.R.; Jacas, J.A.; Peña, J.E.; Vidyasagar, P.S.P.V. Biology and management of the red palm weevil, Rhynchophorus ferrugineus. In Potential Invasive Pests of Agricultural Crops; Peña, J.E., Ed.; Commonwealth Agricultural Bureau (CAB) International: London, UK, 2013; Chapter 1; pp. 1–34. [Google Scholar]
- Lin, G.L.E.; Salim, J.M.; Halim, M.F.A.; Azmi, W.A. Entomopathogenic fungi isolated from the soil of Terengganu, Malaysia as potential bio-pesticides against the red palm weevil, Rhynchophorus ferrugineus. J. Sustain. Sci. Manag. 2017, 12, 71–79. [Google Scholar]
- Masilamany, D.; Tang, B. Pengurusan Bersepadu Kumbang Badak dan Kumbang Jalur Merah—Perosak Utama Tanaman Kelapa (Integrated management of rhinoceros beetle and red palm weevil—Major pests of coconut palm). Bul. Teknol. MARDI 2013, 5, 51–59. [Google Scholar]
- Al-Shawaf, A.; Al-Shagag, A.; Al-Bagshi, M.; Al-Saroj, S.; AlBather, S.; Al-Dandan, A.M.; Faleiro, J.R. A quarantine protocol against red palm weevil Rhynchophorus ferrugineus (Olivier) (Coleptera: Curculiondae) in date palm. J. Plant Prot. Res. 2013, 53, 409–415. [Google Scholar] [CrossRef]
- Peri, E.; Colazza, S.; Guarino, S.; Suma, P.; Pergola, A.; Longo, S. The red palm weevil in Sicily: The introduction and spread of an invasive alien species. In Proceedings of the Palm Pest Mediterranean Conference (171–177), Nice, France, 16–18 January 2013. [Google Scholar]
- Al-Dosary, N.M.N.; Al-Dobai, S.; Faleiro, J.R. Review on the management of red palm weevil Rhynchophorus ferrugineus Olivier in date palm Phoenix dactylifera L. Emir. J. Food Agric. 2016, 28, 34–44. [Google Scholar] [CrossRef]
- Llopis, V.N.; González, S.V.; Jaques, J.A.; Moraga, E.Q.; Vives, Ó.D.; Moya, P. Desarrollo de un sistema de atracción-infección para el control del picudo rojo de la palmera (Rhynchophorus ferrugineus) mediante hongos entomopatógenos. Phytoma España 2018, 298, 36–39. [Google Scholar]
- DOA (Department of Agriculture). Report on Current Status of Attack of the Red Palm Weevil, Rhynchophorus ferrugineus in Terengganu, Malaysia; Government Press: Kuala Lumpur, Malaysia, 2016.
- Kielbaso, J.J.; Davidson, H.; Hart, J.; Jones, A.; Kennedy, M.K. Characteristics of successful systemic chemicals. In Proceedings of Symposium on Systemic Chemical Treatment in Tree Culture; Kielbaso, J.J., Ed.; Michigan State University: East Lansing, MI, USA, 1979; pp. 19–34. [Google Scholar]
- Dembilio, Ó.; Riba, J.M.; Gamón, M.; Jacas, J.A. Mobility and efficacy of abamectin and imidacloprid against Rhynchophorus ferrugineus in Phoenix canariensis by different application methods. Pest Manag. Sci. 2014, 71, 1091–1098. [Google Scholar] [CrossRef]
- Ferry, M.; Gomez, S. Red palm weevil, focus on it control. Phytoma 2012, 658, 38–41. [Google Scholar]
- Chihaoui-Meridja, S.; Harbi, A.; Abbes, K.; Chaabane, H.; La Pergola, A.; Chermiti, B.; Suma, P. Systematicity, persistence and efficacy of selected insecticides used in endotherapy to control the red palm weevil Rhynchophorus ferrugineus (Olivier, 1790) on Phoenix canariensis. Phytoparasitica 2020, 48, 75–85. [Google Scholar] [CrossRef]
- Nardi, S.; Ricci, E.; Lozzi, R.; Marozzi, F.; Ladurner, E.; Chiabrando, F.; Granchelli, L.; Verdolini, E.; Isidoro, N.; Riolo, P. Control of Rhynchophorus ferrugineus (Olivier, 1790) according to EU Decision 2007/365/EC in the Marche region (Central-Eastern Italy). OEPP/EPPO Bull. 2011, 41, 103–115. [Google Scholar] [CrossRef]
- EPA Toxicology Chapter for Chlorpyrifos; Environmental Protections Agency; Office of Prevention, Pesticides and Toxic Substances: Washington, DC, USA, 2000. Available online: http://www.epa.gov/oppsrrd1/op/chlorpyrifos/rev_tox.pdf (accessed on 18 April 2000).
- Watts, M. Chlorpyrifos as a Possible Global POP; Pesticide Action Network North America: Oakland, CA, USA, 2012; Available online: https://www.rapaluruguay.org/sitio_1/agrotoxicos/Chlorpyrifos_as_POP.pdf (accessed on 20 June 2019).
- Ubaid, U.; Rahman, H.; Asghar, W.; Nazir, W.; Sandhu, M.A.; Ahmed, A.; Khalid, N. A comprehensive review on chlorpyrifos toxicity with special reference to endocrine disruption: Evidence of mechanisms, exposures and mitigation strategies. Sci. Total Environ. 2021, 755 Pt 2, 142649. [Google Scholar] [CrossRef]
- Gervais, J.A.; Luukinen, B.; Buhl, K.; Stone, D. Imidacloprid Technical Fact Sheet; National Pesticide Information Center: Corvallis, OR, USA, 2010. [Google Scholar]
- Stoner, K.A.; Eitzer, B.D. Movement of soil-applied imidacloprid and thiamethoxam into nectar and pollen of squash (Cucurbita pepo). PLoS ONE 2012, 7, 39114. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.D.; Pettis, J.S. A survey of imidacloprid levels in water sources potentially frequented by honeybees (Apis mellifera) in the Eastern USA. Water Air Soil Pollut. 2014, 225, 2127. [Google Scholar] [CrossRef]
- Chagnon, M.; Kreutzweiser, D.; Mitchell, E.A.D.; Morrissey, C.A.; Noome, D.A.; Van der Sluijs, J.P. Risks of large-scale use of systemic insecticides to ecosystem functioning and services. Environ. Sci. Pollut. Res. 2015, 22, 119–134. [Google Scholar] [CrossRef]
- Rogers, K.H.; McMillin, S.; Olstad, K.J.; Poppenga, R.H. Imidacloprid Poisoning of Songbirds Following a Drench Application of Trees in a Residential Neighborhood in California, USA. Environ. Toxicol. Chem. 2019, 38, 1724–1727. [Google Scholar] [CrossRef]
- Cinnirella, A.; Bisci, C.; Nardi, S.; Ricci, E.; Palermo, A.P.; Bracchetti, L. Analysis of the spread of Rhynchophorus ferrugineus in an urban area, using GIS techniques: A study case in Central Italy. Urban Ecosyst. 2020, 23, 255–269. [Google Scholar] [CrossRef]
- Cohen, Y.; Alchanatis, V.; Prigojin, A.; Levi, A.; Soroker, V.; Cohen, Y. Use of aerial thermal imaging to estimate water status of palm trees. Precis. Agric. 2012, 13, 123–140. [Google Scholar] [CrossRef]
- Hogan, S.; Kelly, M.; Stark, B.; Chen, Y. Unmanned aerial systems for agriculture and natural resources. Calif. Agric. 2017, 71, 5–14. [Google Scholar] [CrossRef]
- Pesaresi, S.; Biondi, E.; Casavecchia, S. Bioclimates of Italy. J. Maps 2017, 13, 955–960. [Google Scholar] [CrossRef]
- Fazzini, M.; Beltrando, G.; Billi, P. Intense rainfalls and flooding problems in the beach resort of San Benedetto del Tronto, Adriatic Sea, Central Italy. In Proceedings of the 3°GEOMED 2013, Antalya, Turkey, 10–14 June 2013; pp. 128–131. [Google Scholar]
- Municipality of di San Benedetto del Tronto, Search for Administrative Actions Service. Available online: https://www.comunesbt.it/L-Amministrazione/Ricerca-Atti-Amministrativi (accessed on 16 June 2020).
- ESRI. ArcGIS 10.X. Redlands: Environmental Systems Research Institute ESRI. GIS Mapping Software, Solutions, Services, Map Apps, and Data 2012. Available online: http://www.esri.com/ (accessed on 12 May 2021).
- Whitlock, M.; Schluter, D. The Analysis of Biological Data; W.H. Freeman and Company: New York, NY, USA, 2015; pp. 242–245. [Google Scholar]
- Roscoe, J.T.; Byars, J.A. Sample size restraints commonly imposed on the use of the chi-square statistic. J. Am. Stat. Assoc. 1971, 66, 755–759. [Google Scholar] [CrossRef]
- Iskar, M.; Campillos, M.; Kuhn, M.; Jensen, L.J.; van Noort, V.; Bork, P. Drug-Induced Regulation of Target Expression. PLoS Comput. Biol. 2010, 6, e1000925. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S. In silico analysis identifies genes common between five primary gastrointestinal cancer sites with potential clinical applications. Ann. Gastroenterol. 2014, 27, 231–236. [Google Scholar]
- Smart, T.G.; Paoletti, P. Synaptic neurotransmitter-gated receptors. Cold Spring Harb. Perspect. Biol. 2012, 4, a009662. [Google Scholar] [CrossRef]
- Shimada, S.; Kamiya, M.; Shigetou, S.; Tomiyama, K.; Komori, Y.; Magara, L.; Makoto, I.; Matsuda, K. The mechanism of loop C-neonicotinoid interactions at insect nicotinic acetylcholine receptor α1 subunit predicts resistance emergence in pests. Sci. Rep. 2020, 10, 7529. [Google Scholar] [CrossRef]
- Ihara, M. Ligand-gated ion channels as targets of neuroactive insecticides. Biosci. Biotechnol. Biochem. 2022, 86, 157–164. [Google Scholar] [CrossRef]
- Burke, A.P.; Niibori, Y.; Terayama, H.; Ito, M.; Pidgeon, A.; Arsenault, J.; Camarero, P.R.; Cummins, C.L.; Mateo, R.; Sakabe, K.; et al. Mammalian susceptibility to a neonicotinoid insecticide after fetal and early postnatal exposure. Sci. Rep. 2018, 8, 16639. [Google Scholar] [CrossRef]
- Berheim, E.H.; Jenks, J.A.; Lundgren, J.G.; Michel, E.S.; Grove, D.; Jensen, W.F. Effects of neonicotinoid insecticides on physiology and reproductive characteristics of captive female and fawn white-tailed deer. Sci. Rep. 2019, 9, 4534. [Google Scholar] [CrossRef]
- Kimura-Kuroda, J.; Komuta, Y.; Kuroda, Y.; Hayashi, M.; Kawano, H. Nicotine-Like Effects of the Neonicotinoid Insecticides Acetamiprid and Imidacloprid on Cerebellar Neurons from Neonatal Rats. PLoS ONE 2012, 7, e32432. [Google Scholar] [CrossRef] [PubMed]
- Loser, D.; Hinojosa, M.G.; Blum, J.; Schaefer, J.; Brüll, M.; Johansson, Y.; Suciu, I.; Grillberger, K.; Danker, T.; Möller, C.; et al. Functional alterations by a subgroup of neonicotinoid pesticides in human dopaminergic neurons. Arch. Toxicol. 2021, 95, 2081–2107. [Google Scholar] [CrossRef] [PubMed]
- Berg, E.L.; Ching, T.M.; Bruun, D.A.; Rivera, J.K.; Careaga, M.; Ellegood, J.; Lerch, J.P.; Wöhr, M.; Lein, P.J.; Silverman, J.L. Translational outcomes relevant to neurodevelopmental disorders following early life exposure of rats to chlorpyrifos. J. Neurodevelop. Disord. 2020, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, T.A.; Seidler, F.J. The alterations in CNS serotonergic mechanisms caused by neonatal chlorpyrifos exposure are permanent. Braines Dev. Brain Res. 2005, 158, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Beasley, V.R. Direct and Indirect Effects of Environmental Contaminants on Amphibians. In Refrence Module in Earth Systems and Environmental Sciences; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Hodgson, E. Biotransformation of Individual Pesticides: Some Examples. In Pesticide Biotransformation and Disposition, Third edition; Elsevier: Oxford, UK, 2012; pp. 195–208. [Google Scholar]
- Tanvir, E.M.; Afroz, R.; Chowdhury, M.; Gan, S.H.; Karim, N.; Islam, M.N.; Khalil, M.I. A model of chlorpyrifos distribution and its biochemical effects on the liver and kidneys of rats. Hum. Exp. Toxicol. 2016, 35, 991–1004. [Google Scholar] [CrossRef]
- Abolaji, A.O.; Ojo, M.; Afolabi, T.T.; Arowoogun, M.D.; Nwawolor, D.; Farombi, E.O. Protective properties of 6-gingerol-rich fraction from Zingiber officinale (Ginger) on chlorpyrifos-induced oxidative damage and inflammation in the brain, ovary and uterus of rats. Chem. Biol. Interact. 2017, 25, 15–23. [Google Scholar] [CrossRef]
- Sawyer, A.J.; Casagrande, R.A. Urban pest management: A conceptual framework. Urban Ecol. 1983, 7, 145–157. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Number |
|---|---|
| Total palms | 7385 |
| Healthy | 3442 |
| Dead-infested | 3943 (of which 2456 have died since 2013) |
| Treated (since 2013) | 1627 |
| Healthy and treated | 1460 |
| Dead-infested and treated | 167 |
| Untreated | 5758 |
| Healthy and untreated | 1982 (of which 1032 were in nurseries or villas/hotels) |
| Dead-infested and untreated | 3776 (of which 272 were in nurseries or villas/hotels) |
| Chemicals | Year |
|---|---|
| Afidina quick (clorpirifos, deltamethrin) | 2013–2014 |
| Khoinor plus (imidacloprid, ciflutrin) | 2015 |
| Reldan (clorpirifos-metile) | 2016–2019 |
| Asset five (pyrethrins) | 2020 |
| Intercept granular (imidacloprid) | 2013–2020 |
| Pathway ID | Pathway Description | #Protein | Matching Proteins in the Network |
|---|---|---|---|
| Apis mellifera | |||
| IPR006201 IPR006202 IPR018000 IPR006029 AME04040 | Neurotransmitter-gated ion channel family | 8 | nAChRa2, nAChRa3, nAChRa4, nAChRa5, nAChRa6, nAChRa7, nAChRa8, nAChRb1 |
| GO:0042135 GO:0001507 | Neurotransmitter catabolic process | 3 | AChE-1, AChE-2, LOC410721 |
| AME00001 | Metabolism | 2 | GB18660-PA, LOC552229 |
| Xenopus silurana | |||
| PF02931 PF02932 | Neurotransmitter-gated ion channel family | 8 | Chrna3, chrnb4, ENSXETG00000018003, chrna2, chrna6, chrna7, chrnb2, ebn |
| GO:0006581 | Neurotransmitter catabolic process | 1 | AChE |
| XTR00199 XTR00001 | Metabolism | 2 | CYP1A1, chat |
| XTR04052 | Signalling and cellular processes | 2 | fgf2, fgf13 |
| Danio rerio | |||
| PF02931 PF02932 | Neurotransmitter-gated ion channel family | 11 | Chrnab4, ENSDARG00000037427, ENSDARG00000058393, LOC568467, chrna2a, chrna2b, chrna4, chrna6, chrna7, chrnb2a, chrnb2b |
| GO:0042135 | Neurotransmitter catabolic process | 3 | AChE, ENSDARG00000053709, ENSDARG00000058492 |
| DRE00199 DRE00001 | Metabolism | 2 | CYP1A1, chat |
| DRE04052 DRE03037 | Signalling and cellular processes | 3 | fgf2, fgf13a, fgf13b, lyz |
| Gallus gallus | |||
| PF02931 PF02932 GGA04040 | Neurotransmitter-gated ion channel family | 8 | Chrna6, chrna7, chrnb3, LOC396120, chrna3, chrna4, chrnb4, chrna2 |
| GGA04147 | Signalling and cellular processes | 3 | LYZ, fgf2, fgf13 |
| GGA01000 GGA00199 | Metabolism | 3 | BCHE, CYP1A4, chat |
| Homo sapiens | |||
| GO:0005230 | Neurotransmitter-gated ion channel family | 3 | Chrna4, Chrna7, Chrfam7A |
| GO:0042136 | Neurotransmitter catabolic process | 1 | AChE |
| GO.0017144 GO:0006805 GO:0006629 GO:0002682 GO:0044255 GO:1901575 | Metabolism | 11 | BCHE, CYP1A1, CYP1A2, CYP2B6, CYP2C19, CYP2E1, CYP3A4, NR1I2, chat, CAT, PON1 |
| GO:0032870 GO:0008286 GO:0009967 GO:0060397 GO:0060396 HSA04010 IPR008996 | Signalling and cellular processes | 16 | AKT2, CASP9, CASP3, ESR1, FGF2, FGF20, IL10, FGF13, TNF, MAPK1, MAPK3, NR1I2, PARP1, LYZ, ANXA5, IL1B |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bracchetti, L.; Cocci, P.; Palermo, F.A. Multiple Aspects of the Fight against the Red Palm Weevil in an Urban Area: Study Case, San Benedetto del Tronto (Central Italy). Insects 2023, 14, 502. https://doi.org/10.3390/insects14060502
Bracchetti L, Cocci P, Palermo FA. Multiple Aspects of the Fight against the Red Palm Weevil in an Urban Area: Study Case, San Benedetto del Tronto (Central Italy). Insects. 2023; 14(6):502. https://doi.org/10.3390/insects14060502
Chicago/Turabian StyleBracchetti, Luca, Paolo Cocci, and Francesco Alessandro Palermo. 2023. "Multiple Aspects of the Fight against the Red Palm Weevil in an Urban Area: Study Case, San Benedetto del Tronto (Central Italy)" Insects 14, no. 6: 502. https://doi.org/10.3390/insects14060502
APA StyleBracchetti, L., Cocci, P., & Palermo, F. A. (2023). Multiple Aspects of the Fight against the Red Palm Weevil in an Urban Area: Study Case, San Benedetto del Tronto (Central Italy). Insects, 14(6), 502. https://doi.org/10.3390/insects14060502