
The Genus Heterogynis Rambur, 1866 (Heterogynidae, Lepidoptera): Congruence of Molecular, Morphological and Morphometric Evidence Reveal New Species in Serbia
 , , , , and
, , , , and
Simple Summary
Abstract
1. Introduction
2. Material and Methods
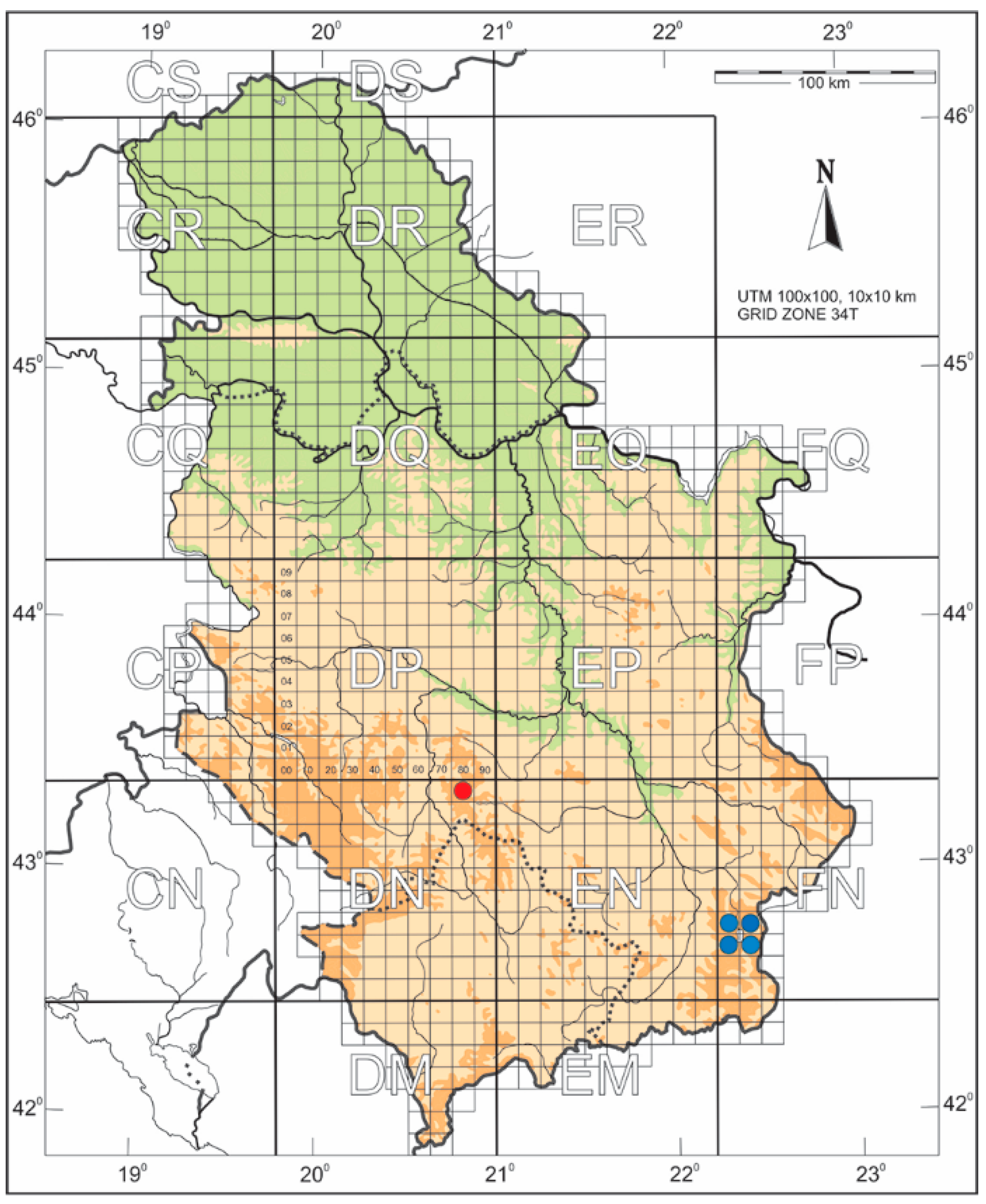
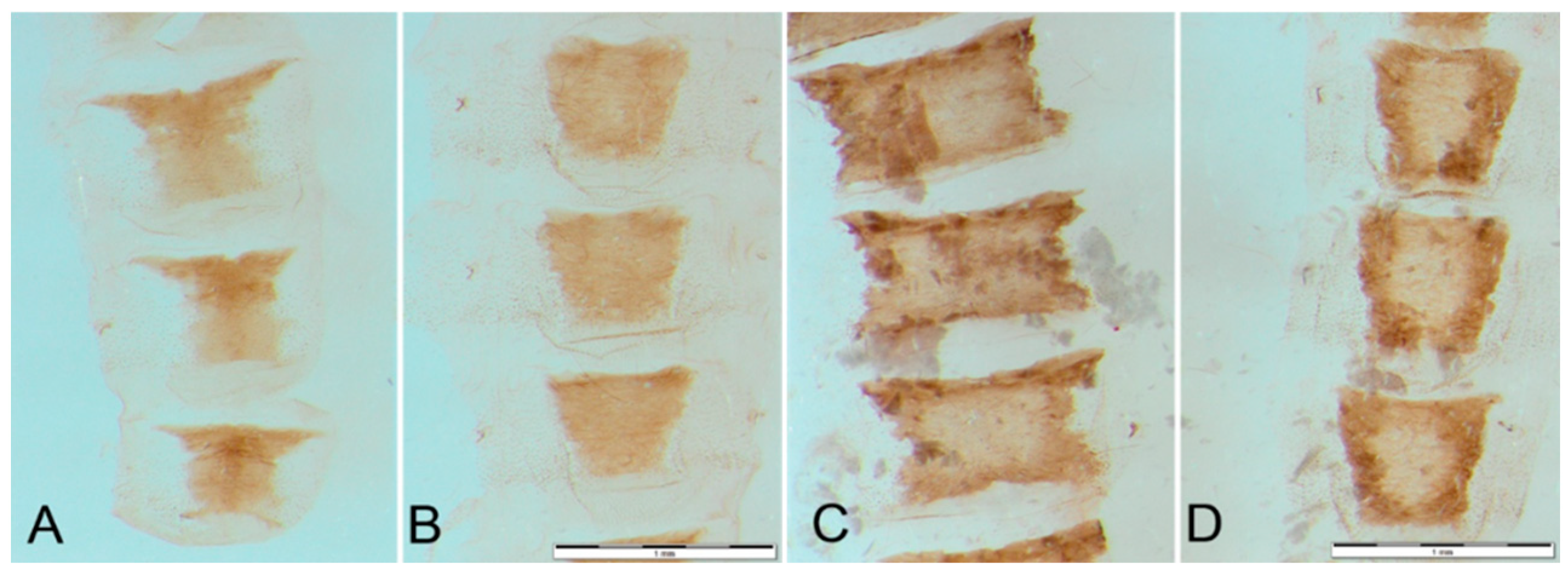
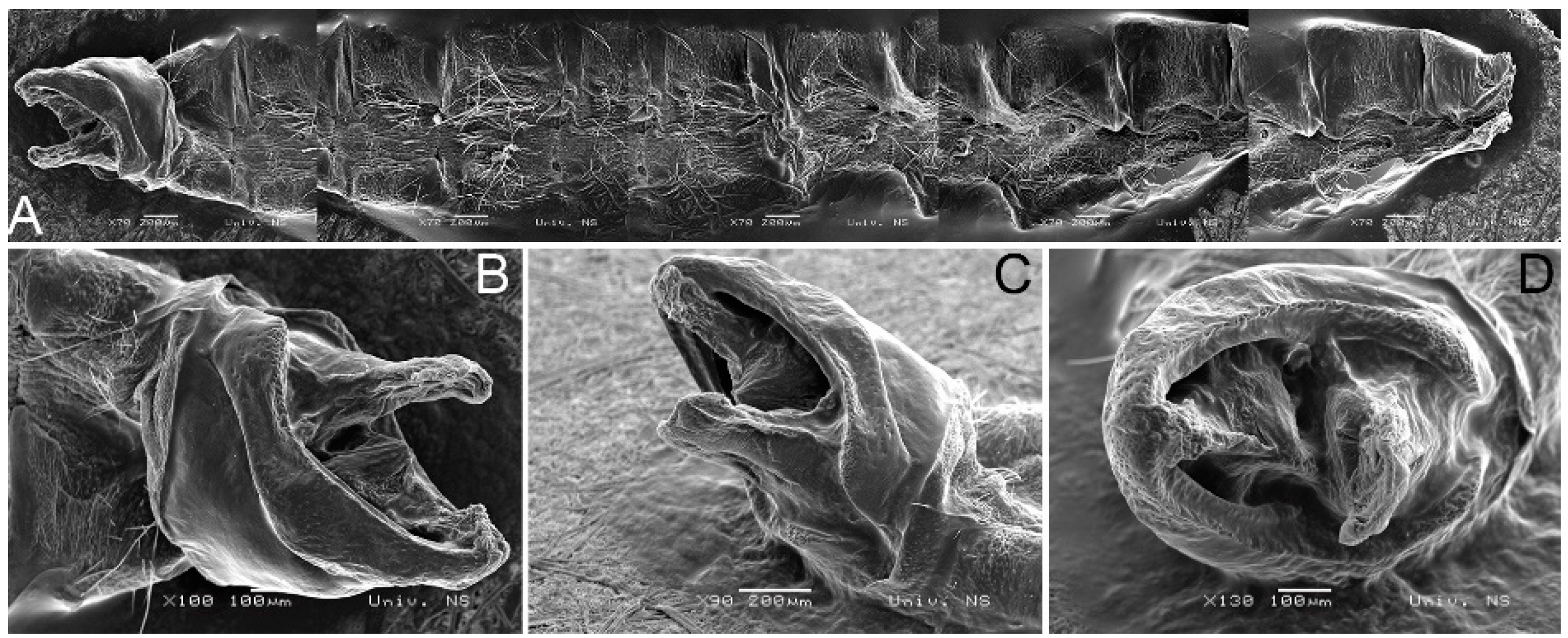
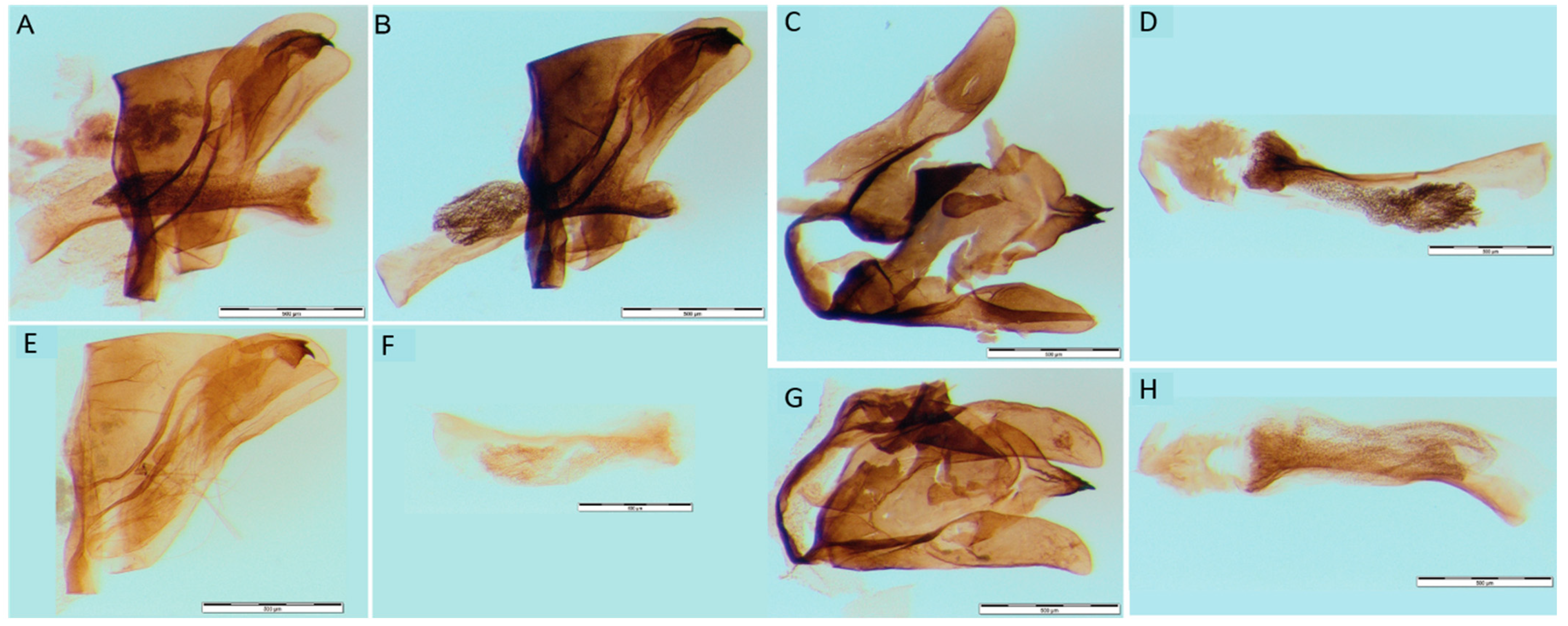
2.1. Collection and Morphological Analysis
2.2. Morphometric Analysis
2.3. DNA Isolation, Amplification and Sequencing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers for COI Genes | Primer Sequences | Species | Reference |
|---|---|---|---|
| LepF1 | 5′-ATTCAACCAATCATAAAGATATTG-3′ | H. affinis | [3] |
| LepR1 | 5′-TAAACTTCTGGATGTCCAAAAAATC-3′ | H. cynetis | |
| LEP-F1 | 5′-ATTCAACCAATCATAAAGATAT-3′; | Astraptes fulgerator | [25] |
| LEP-R1 | 5′-TAAACTTCTGGATGTCCAAAAA-3′ | ||
| LEP-R2 | 5′-CTTATATTATTTATTCGTGGGAAAGC-3′ | ||
| LCO1490 | 5′-GGTCAACAAATCATAAAGATATTGG-3′ | Diverse metazoan invertebrate | [29] |
| HCO2198 | 5′-TAAACTTCAGGGTGACCAAAAAATCA-3′ | ||
| LCO1490 | 5′-GGTCAACAAATCATAAAGATATTGG-3′ | Taxon barcode | [14,29] |
| HCO2198 | 5′-TAAACTTCAGGGTGACCAAAAAATCA-3′ | ||
| CO1-exF | 5′-ATCGCCTAAACTTCAGCCATT-3′ | Cucullia umbratica | [30,31] |
| TL-N-3017 | 5′-CTTAAATCCATTGCACTAATCTGCCATA-3′ | ||
| “Jerry” | 5′-CAACATTTATTTTGATTTTTTGG-3′ | Crematogaster fraxatrix | [27,28] |
| “Pat” | 5′-TCCAATGCACTAATCTGCCATATTA- 3′ |
2.4. Nucleotide Sequence Analyses
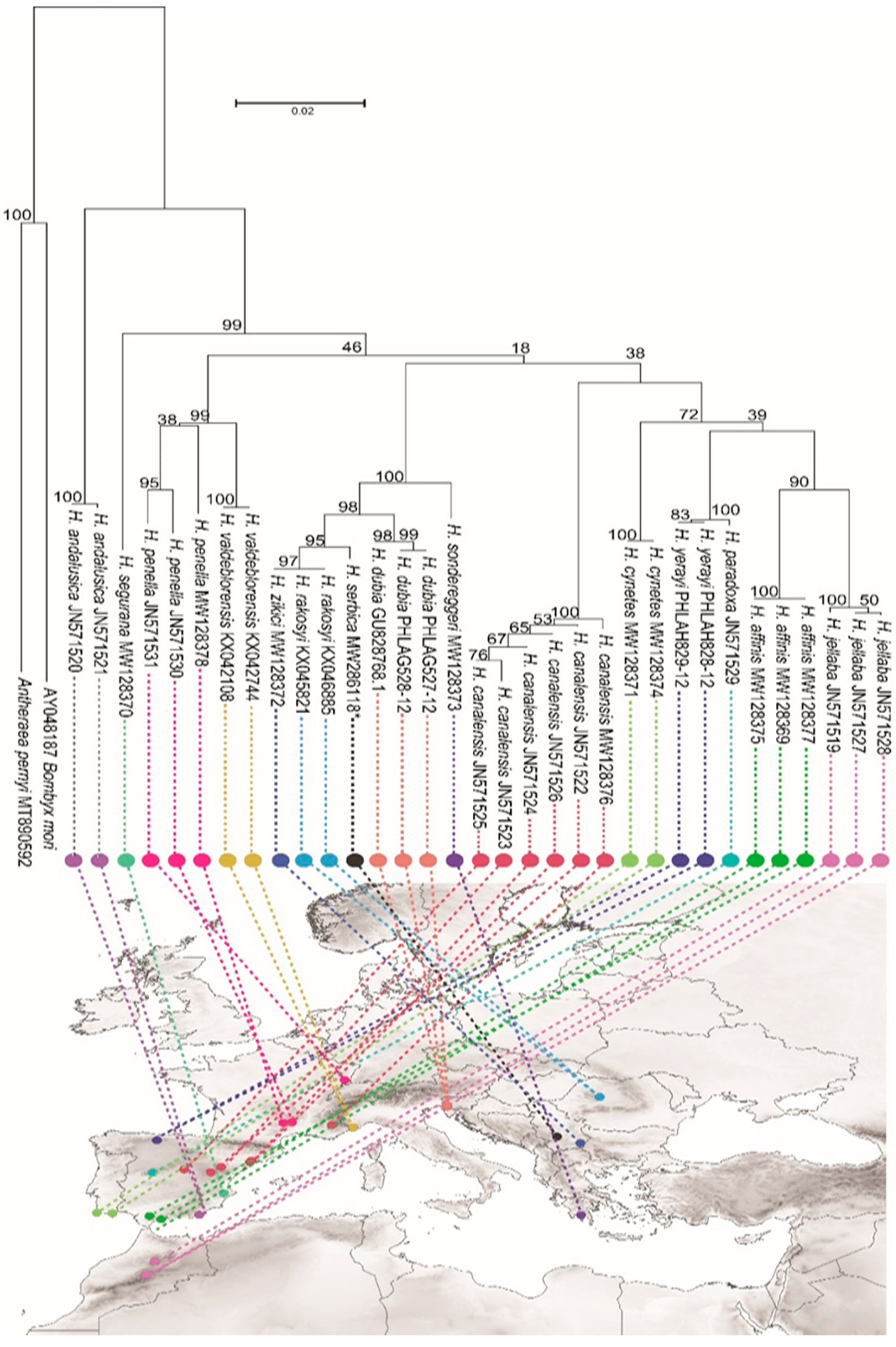
3. Results
3.1. Taxonomy
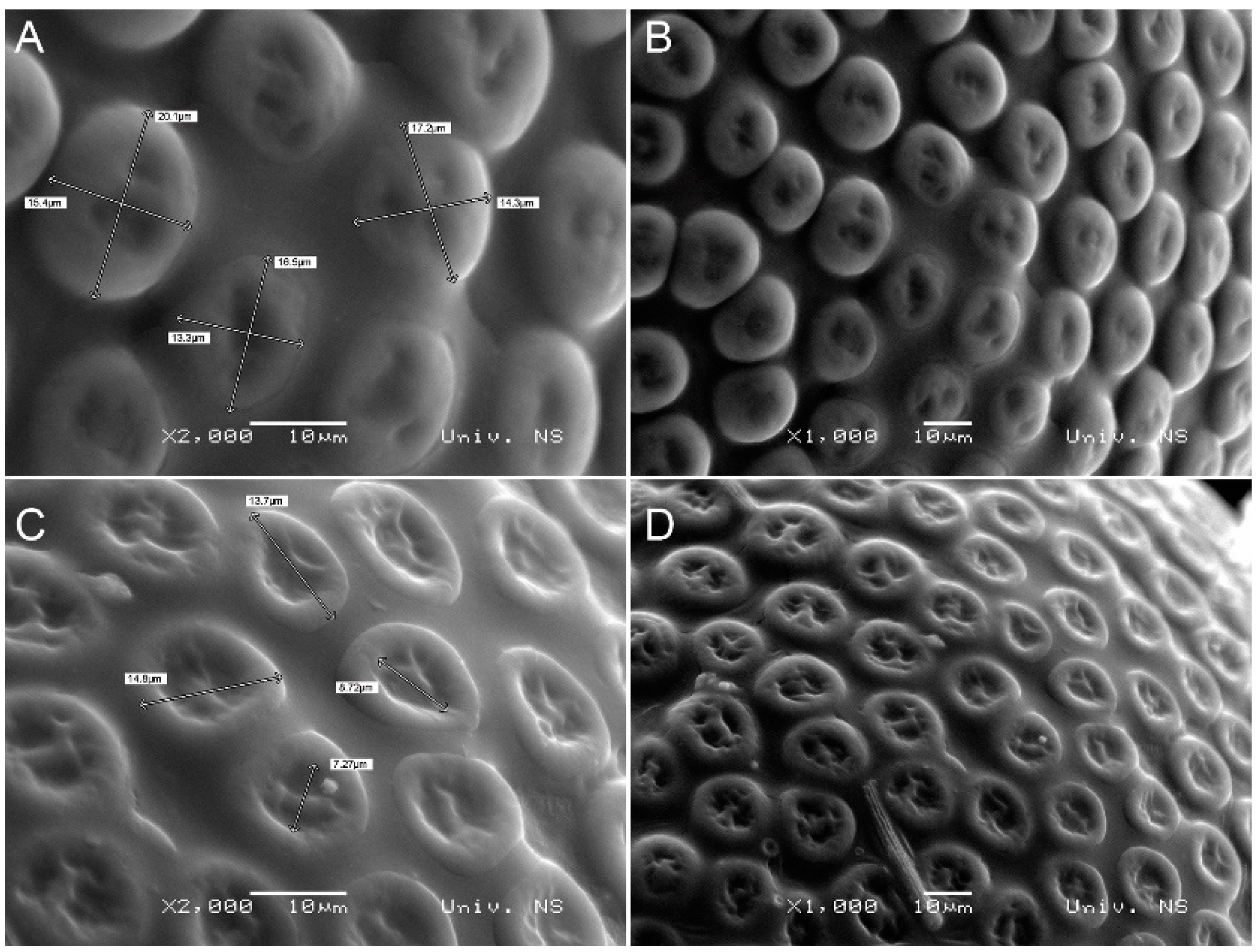
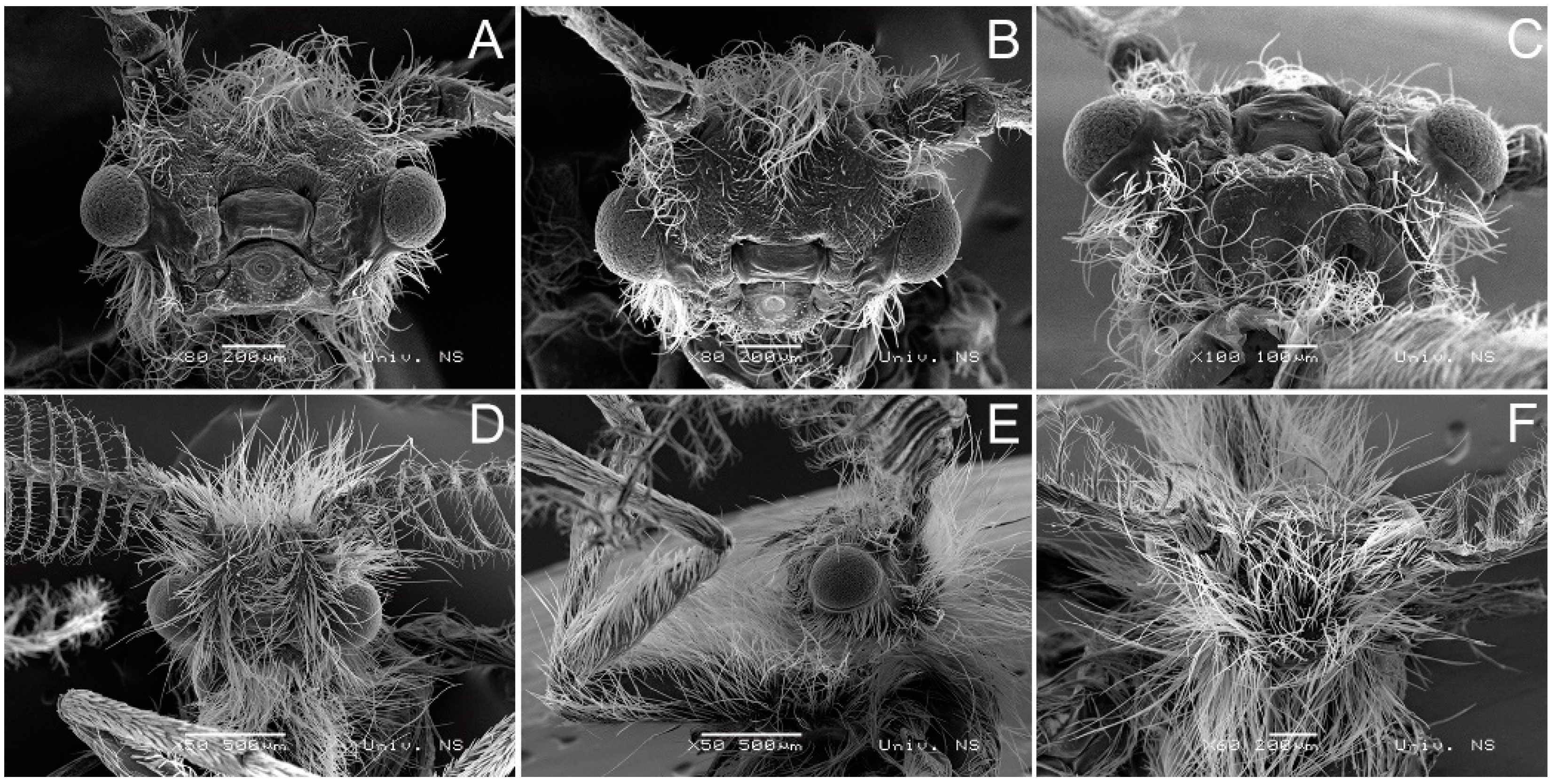
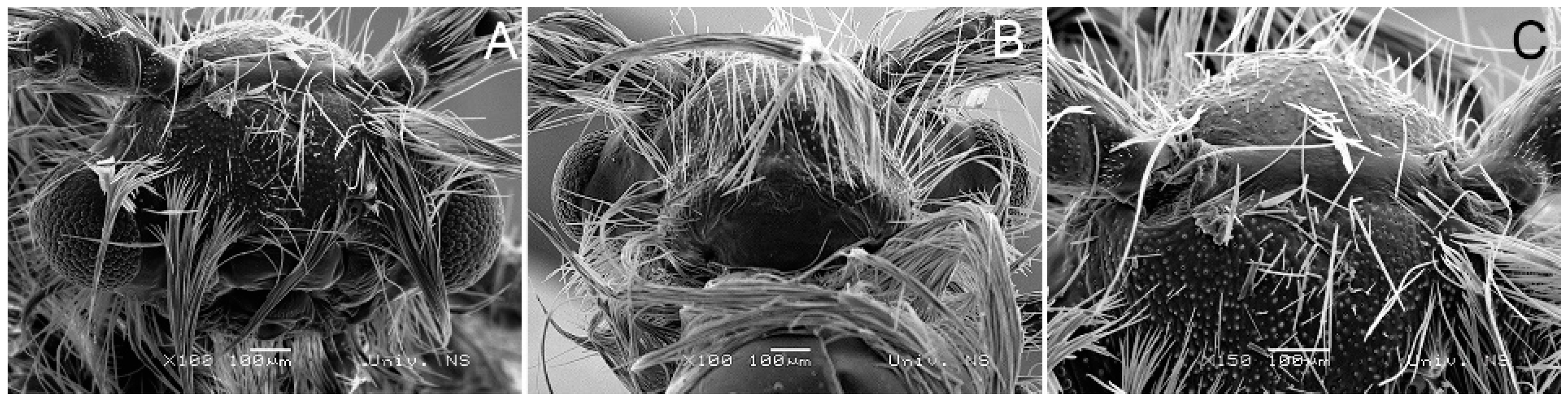
3.2. Morphological Diagnosis
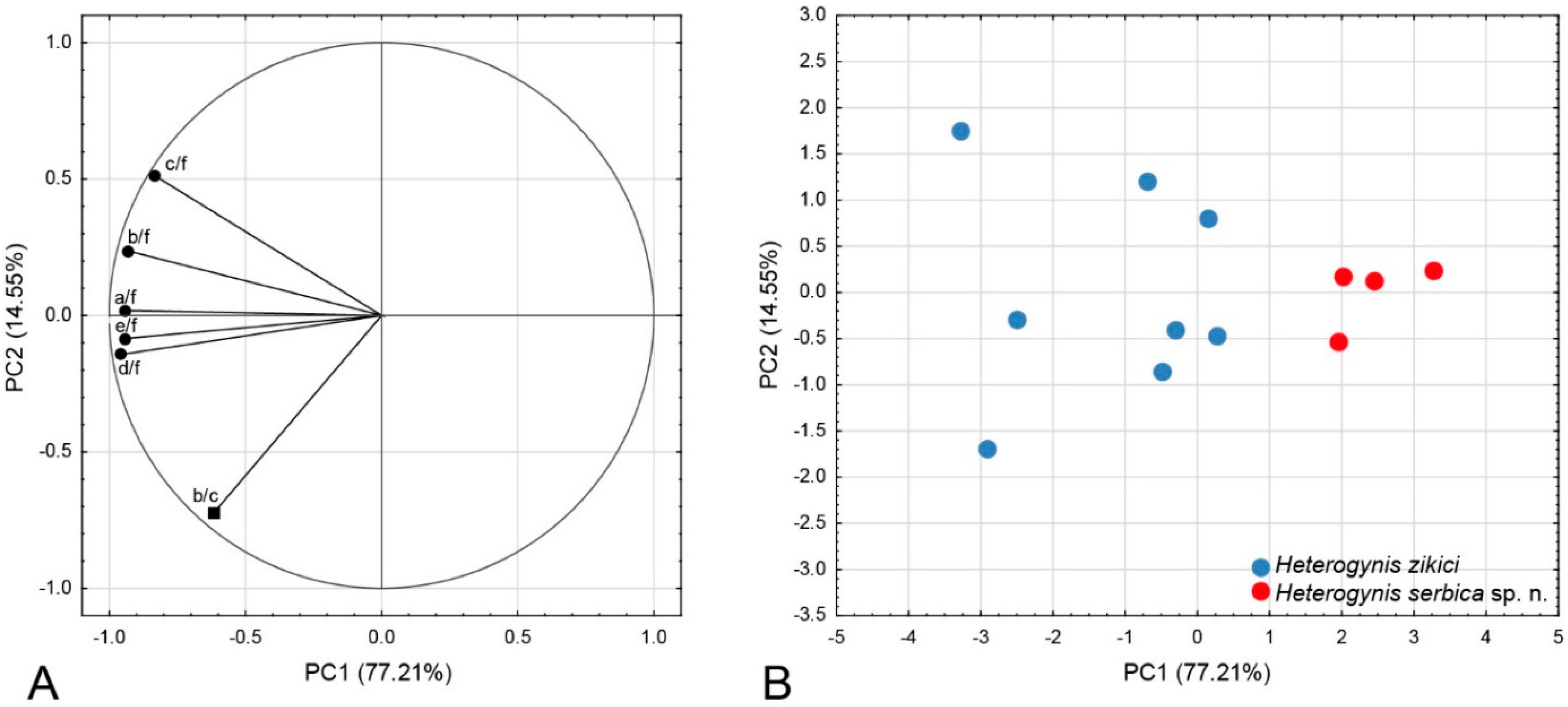
3.3. Morphometric Analysis
3.4. Molecular Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leraut, P. Moths of Europe, 1st ed.; N.A.P. Editions: Verrieres le Buisson, France, 2006; pp. pp. 124–128, 287. [Google Scholar]
- De Freina, J.J. Distribution and taxonomy of the complex of Heterogynis paradoxa Rambur, 1837, with description of Heterogynis yerayi sp. nov. from the Cantabrian Mountains (Lepidoptera: Zygaenoidea, Heterogynidae. NEVA 2011, 32, 9–24. [Google Scholar]
- De Freina, J.; Monasterio-Léon, Y.; Escobés-Jiménez, R.; Hinojosa, J.C.; Vila, R. Heterogynis Rambur, 1837 (Lepidoptera: Zygaenoidea, Heterogynidae) from Andalusia and the Algarve (Spain and Portugal): A taxonomic and faunistic review. Boletín SEA 2020, 67, 75–85. [Google Scholar]
- De Freina, J.J. A new subgenus of Heterogynis Rambur, 1837 with reconsideration of the systematic position of Heterogynis (Artemisogynis) subg. n. andalusica (Daniel, 1966) comb. n. (Lepidoptera: Zygaenoidea, Heterogynidae). NEVA 2017, 38, 79–84. [Google Scholar]
- De Freina, J.J. A faunistic study of the Heterogynis penella (Hübner, [1818,1819])-species group on the Balkan peninsula with description of Heterogynis zikici nov.sp. (Lepidoptera: Zygaenoidea, Heterogynidae). Entomofauna 2018, 39, 59–71. [Google Scholar]
- Kownacki, A.; Szarek-Gwiazda, E.; Woźnicka, O. The importance of scanning electron microscopy (SEM) in taxonomy and morphology of Chironomidae (Diptera). Eur. J. Environ. Sci. 2015, 5, 41–44. [Google Scholar] [CrossRef]
- Prieto, C.G.; Munguira, M.L.; Romo, H. Morphometric analysis of genitalia and wing pattern elements in the genus Cupido (Lepidoptera, Lycaenidae): Are Cupido minimus and C. carswelli different species? Dtsch. Entomol. Z. 2009, 56, 137–147. [Google Scholar] [CrossRef]
- Lumley, L.M.; Sperling, F.A. Integrating morphology and mitochondrial DNA for species delimitation within the spruce budworm (Choristoneura fumiferana) cryptic species complex (Lepidoptera: Tortricidae). Syst. Entomol. 2010, 35, 416–428. [Google Scholar] [CrossRef]
- Cañas-Hoyos, N.; Marquez, E.J.; Saldamando-Benjumea, C.I. Heritability of wing size and shape of the rice and corn strains of Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Neotrop. Entomol. 2016, 45, 411–419. [Google Scholar] [CrossRef]
- Akand, S.; Bashar, M.A.; Rahman, S.; Khan, H.R. Morphometric variation in the species of two subfamilies of lycaenid butterflies (Lepidoptera: Lycaenidae) of Bangladesh. J. Biodivers. Conserv. Bioresour. Manag. 2017, 3, 9–16. [Google Scholar] [CrossRef][Green Version]
- Goonesekera, K.; van der Poorten, G.; Ranawaka, G.R. Morphometry as a tool in species identification: A study with special reference to species of the genus Mycalesis (Lepidoptera: Nymphalidae). J. Natl. Sci. Found. Sri Lanka 2018, 46, 311–328. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Li, H.H. Phylogeny of the superfamily Gelechioidea (Lepidoptera: Obtectomera), with an exploratory application on geometric morphometrics. Zool. Scr. 2020, 49, 307–328. [Google Scholar] [CrossRef]
- Machida, R.J.; Leray, M.; Ho, S.L.; Knowlton, N. Metazoan mitochondrial gene sequence reference datasets for taxonomic assignment of environmental samples. Sci. Data 2017, 4, 170027. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- de Freina, J.J.; Witt, T.J. Familie Heterogynidae. In Die Bombyces und Sphinges der Westpalaearktis; De Freina, J.J., Witt, T.J., Eds.; Edition Forschung and Wissenschaft: Munich, Germany, 1990; pp. 74–80. [Google Scholar]
- de Freina, J.J. Zur kenntnis der biologie und taxonomievon Heterogynis andalusica Daniel, 1966 sowie ergänzende Bemerkungen zum Status von Heterogynis thomas Zilli, 1987 stat. rev. (Lepidoptera, Zygaenoidea, Heterogynidae). Atalanta 2003, 34, 179–192. [Google Scholar]
- de Freina, J.J. Heterogynis jellaba spec. nov. und Heterogynis rifensisspec. nov., zwei neue nordafrikanische Heterogynis-Artenaus Marokko mit ergänzenden Bemerkungen zum Verbreitungsbildund Artenspektrum der Gattung Heterogynis Rambur, 1837 (Lepidoptera, Zygaenoidea, Heterogynidae). Atalanta 2003, 34, 193–208. [Google Scholar]
- de Freina, J.J. Heterogynidae auf dem Balkan, mit Beschreibung von Heterogynis sondereggeri sp. n. aus den Hochlagen des Peloponnes (Lepidoptera: Zygaenoidea, Heterogynidae). Nachr. Entomol. Ver. Apollo 2012, 33, 129–138. [Google Scholar]
- de Freina, J.J. New and supplementary information on the systematics, biology and geographical distribution of Heterogynis canalensis Chapman, 1904 (Lepidoptera: Zygaenoidea, Heterogynidae). Nachr. Entomol. Ver. Apollo 2014, 35, 97–107. [Google Scholar]
- de Freina, J.J. The species-group of Heterogynis penella (Hübner, 1819) on the Iberian Peninsula, with the description of Heterogynis chapmani sp. n. from central Spain (Lepidoptera: Zygaenoidea, Heterogynidae). Nachr. Entomol. Ver. Apollo 2014, 34, 185–194. [Google Scholar]
- de Freina, J.J. Contribution to the species group of Heterogynis penella (Hübner, 1819 [“1816”]) in the Iberian Peninsula: Description of Heterogynis segurana sp. n. and further details on other species (Lepidoptera: Zygaenoidea, Heterogynidae). Nachr. Entomol. Ver. Apollo 2015, 36, 208–216. [Google Scholar]
- de Freina, J.J.; Tschorsnig, H.P. Raupenfliegen (Diptera: Tachinidae) aus Heterogynis spp. (Lepidoptera: Heterogynidae). Nachr. Bayer. Entomol. 2005, 54, 95–100. [Google Scholar]
- Stojanović, D.V.; Ćurčić, S.B.; Stanisavliević, L.Ž.; Orlović, S.S. New and rare moth species (Insecta: Lepidoptera) from Serbia. North-West J. Zool. 2014, 10, 318–324. [Google Scholar]
- Rohlf, F.J. TpsDig—Digitize Landmarks and Outlines. Version 2.31; Department of Ecology, Evolution and Anthropology, Stony Brook, State University: New York, NY, USA, 2017. [Google Scholar]
- Hebert, P.D.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Ten species in one: DNA barcoding reveals cryptic species in the neotropical skipper butterfly Astraptes fulgerator. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed]
- Hajibabaei, M.; Janzen, D.H.; Burns, J.M.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes distinguish species of tropical Lepidoptera. Proc. Natl. Acad. Sci. USA 2006, 103, 968–971. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Hosoishi, S.; Ogata, K. Description and DNA barcoding of Crematogaster fraxatrix Forel, 1911 and two new closely related species from Cambodia and Indonesia (Hymenoptera, Formicidae). ZooKeys 2014, 374, 57–68. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Scheffer, S.J.; Giblin-Davis, R.M.; Taylor, G.S.; Davies, K.A.; Purcell, M.; Lewis, M.L.; Goolsby, J.; Center, T.D. Phylogenetic relationships, species limits, and host specificity of gall-forming Fergusonina flies (Diptera: Fergusoninidae) feeding on Melaleuca (Myrtaceae). Ann. Entomol. Soc. Am. 2004, 97, 1216–1221. [Google Scholar] [CrossRef]
- Noguchi, S.; Ochiai, T. The first record of Cucullia umbratica (Lepidoptera: Noctuidae) from Japan. Biodivers. Data J. 2019, 7, 34197. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, 636–641. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Matoničkin, I.; Erben, R. Opća Zoologija, 4th ed.; Školska knjiga: Zagreb, Croatia, 2002. [Google Scholar]
- Lakušić, D. (Ed.) Staništa Srbije, Rezultati Projekta, “Harmonizacija Nacionalne Nomenklature u Klasifikaciji Staništa sa Standardima Međunarodne Zajednice”; Institute of Botany and Botanical Garden Jevremovac, Faculty of Biology, University of Belgrade, the Ministry of Science and Environment the Republic of Serbia: Belgrade, Serbia, 2005. [Google Scholar]
- Momčilović Petronijević, A.; Trajković, S.; Ranđelović, V. Anthropogenic effects on environment. Case study: Peat islands of lake Vlasina. FU Arch. Civ. Eng. 2009, 7, 95–105. [Google Scholar] [CrossRef]
- de Freina, J.J. Weitere erkenntnisse über die verbreitung der artengruppe von Heterogynis penella (Hübner, [1818,1819]) auf dem Balkan mit beschreibung von Heterogynis rakosyi sp. n. (Lepidoptera: Zygaenoidea, Heterogynidae). Nachr. Entomol. Ver. Apollo. 2019, 40, 65–74. [Google Scholar]
- Mutanen, M.; Kivelä, S.M.; Vos, R.A.; Doorenweerd, C.; Ratnasingham, S.; Hausmann, A.; Huemer, P.; Dincă, V.; Van Nieukerken, E.J.; Lopez-Vaamonde, C. Species-level para- and polyphyly in DNA barcode gene trees: Strong operational bias in European Lepidoptera. Syst. Biol. 2016, 65, 1024–1040. [Google Scholar] [CrossRef] [PubMed]
- Previšić, A.; Walton, C.; Kučinić, M.; Mitrikeski, P.T.; Kerovec, M. Pleistocene divergence of Dinaric Drusus endemics (Trichoptera, Limnephilidae) in multiple microrefugia within the Balkan Peninsula. Mol. Ecol. 2009, 18, 634–647. [Google Scholar] [CrossRef]
- Dapporto, L. Speciation in Mediterranean refugia and post-glacial expansion of Zerynthia polyxena (Lepidoptera, Papilionidae). J. Zool. Syst. Evol. Res. 2010, 48, 229–237. [Google Scholar]
- Radenković, S.; Šašić Zorić, L.; Djan, M.; Obreht Vidaković, D.; Ačanski, J.; Ståhls, G.; Veličković, N.; Markov, Z.; Petanidou, T.; Kočiš Tubić, N.; et al. Cryptic speciation in the Merodon luteomaculatus complex (Diptera: Syrphidae) from the eastern Mediterranean. J. Zool. Syst. Evol. Res. 2018, 56, 170–191. [Google Scholar] [CrossRef]
- Vrbica, M.; Petrović, A.; Pantelić, D.; Krmpot, A.J.; Rabasović, M.D.; Pavlović, D.; Jovanić, S.; Guéorguiev, B.; Goranov, S.; Vesović, N.; et al. The genus Pheggomisetes Knirsch, 1923 (Coleoptera: Carabidae: Trechinae) in Serbia: Taxonomy, morphology and molecular phylogeny. Zool. J. Linn. Soc. 2018, 183, 347–371. [Google Scholar] [CrossRef]
- Drekić, M.; Poljaković-Pajnik, L.; Kovačević, B.; Milović, M.; Pilipović, A.; Vasić, V. Sessile oak fauna of Cynipid gall wasps of Mt Fruška Gora. Poplar 2020, 205, 59–66. [Google Scholar]
- Kereši, T. Recent records of the Cypress Jewel beetle—Lamprodila (Palmar) festiva (Linnaeus, 1767)(Coleoptera: Buprestidae) in Serbia. Poplar 2020, 205, 25–31. [Google Scholar] [CrossRef]
- Tanovski, V.; Matović, B.; Kesić, L.; Stojanović, D.B. A review of the influence of climate change on coniferous forests in the Balkan peninsula. Poplar 2022, 210, 41–64. [Google Scholar] [CrossRef]

















| PCs | Eigenvalue | % Total (Variance) | Cumulative (%) |
|---|---|---|---|
| PC1 | 4.6324 | 77.21 | 77.21 |
| PC2 | 0.87283 | 14.55 | 91.75 |
| PC3 | 0.36108 | 6.02 | 97.77 |
| PC4 | 0.11374 | 1.90 | 99.67 |
| PC5 | 0.01984 | 0.33 | 99.99 |
| PC6 | 0.00012 | 0.00 | 100.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stojanović, D.V.; Galović, V.; Terzin, T.; Ačanski, J.; Vidović, M.; Orlović, S. The Genus Heterogynis Rambur, 1866 (Heterogynidae, Lepidoptera): Congruence of Molecular, Morphological and Morphometric Evidence Reveal New Species in Serbia. Insects 2023, 14, 455. https://doi.org/10.3390/insects14050455
Stojanović DV, Galović V, Terzin T, Ačanski J, Vidović M, Orlović S. The Genus Heterogynis Rambur, 1866 (Heterogynidae, Lepidoptera): Congruence of Molecular, Morphological and Morphometric Evidence Reveal New Species in Serbia. Insects. 2023; 14(5):455. https://doi.org/10.3390/insects14050455
Chicago/Turabian StyleStojanović, Dejan V., Vladislava Galović, Tomislav Terzin, Jelena Ačanski, Marija Vidović, and Saša Orlović. 2023. "The Genus Heterogynis Rambur, 1866 (Heterogynidae, Lepidoptera): Congruence of Molecular, Morphological and Morphometric Evidence Reveal New Species in Serbia" Insects 14, no. 5: 455. https://doi.org/10.3390/insects14050455
APA StyleStojanović, D. V., Galović, V., Terzin, T., Ačanski, J., Vidović, M., & Orlović, S. (2023). The Genus Heterogynis Rambur, 1866 (Heterogynidae, Lepidoptera): Congruence of Molecular, Morphological and Morphometric Evidence Reveal New Species in Serbia. Insects, 14(5), 455. https://doi.org/10.3390/insects14050455