
Are Botanical Biopesticides Safe for Bees (Hymenoptera, Apoidea)?
 , , ,
, , ,  ,
, 
Simple Summary
Abstract
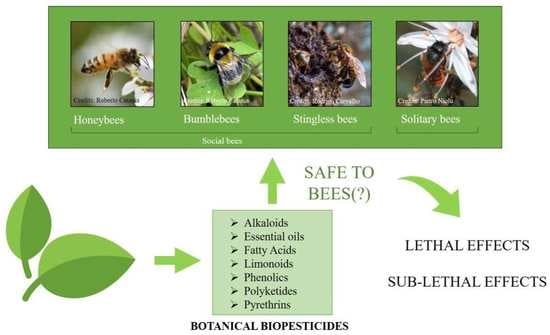
1. Introduction
2. Botanical Biopesticides
3. Risk Assessment of Biopesticides on Bees
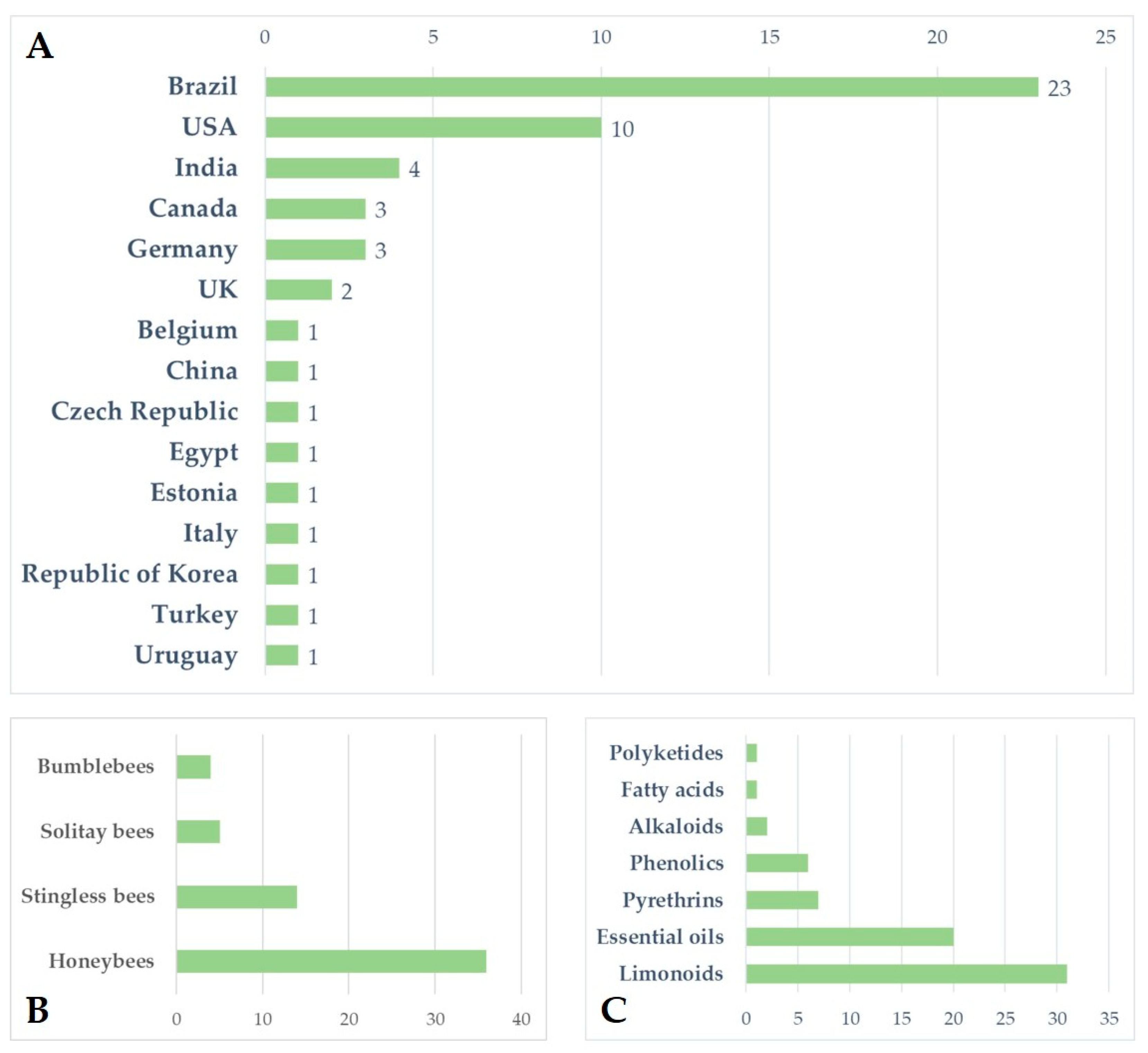
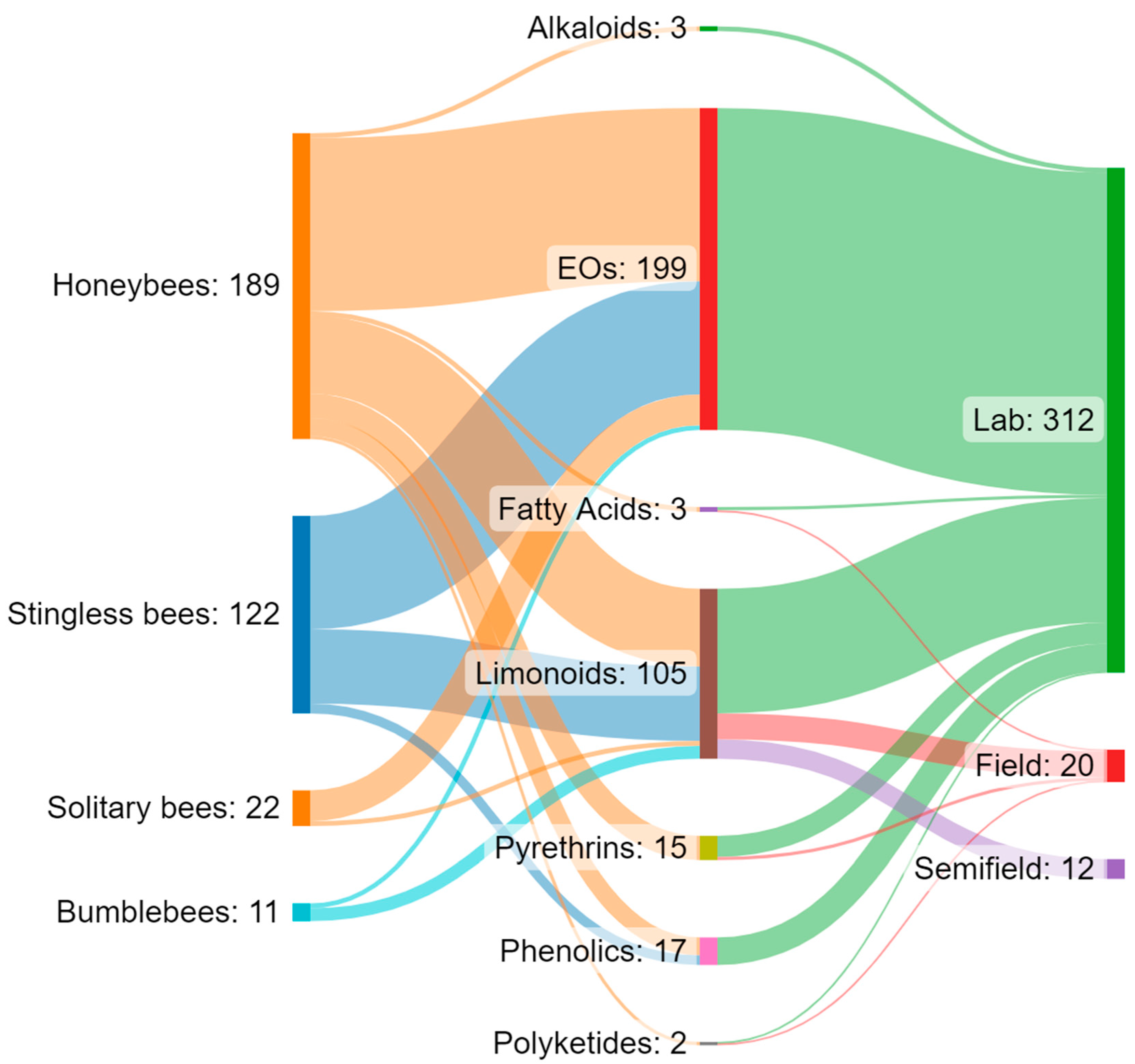
4. Materials and Methods
5. Effects of Botanical Biopesticides on Eusocial Bees
5.1. Honeybees
5.2. Bumblebees
5.3. Stingless Bees
6. Effects of Botanical Biopesticides on Solitary Bees

{kind=link}
{kind=link}
{kind=link}
| Bee Species | Botanical Substance | Assay | Application | Target * | Effects | Country | Year | References |
|---|---|---|---|---|---|---|---|---|
| Social species | ||||||||
| Honeybees (Apis spp.) | ||||||||
| Apis cerana cerana | Azadirachtin | laboratory (L, S) | ingestion | adults | increase in mortality at the higher doses, anti-feeding and inhibition on the immune response | China | 2022 | [115] ** |
| Apis cerana indica | Annonin, azadirachtin | laboratory (L) and field | topical, crop spraying | adults | increase in mortality with both compounds, reduction of the number and speed of foraging bees with annonin | India | 2019 | [106] |
| Apis cerana indica | Azadirachtin | field | crop spraying | adults | reduction of the number of foraging bees | India | 2010 | [91] |
| Apis dorsata | Azadirachtin | field | crop spraying | adults | reduction of the number of foraging bees | India | 2010 | [91] |
| Apis mellifera | Azadirachtin | laboratory (L) | ingestion and topical | larvae, adults | increase in mortality, larvae more susceptible than adults | Brazil | 2016 | [118] ** |
| Apis mellifera | Sabadilla dust | laboratory (L) | contact | adults | increase in mortality | USA | 1958 | [95] |
| Apis mellifera | Aramite (blend of natural oils) | laboratory (L) | contact | adults | low increase in mortality | USA | 1952 | [96] |
| Apis mellifera | Formulations containing pyrethrins, rotenone, and pine oil, three formulations containing pyrethrins | laboratory (L, S) | spray | adults | increase in mortality and knockdown effects for all formulations | USA | 1990 | [126] |
| Apis mellifera | Aramite, pyrethrins, rotenone, ryania, and sabadilla dust | laboratory (L) | contact | adults | increase in mortality with sabadilla, medium and low increase in mortality with the other compounds | USA | 1954 | [94] |
| Apis mellifera | Pyrethrum | field | spray on cage and colony | adults, colony | low increase in mortality | USA | 1979 | [128] |
| Apis mellifera | Mentha piperita, Origanum vulgare, Thymus vulgaris, and Zingiber officinale EOs | laboratory (L, S) | topical, contact, ingestion | adults | increase in mortality with O. vulgare, and T. vulgaris, reduction of movements with O. vulgare | Brazil | 2020 | [102] ** |
| Apis mellifera | Neem oil, pyroligneous extract, and rotenone | laboratory (L) | topical, ingestion | adults | reduction in survival with rotenone on topical application | Brazil | 2012 | [97] |
| Apis mellifera | Azadirachtin | field | crop spraying | adults | no reduction of the numbers of foraging bees | USA | 2004 | [108] |
| Apis mellifera | Pyrethrum extract | laboratory (L) | fumigation | adults | no effects on mortality | USA | 1930 | [127] ** |
| Apis mellifera | Rotenone, and pyrethrum extract | laboratory (L) | spray | adults | increase in mortality | USA | 1932 | [124] ** |
| Apis mellifera | Azadirachtin | laboratory (L, S) | ingestion | adults | high mortality, effects on haemolymph amino acid composition | Egypt | 2015 | [114] |
| Apis mellifera | Neem oil | laboratory (L) | ingestion and topical | larvae | reduction in survival | India | 2022 | [119] ** |
| Apis mellifera | Neem oil | laboratory (L) | topical, ingestion | adults | reduction in survival in contact application | USA | 2005 | [116] |
| Apis mellifera | Pellitorine extracted from Piper tuberculatum | laboratory (L) and field | ingestion and topical | larvae, adults | high mortality at the highest rates | Brazil | 2003 | [105] ** |
| Apis mellifera | Azadirachtin | laboratory (L, S) | contact, ingestion | adults | increase in mortality, no repellent effects, reduction in flight ability | Brazil | 2020 | [117] |
| Apis mellifera | Neem seed extract | laboratory (S) and field | ingestion, crop spraying | adults | food repellency in laboratory bioassays, however no effects on the number of the foraging bees in the field | Canada | 1994 | [112] ** |
| Apis mellifera | Neem oil | field | ingestion and topical | larvae | increase in mortality at the higher concentration | Canada | 1996 | [113] ** |
| Apis mellifera | century plant, citronella, garlic, parsley, rue, and tobacco extracts | laboratory (S) and field | ingestion | adults | repellent effects in laboratory and in field | Brazil | 2004 | [104] ** |
| Apis mellifera | Pyrethrum extract, pyrethrum extract in nanoparticles | laboratory (L, S) | ingestion | adults | reduction in survival, morphological alterations in the epithelium of midgut | Brazil | 2019 | [129] |
| Apis mellifera | Azadirachtin | field | crop spraying | adults | reduction of the number of foraging bees | India | 2010 | [91] |
| Apis mellifera | Agave americana, Anadenanthera colubrina, and Nicotiana tabacum extracts | laboratory (L, S) | contact, ingestion | adults | increased mortality with N. tabacum, low increase in mortality with the other compounds, no effects on respiration rates or flight | Brazil | 2020 | [93] |
| Apis mellifera | Echinodorus grandiflorus, Matricaria chamomilla, Origanum majorana, and Punica granatum extracts | laboratory (L, S) | contact, ingestion, spray | adults | increase in mortality and reduction of the length of mesenteric cells with O. majorana and P. granatum | Brazil | 2020 | [103] ** |
| Apis mellifera | Neem oil | laboratory (L) | contact | adults | increase in mortality | India | 2017 | [107] |
| Apis mellifera | Neem seed extracts | laboratory (L, S) | ingestion and topical | larvae | effects on survival of larvae, no anti-feeding effects, morphological alteration | Germany | 1980 | [109] ** |
| Apis mellifera | Azadirachtin | laboratory (L, S) | ingestion and topical | larvae | increase in mortality, no anti-feeding effects, morphological alteration | Germany | 1982 | [110] ** |
| Apis mellifera | Geraniol and Cymbopogon martinii EO | laboratory (L, S) | topical, ingestion | adults | no effects on mortality, no effects on immune response, on behaviour, and locomotion | Brazil | 2018 | [99] |
| Apis mellifera | Neem seed extracts | field | crop spraying | colony | effects on the hatching observed in the smaller hives, morphological alteration, non-repellent effects on treated flowers | Germany | 1987 | [111] ** |
| Apis mellifera | Azadirachtin | semi-field | crop granules, crop spraying | colony | no effects on mortality, reduction in foraging activity and brood development with spray treatment | Czech Republic | 2005 | [120] ** |
| Apis mellifera | Pyrethrins and rotenone | laboratory (L) | topical, ingestion | adults | increased mortality with both compounds | UK | 1978 | [125] |
| Apis mellifera | Azadirachtin | field | ingestion | colony | colony overwintering failure, no effects on brood and queen development | UK | 2005 | [121] |
| Apis mellifera | Azadirachtin | field | crop spraying | adults | effects on flower visitation rates | Brazil | 2019 | [122] |
| Apis mellifera | Artemisia absinthium, and Eupatorium buniifolium EOs | laboratory (L) | topical, contact | adults | no effects on mortality in the topical test, increased mortality in the contact test | Uruguay | 2017 | [98] ** |
| Apis mellifera | Andiroba, citronella, eucalyptus, and neem oil, garlic extract, and rotenone | laboratory (L, S) | contact, ingestion and topical | larvae, adults | increase in mortality of bee larvae with andiroba, neem oil, and garlic extract, reduction of body mass of adults, reduction in walking activity and repellent effects in adult workers | Brazil | 2015 | [100] ** |
| Bumblebees (Bombus spp.) | ||||||||
| Bombus terrestris | Azadirachtin | laboratory (L, S) | ingestion | adults, microcolony | increase in mortality, repellent effects, reduction in egg-laying, in production of drones, and in ovarian length | Belgium | 2015 | [61] |
| Bombus terrestris | Azadirachtin | laboratory (L) | ingestion | adults | increase in mortality | Turkey | 2022 | [137] ** |
| Bombus terrestris | mixture of Perilla frutescens var. crispa extracts and phytoncide oil | laboratory (L, S) | contact | adults | high mortality, no effects on walking behaviour, reduction in gene expression | Republic of Korea | 2018 | [139] ** |
| Bombus terrestris | Azadirachtin | field | ingestion | adults | reduction in pollen foraging | Estonia | 2009 | [138] ** |
| Stingless bees (Meliponini) | ||||||||
| Melipona quadrifasciata | Azadirachtin | laboratory (L, S) | ingestion and topical | larvae | increase in mortality at higher doses, development of deformed pupae and adults | Brazil | 2015 | [153] ** |
| Melipona quadrifasciata | Azadirachtin | laboratory (L, S) | contact, ingestion | adults | no effects on mortality, no anti-feeding effects, modifications in walking behaviour, no effects on flight and respiration rate | Brazil | 2017 | [62] ** |
| Melipona quadrifasciata | Azadirachtin | laboratory (S) | ingestion | adults | reduction of gene expression of vitellogenin (Vg), increase of the number of haemocytes | Brazil | 2021 | [156] ** |
| Nannotrigona aff. testaceicornis | Lippia sidoides EO, and main compounds | laboratory (L, S) | topical | adults | low increase in mortality, low reduction in locomotion ability and flight orientation, avoidance effects | Brazil | 2021 | [149] |
| Nannotrigona testaceicornis | Andiroba, citronella, eucalyptus, and neem oil, garlic extract, and rotenone | laboratory (S) | contact | adults | no effects on handling behaviour | Brazil | 2010 | [152] ** |
| Partamona helleri | Azadirachtin | laboratory (L, S) | contact, ingestion | adults | no effects on mortality, anti-feeding effects, no effects on walking, flight and respiration rate | Brazil | 2017 | [62] ** |
| Partamona helleri | Azadirachtin | laboratory (L, S) | ingestion and topical | larvae of queens | increase of mortality at the higher doses, delayed development and production of deformed queens, no effects on walking behaviour, reduction in the ovarian morphometry | Brazil | 2018 | [63] |
| Partamona helleri | Agave americana, Anadenanthera colubrina, and Nicotiana tabacum extracts | laboratory (L, S) | contact, ingestion | adults | increased mortality with N. tabacum, low increase in mortality with the other compounds, no effects on respiration rates or flight | Brazil | 2020 | [93] |
| Partamona helleri | Azadirachtin | laboratory (L, S) | contact, ingestion | adults | low increase in mortality, no effects on overall group activity, reduction of flight take-off of worker | Brazil | 2015 | [154] |
| Plebeia sp. | Azadirachtin | field | crop spraying | adults | no effects on flower visitation rates | Brazil | 2019 | [122] |
| Scaptotrigona xanthotrica | Azadirachtin | laboratory (L, S) | contact, ingestion | adults | low increase in mortality, no effects on overall group activity, reduction of flight take-off of worker | Brazil | 2015 | [154] |
| Tetragonisca angustula | Azadirachtin | laboratory (S), and semi-field | contact | adults and colony | reduction in gene expression of esterase isoenzymes, changes in protein synthesis | Brazil | 2020 | [155] ** |
| Tetragonisca angustula | Corymbia citriodora EO | laboratory (L) | topical | adults | increase in mortality | Brazil | 2018 | [150] ** |
| Tetragonisca angustula | Artemisia annua EO | laboratory (L) | topical | adults | increase in mortality | Brazil | 2018 | [151] ** |
| Tetragonisca angustula | Andiroba, citronella, eucalyptus, and neem oil, garlic extract, and rotenone | laboratory (S) | contact | adults | no effects on handling behaviour | Brazil | 2010 | [152] ** |
| Trigona hyalinata | Mentha piperita, Origanum vulgare, Thymus vulgaris, and Zingiber officinale EOs | laboratory (L, S) | topical, contact, ingestion | adults | low increase in mortality, reduction in movements with oregano and thyme EOs | Brazil | 2020 | [102] ** |
| Trigona spinipes | Azadiracha indica, Lippiasidoides, Sapindus saponaria, Anonna squamosa, Cymbopogon winterianum, Corimbia citriodora, Jatropha curcas, Ricinus communis leaf and seed extracts | laboratory (L) | topical | adults | increase in mortality with A. indica, A. squamosa, and R. communis | Brazil | 2012 | [148] ** |
| Solitary species | ||||||||
| Halictus sp. *** | Azadirachtin | field | crop spraying | adults | reduction of flower visitation rates | Brazil | 2019 | [122] |
| Megachile rotundata | Ajwain, basil, cinnamon, clove, coriander, cumin, fenugreek, fennel, ginger, nutmeg, oregano, rosemary, sage, thyme, and turmeric powders (containing EOs) | laboratory (L) | contact | adult males | increase in mortality | Canada | 2020 | [57] ** |
| Osmia cornifrons | Wintergreen oil | laboratory (L) | topical, contact | adults | increase in mortality | USA | 2009 | [158] ** |
| Osmia cornuta | Quassia amara extract | laboratory (L) | contact | eggs, larvae | increase in mortality | Italy | 2003 | [157] |
| Osmia lignaria | Neem oil | laboratory (L) | topical, ingestion | adults | increase in mortality | USA | 2005 | [116] |
| Group | Bee Species | Botanical Substance | Target | Application | Toxicological Parameters | References |
|---|---|---|---|---|---|---|
| HONEYBEES | Apis cerana indica | Annonin | adults | topical | LC50(%)(72 h): 1.22 | [106] |
| Azadirachtin | adults | topical | LC50(%)(72 h): 1.64 | |||
| Apis mellifera | Mentha piperita EO | adults | contact | LC50(%)(24 h): 13.35 | [102] | |
| adults | topical | LD50(%)(24 h): 12.58 | ||||
| Origanum vulgare EO | adults | contact | LC50(%)(24 h): 0.95 | |||
| adults | topical | LD50(%)(24 h): 2.03 | ||||
| Thymus vulgaris EO | adults | contact | LC50(%)(24 h): 2.61 | |||
| adults | topical | LD50(%)(24 h): 3.30 | ||||
| Zingiber officinale EO | adults | contact | LC50(%)(24 h): 22.01 | |||
| adults | topical | LD50(%)(24 h): 17.98 | ||||
| Pellitorine extracted from Piper tuberculatum | larvae | ingestion and topical | LD50 (μg a.i./bee)(96 h): 0.8048 | [105] | ||
| adults | ingestion | LD50 (μg a.i./bee)(96 h): 0.759 | ||||
| topical | LD50 (μg a.i./bee)(96 h): 0.357 | |||||
| Neem oil | I instar larvae | ingestion and topical | LD50 (μg a.i./g)(6 d): 37 | [113] | ||
| IV instar larvae | ingestion and topical | LD50 (μg a.i./g)(10 d): 61 | ||||
| Nicotine extracted from Nicotiana tabacum | adults | contact | LC50 (ng/bee)(48 h): 60.15 | [93] | ||
| adults | ingestion | LC50 (ng/bee)(48 h): 32.45 | ||||
| β-Caryophyllene extracted from Agave americana | adults | contact | LC50 (ng/bee)(48 h): 127.4 | |||
| adults | ingestion | LC50 (ng/bee)(48 h): 111.2 | ||||
| Lupeol extracted from Anadenanthera colubrina | adults | contact | LC50 (ng/bee)(48 h): 222.5 | |||
| adults | ingestion | LC50 (ng/bee)(48 h): 210.1 | ||||
| Cymbopogon martinii EO | adults | ingestion | LD50 (μg/bee)(24 h): 465 | [99] | ||
| adults | topical | LD50 (μg/bee)(24 h): 73 | ||||
| Geraniol | adults | ingestion | LD50 (μg/bee)(24 h): 290 | |||
| adults | topical | LD50 (μg/bee)(24 h): 43 | ||||
| Pyrethrins | adults | ingestion | LD50 (μg/bee): 0.29–0.13 | [125] | ||
| adults | topical | LD50 (μg/bee): 0.15 | ||||
| Rotenone | adults | ingestion | LD50 (μg/bee): >60 | |||
| adults | topical | LD50 (μg/bee): >30 | ||||
| Artemisia absinthium EO | adults | topical | LD50 (μg/bee)(24 h): 252 | [98] | ||
| adults | complete exposure | LD50 (mg/cm2)(24 h): 0.15 | ||||
| Eupatorium buniifolium EO | adults | topical | LD50 (μg/bee)(24 h): 197 | |||
| adults | complete exposure | LD50 (mg/cm2)(24 h): 0.26 | ||||
| STINGLESS BEES | Nannotrigona aff. testaceicornis | Lippia sidoides EO | adults | topical | LD50 (μg/bee)(24 h): 33.7 | [149] |
| Thymol (compound of Lippia sidoides EO) | adults | topical | LD50 (μg/bee)(24 h): 33.6 | |||
| ρ—cymene (compound of Lippia sidoides EO) | adults | topical | LD50 (μg/bee)(24 h): 198 | |||
| (E)—caryophyllene (compound of Lippia sidoides EO) | adults | topical | LD50 (μg/bee)(24 h): 306 | |||
| Partamona helleri | Nicotine extracted from Nicotiana tabacum | adults | contact | LC50 (ng/bee)(48 h): 44.32 | [93] | |
| adults | ingestion | LC50 (ng/bee)(48 h): 38.76 | ||||
| β-Caryophyllene extracted from Agave americana | adults | contact | LC50 (ng/bee)(48 h): 122.2 | |||
| adults | ingestion | LC50 (ng/bee)(48 h): 117.1 | ||||
| Lupeol extracted from Anadenanthera colubrina | adults | contact | LC50 (ng/bee)(48 h): 200.1 | |||
| adults | ingestion | LC50 (ng/bee)(48 h): 212.2 | ||||
| Trigona hyalinata | Mentha piperita EO | adults | contact | LC50(%)(24 h): 21.61 | [102] | |
| adults | topical | LD50(%)(24 h): 16.38 | ||||
| Origanum vulgare EO | adults | contact | LC50(%)(24 h): 7.14 | |||
| adults | topical | LD50(%)(24 h): 4.57 | ||||
| Thymus vulgaris EO | adults | contact | LC50(%)(24 h): 8.29 | |||
| adults | topical | LD50(%)(24 h): 6.53 | ||||
| Zingiber officinale EO | adults | contact | LC50(%)(24 h): 24.17 | |||
| adults | topical | LD50(%)(24 h): 32.65 |
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Michener, C.D. The Bees of the World, 2nd ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 2007; p. 953. [Google Scholar]
- Orr, M.C.; Hughes, A.C.; Chesters, D.; Pickering, J.; Zhu, C.D.; Ascher, J.S. Global Patterns and Drivers of Bee Distribution. Curr. Biol. 2021, 31, 451–458.e4. [Google Scholar] [CrossRef]
- Ollerton, J.; Winfree, R.; Tarrant, S. How Many Flowering Plants Are Pollinated by Animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef]
- Kremen, C. The Value of Pollinator Species Diversity Most Crop-Visiting Species Are Needed to Ensure High Levels of Crop Pollination. Science 2018, 359, 741–742. [Google Scholar] [CrossRef]
- Porto, R.G.; de Almeida, R.F.; Cruz-Neto, O.; Tabarelli, M.; Viana, B.F.; Peres, C.A.; Lopes, A.V. Pollination Ecosystem Services: A Comprehensive Review of Economic Values, Research Funding and Policy Actions. Food Secur. 2020, 12, 1425–1442. [Google Scholar] [CrossRef]
- Danforth, B.N.; Minckley, R.L.; Neff, J.L. The Solitary Bees: Biology, Evolution, Conservation; Princeton University Press, Princeton and Oxford: Princeton, NJ, USA, 2019; p. 488. [Google Scholar] [CrossRef]
- Sgolastra, F.; Hinarejos, S.; Pitts-Singer, T.L.; Boyle, N.K.; Joseph, T.; Luckmann, J.; Raine, N.E.; Singh, R.; Williams, N.M.; Bosch, J. Pesticide Exposure Assessment Paradigm for Solitary Bees. Environ. Entomol. 2019, 48, 22–35. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 340, 1608–1611. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Conrad, K.F.; Warren, M.S.; Fox, R.; Parsons, M.S.; Woiwod, I.P. Rapid Declines of Common, Widespread British Moths Provide Evidence of an Insect Biodiversity Crisis. Biol. Conserv. 2006, 132, 279–291. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Potts, S.G.; Roberts, S.P.M.; Dean, R.; Marris, G.; Brown, M.A.; Jones, R.; Neumann, P.; Settele, J. Declines of Managed Honey Bees and Beekeepers in Europe. J. Apic. Res. 2010, 49, 15–22. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Bommarco, R.; Breeze, T.D.; Carvalheiro, L.G.; Franzén, M.; González-Varo; Schweiger, O. Status and Trends of European Pollinators; Pensoft Publishers: Sofia, Bulgaria, 2015; ISBN 9789546427625. [Google Scholar]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee Declines Driven by Combined Stress from Parasites, Pesticides, and Lack of Flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 Percent Decline over 27 Years in Total Flying Insect Biomass in Protected Areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef]
- Seibold, S.; Gossner, M.M.; Simons, N.K.; Blüthgen, N.; Müller, J.; Ambarl, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, J.C.; et al. Arthropod Decline in Grasslands and Forests Is Associated with Landscape-Level Drivers. Nature 2019, 574, 671–674. [Google Scholar] [CrossRef]
- Cardoso, P.; Barton, P.S.; Birkhofer, K.; Chichorro, F.; Deacon, C.; Fartmann, T.; Fukushima, C.S.; Gaigher, R.; Habel, J.C.; Hallmann, C.A.; et al. Scientists’ Warning to Humanity on Insect Extinctions. Biol. Conserv. 2020, 242, 108426. [Google Scholar] [CrossRef]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect Decline in the Anthropocene: Death by a Thousand Cuts. Proc. Natl. Acad. Sci. USA 2021, 118, e2023989118. [Google Scholar] [CrossRef]
- Ollerton, J. Pollinator Diversity: Distribution, Ecological Function, and Conservation. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 353–376. [Google Scholar] [CrossRef]
- Reilly, J.R.; Artz, D.R.; Biddinger, D.; Bobiwash, K.; Boyle, N.K.; Brittain, C.; Brokaw, J.; Campbell, J.W.; Daniels, J.; Elle, E.; et al. Crop Production in the USA Is Frequently Limited by a Lack of Pollinators. Proc. R. Soc. B Biol. Sci. 2020, 287, 20200922. [Google Scholar] [CrossRef]
- Nieto, A.; Roberts, S.P.M.; Kemp, J.; Rasmont, P.; Kuhlmann, M.; García Criado, M.; Biesmeijer, J.C.; Bogusch, P.; Dathe, H.H.; De la Rúa, P.; et al. European Red List of Bees; Publication Office of the European Union: Luxembourg, 2014; p. 86. ISBN 9789279445125. [Google Scholar]
- Kopec, K.; Burd, L.A. Pollinators In Peril: A Systematic Status Review of North American and Hawaiian Native Bees. Cent. Biol. Divers. 2017, 14. [Google Scholar]
- Goulson, D.; Nicholls, E. Anthropogenic Influences on Bee Foraging. Science 2022, 375, 970–972. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority). Guidance on the Risk Assessment of Plant Protection Products on Bees (Apis mellifera, Bombus spp. and Solitary Bees). EFSA J. 2013, 11, 3295. [Google Scholar] [CrossRef]
- Bernardes, R.C.; Botina, L.L.; Araújo, R.d.S.; Guedes, R.N.C.; Martins, G.F.; Lima, M.A.P. Artificial Intelligence-Aided Meta-Analysis of Toxicological Assessment of Agrochemicals in Bees. Front. Ecol. Evol. 2022, 10, 845608. [Google Scholar] [CrossRef]
- Mayer, D.F.; Lunden, J.D. Effects of Imidacloprid Insecticide on Three Bee Pollinators. Hortic. Sci. 1997, 29, 93–97. [Google Scholar]
- Blacquière, T.; Smagghe, G.; Van Gestel, C.A.M.; Mommaerts, V. Neonicotinoids in Bees: A Review on Concentrations, Side-Effects and Risk Assessment. Ecotoxicology 2012, 21, 973–992. [Google Scholar] [CrossRef]
- Siviter, H.; Richman, S.K.; Muth, F. Field-Realistic Neonicotinoid Exposure Has Sub-Lethal Effects on Non-Apis Bees: A Meta-Analysis. Ecol. Lett. 2021, 24, 2586–2597. [Google Scholar] [CrossRef]
- Domenica, A.; Maria, A.; Stefania, B.; Alessio, I.; Alberto, L.; Tunde, M.; Rachel, S.; Csaba, S.; Benedicte, V.; Alessia, V. Neonicotinoids and Bees: The Case of the European Regulatory Risk Assessment. Sci. Total Environ. 2017, 579, 966–971. [Google Scholar] [CrossRef]
- Sgolastra, F.; Medrzycki, P.; Bortolotti, L.; Maini, S.; Porrini, C.; Simon-Delso, N.; Bosch, J. Bees and Pesticide Regulation: Lessons from the Neonicotinoid Experience. Biol. Conserv. 2020, 241, 108356. [Google Scholar] [CrossRef]
- Arena, M.; Sgolastra, F. A Meta-Analysis Comparing the Sensitivity of Bees to Pesticides. Ecotoxicology 2014, 23, 324–334. [Google Scholar] [CrossRef]
- Abati, R.; Sampaio, A.R.; Maciel, R.M.A.; Colombo, F.C.; Libardoni, G.; Battisti, L.; Lozano, E.R.; Ghisi, N.d.C.; Costa-Maia, F.M.; Potrich, M. Bees and Pesticides: The Research Impact and Scientometrics Relations. Environ. Sci. Pollut. Res. 2021, 28, 32282–32298. [Google Scholar] [CrossRef]
- Biondi, A.; Mommaerts, V.; Smagghe, G.; Viñuela, E.; Zappalà, L.; Desneux, N. The Non-Target Impact of Spinosyns on Beneficial Arthropods. Pest Manag. Sci. 2012, 68, 1523–1536. [Google Scholar] [CrossRef]
- Borges, S.; Alkassab, A.T.; Collison, E.; Hinarejos, S.; Jones, B.; McVey, E.; Roessink, I.; Steeger, T.; Sultan, M.; Wassenberg, J. Overview of the Testing and Assessment of Effects of Microbial Pesticides on Bees: Strengths, Challenges and Perspectives. Apidologie 2021, 52, 1256–1277. [Google Scholar] [CrossRef]
- Cappa, F.; Baracchi, D.; Cervo, R. Biopesticides and Insect Pollinators: Detrimental Effects, Outdated Guidelines, and Future Directions. Sci. Total Environ. 2022, 837, 155714. [Google Scholar] [CrossRef]
- Erler, S.; Eckert, J.H.; Steinert, M.; Alkassab, A.T. Impact of Microorganisms and Entomopathogenic Nematodes Used for Plant Protection on Solitary and Social Bee Pollinators: Host Range, Specificity, Pathogenicity, Toxicity, and Effects of Experimental Parameters. Environ. Pollut. 2022, 302, 119051. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Spochacz, M.; Adamski, Z. The Role of Botanical Treatments Used in Apiculture to Control Arthropod Pests. Apidologie 2022, 53, 27. [Google Scholar] [CrossRef]
- Copping, L.G.; Duke, S.O. Natural Products That Have Been Used Commercially as Crop Protection Agents. Pest Manag. Sci. 2007, 63, 524–554. [Google Scholar] [CrossRef]
- Seiber, J.N.; Coats, J.; Duke, S.O.; Gross, A.D. Biopesticides: State of the Art and Future Opportunities. J. Agric. Food Chem. 2014, 62, 11613–11619. [Google Scholar] [CrossRef]
- Nakasu, E.Y.T.; Williamson, S.M.; Edwards, M.G.; Fitches, E.C.; Gatehouse, J.A.; Wright, G.A.; Gatehouse, A.M.R. Novel Biopesticide Based on a Spider Venom Peptide Shows No Adverse Effects on Honeybees. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140619. [Google Scholar] [CrossRef]
- Jacobsson, E.; Andersson, H.S.; Strand, M.; Peigneur, S.; Eriksson, C.; Lodén, H.; Shariatgorji, M.; Andrén, P.E.; Lebbe, E.K.M.; Rosengren, K.J.; et al. Peptide Ion Channel Toxins from the Bootlace Worm, the Longest Animal on Earth. Sci. Rep. 2018, 8, 4596. [Google Scholar] [CrossRef]
- US EPA—US Environmental Protection Agency; Health Canada; California Department of Pesticide Regulation. Guidance for Assessing Pesticide Risk to Bees; US EPA: Washington, DC, USA, 2014.
- Giunti, G.; Benelli, G.; Palmeri, V.; Laudani, F.; Ricupero, M.; Ricciardi, R.; Maggi, F.; Lucchi, A.; Guedes, R.N.C.; Desneux, N.; et al. Non-Target Effects of Essential Oil-Based Biopesticides for Crop Protection: Impact on Natural Enemies, Pollinators, and Soil Invertebrates. Biol. Control 2022, 176, 105071. [Google Scholar] [CrossRef]
- Acheuk, F.; Basiouni, S.; Shehata, A.A.; Dick, K.; Hajri, H.; Lasram, S.; Yilmaz, M.; Emekci, M.; Tsiamis, G.; Spona-Friedl, M.; et al. Status and Prospects of Botanical Biopesticides in Europe and Mediterranean Countries. Biomolecules 2022, 12, 311. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical Insecticides, Deterrents, and Repellents in Modern Agriculture and an Increasingly Regulated World. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Singh, A. Biopesticides: Present Status and the Future Prospects. J. Biofertil. Biopestic. 2015, 6, 100–129. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical Insecticides in the Twenty-First Century-Fulfilling Their Promise? Annu. Rev. Entomol. 2020, 65, 233–249. [Google Scholar] [CrossRef]
- Chowański, S.; Adamski, Z.; Marciniak, P.; Rosiński, G.; Büyükgüzel, E.; Büyükgüzel, K.; Falabella, P.; Scrano, L.; Ventrella, E.; Lelario, F.; et al. A Review of Bioinsecticidal Activity of Solanaceae Alkaloids. Toxins 2016, 8, 60. [Google Scholar] [CrossRef]
- Ujvàry, I. Nicotine and Other Insecticidal Alkaloids. In Nicotinoid Insecticides and the Nicotinic Acetylcholine Receptor; Yamamoto, I., Casida, J.E., Eds.; Springer: Tokyo, Japan, 1999; pp. 29–69. [Google Scholar]
- Regnault-Roger, C.; Vincent, C.; Arnason, J.T. Essential Oils in Insect Control: Low-Risk Products in a High-Stakes World. Annu. Rev. Entomol. 2012, 57, 405–424. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Mossa, A.T.H. Green Pesticides: Essential Oils as Biopesticides in Insect-Pest Management. J. Environ. Sci. Technol. 2016, 9, 354–378. [Google Scholar] [CrossRef]
- Campolo, O.; Cherif, A.; Ricupero, M.; Siscaro, G.; Grissa-Lebdi, K.; Russo, A.; Cucci, L.M.; Di Pietro, P.; Satriano, C.; Desneux, N.; et al. Citrus Peel Essential Oil Nanoformulations to Control the Tomato Borer, Tuta absoluta: Chemical Properties and Biological Activity. Sci. Rep. 2017, 7, 13036. [Google Scholar] [CrossRef]
- Sciortino, M.; Scurria, A.; Lino, C.; Pagliaro, M.; D’Agostino, F.; Tortorici, S.; Ricupero, M.; Biondi, A.; Zappalà, L.; Ciriminna, R. Silica-Microencapsulated Orange Oil for Sustainable Pest Control. Adv. Sustain. Syst. 2021, 5, 2000280. [Google Scholar] [CrossRef]
- Ong, M.; Chomistek, N.; Dayment, H.; Goerzen, W.; Baines, D. Insecticidal Activity of Plant Powders against the Parasitoid, Pteromalus venustus, and Its Host, the Alfalfa Leafcutting Bee. Insects 2020, 11, 359. [Google Scholar] [CrossRef]
- Perumalsamy, H.; Chang, K.S.; Park, C.; Ahn, Y.J. Larvicidal Activity of Asarum Heterotropoides Root Constituents against Insecticide-Susceptible and-Resistant Culex pipiens Pallens and Aedes aegypti and Ochlerotatus togoi. J. Agric. Food Chem. 2010, 58, 10001–10006. [Google Scholar] [CrossRef] [PubMed]
- Buxton, T.; Takahashi, S.; Takakura, M.; Niwata, I.; Owusu, E.O.; Kim, C.-S. Insecticidal Activities of Pellitorine Isolated from Zanthoxylum zanthoxyloides Roots against Sitophilus oryzae L. (Coleoptera : Curculionidae). J. Entomol. Zool. Stud. 2017, 5, 163–168. [Google Scholar]
- Morgan, E.D. Azadirachtin, a Scientific Gold Mine. Bioorg. Med. Chem. 2009, 17, 4096–4105. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, W.F.; De Meyer, L.; Guedes, R.N.C.; Smagghe, G. Lethal and Sublethal Effects of Azadirachtin on the Bumblebee Bombus terrestris (Hymenoptera: Apidae). Ecotoxicology 2015, 24, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Bernardes, R.C.; Tomé, H.V.V.; Barbosa, W.F.; Guedes, R.N.C.; Lima, M.A.P. Azadirachtin-Induced Antifeeding in Neotropical Stingless Bees. Apidologie 2017, 48, 275–285. [Google Scholar] [CrossRef]
- Bernardes, R.C.; Barbosa, W.F.; Martins, G.F.; Lima, M.A.P. The Reduced-Risk Insecticide Azadirachtin Poses a Toxicological Hazard to Stingless Bee Partamona helleri (Friese, 1900) Queens. Chemosphere 2018, 201, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Imdorf, A.; Kilchenmann, V.; Bogdanov, S.; Bachofen, B.; Beretta, C. Toxic Effects of Thymol, Camphor, Menthol and Eucalyptol on Varroa jacobsoni Oud and Apis mellifera L. in a Laboratory Test. Apidologie 1995, 26, 27–31. [Google Scholar] [CrossRef]
- Ocampo, D.; Ocampo, R. Bioactividad de La Família Annonaceae. Rev. Univ. Caldas 2006, 1, 135–155. [Google Scholar]
- Ribeiro, L.P.; Zanardi, O.Z.; Gonçalves, G.L.P.; Ansante, T.F.; Yamamoto, P.T.; Vendramim, J.D. Toxicity of an Annonin-Based Commercial Bioinsecticide Against Three Primary Pest Species of Stored Products. Neotrop. Entomol. 2018, 47, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Jeran, N.; Grdiša, M.; Varga, F.; Šatović, Z.; Liber, Z.; Dabić, D.; Biošić, M. Pyrethrin from Dalmatian Pyrethrum (Tanacetum cinerariifolium (Trevir.) Sch. Bip.): Biosynthesis, Biological Activity, Methods of Extraction and Determination. Phytochem. Rev. 2021, 20, 875–905. [Google Scholar] [CrossRef]
- Heard, M.S.; Baas, J.; Dorne, J.L.; Lahive, E.; Robinson, A.G.; Rortais, A.; Spurgeon, D.J.; Svendsen, C.; Hesketh, H. Comparative Toxicity of Pesticides and Environmental Contaminants in Bees: Are Honey Bees a Useful Proxy for Wild Bee Species? Sci. Total Environ. 2017, 578, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Schmolke, A.; Galic, N.; Feken, M.; Thompson, H.; Sgolastra, F.; Pitts-Singer, T.; Elston, C.; Pamminger, T.; Hinarejos, S. Assessment of the Vulnerability to Pesticide Exposures Across Bee Species. Environ. Toxicol. Chem. 2021, 40, 2640–2651. [Google Scholar] [CrossRef]
- Lewis, K.A.; Tzilivakis, J. Wild Bee Toxicity Data for Pesticide Risk Assessments. Data 2019, 4, 98. [Google Scholar] [CrossRef]
- Brittain, C.; Potts, S.G. The Potential Impacts of Insecticides on the Life-History Traits of Bees and the Consequences for Pollination. Basic Appl. Ecol. 2011, 12, 321–331. [Google Scholar] [CrossRef]
- Lima, M.A.P.; Martins, G.F.; Oliveira, E.E.; Guedes, R.N.C. Agrochemical-Induced Stress in Stingless Bees: Peculiarities, Underlying Basis, and Challenges. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2016, 202, 733–747. [Google Scholar] [CrossRef]
- OECD. Bumblebee, Acute Contact Toxicity Test; OECD: Paris, France, 2017; Volume 11. [Google Scholar]
- Gradish, A.E.; Van Der Steen, J.; Scott-Dupree, C.D.; Cabrera, A.R.; Cutler, G.C.; Goulson, D.; Klein, O.; Lehmann, D.M.; Lückmann, J.; O’Neill, B.; et al. Comparison of Pesticide Exposure in Honey Bees (Hymenoptera: Apidae) and Bumble Bees (Hymenoptera: Apidae): Implications for Risk Assessments. Environ. Entomol. 2019, 48, 12–21. [Google Scholar] [CrossRef]
- Auteri, D.; Arce, A.; Ingels, B.; Marchesi, M.; Neri, F.M.; Rundlöf, M.; Wassenberg, J. Analysis of the Evidence to Support the Definition of Specific Protection Goals for Bumble Bees and Solitary Bees. EFSA Support. Publ. 2022, 19, 7125E. [Google Scholar] [CrossRef]
- Uhl, P.; Awanbor, O.; Schulz, R.S.; Brühl, C.A. Is Osmia bicornis an Adequate Regulatory Surrogate? Comparing Its Acute Contact Sensitivity to Apis mellifera. PLoS ONE 2019, 14, e0201081. [Google Scholar] [CrossRef]
- Cham, K.O.; Nocelli, R.C.F.; Borges, L.O.; Viana-Silva, F.E.C.; Tonelli, C.A.M.; Malaspina, O.; Menezes, C.; Rosa-Fontana, A.S.; Blochtein, B.; Freitas, B.M.; et al. Pesticide Exposure Assessment Paradigm for Stingless Bees. Environ. Entomol. 2019, 48, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Uhl, P.; Franke, L.A.; Rehberg, C.; Wollmann, C.; Stahlschmidt, P.; Jeker, L.; Brühl, C.A. Interspecific Sensitivity of Bees towards Dimethoate and Implications for Environmental Risk Assessment. Sci. Rep. 2016, 6, 34439. [Google Scholar] [CrossRef] [PubMed]
- Bireley, R.; Borges, S.; Cham, K.; Epstein, D.; Garber, K.; Hart, C.; Hou, W.; Ippolito, A.; Pistorius, J.; Poulsen, V.; et al. Preface: Workshop on Pesticide Exposure Assessment Paradigm for Non-Apis Bees. Environ. Entomol. 2019, 48, 1–3. [Google Scholar] [CrossRef] [PubMed]
- OECD. Test No. 213: Honeybees, Acute Oral Toxicity Test; OECD Guidel. Test. Chem.: Paris, France, 1998; p. 8. [Google Scholar]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The Sublethal Effects of Pesticides on Beneficial Arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Mommaerts, V.; Smagghe, G. Side-Effects of Pesticides on the Pollinator Bombus: An Overview; In-Tech: Warwick, UK, 2011. [Google Scholar] [CrossRef]
- Lima, M.A.P.; Cutler, G.C.; Mazzeo, G.; Hrncir, M. Editorial: The Decline of Wild Bees: Causes and Consequences. Front. Ecol. Evol. 2022, 10, 1027169. [Google Scholar] [CrossRef]
- OECD. Test No. 75: Guidance Document on the Honey Bee (Apis mellifera L.) Brood Test under Semi-Field Conditions; OECD: Paris, France, 2007. [Google Scholar]
- Kumar Gupta, R.; Reybroeck, W.; Van Veen, J.W.; Gupta, A. Beekeeping for Poverty Alleviation and Livelihood Security: Vol. 1: Technological Aspects of Beekeeping; Springer: Dordrecht, The Netherlands, 2014; ISBN 9789401791991. [Google Scholar]
- Osterman, J.; Aizen, M.A.; Biesmeijer, J.C.; Bosch, J.; Howlett, B.G.; Inouye, D.W.; Jung, C.; Martins, D.J.; Medel, R.; Pauw, A.; et al. Global Trends in the Number and Diversity of Managed Pollinator Species. Agric. Ecosyst. Environ. 2021, 322, 107653. [Google Scholar] [CrossRef]
- Radloff, S.; Hepburn, C.; Hepburn, R.; Fuchs, S.; Hadisoesilo, S.; Tan, K.; Engel, M.S.; Kuznetsov, V. Population Structure and Classification of Apis cerana. Apidologie 2010, 41, 589–601. [Google Scholar] [CrossRef]
- Crane, E. Recent Research on the World History of Beekeeping. Bee World 1999, 80, 174–186. [Google Scholar] [CrossRef]
- IPBES. The Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on Pollinators, Pollination and Food Production; IPBES: Bonn, Germany, 2016. [Google Scholar]
- Pandey, A. Suitability Assessment of Neem Formulations as Biopesticide against Honey Bees and Other Pollinators. J. Ecophysiol. Occup. Health 2010, 10, 49–52. [Google Scholar]
- Detzel, A.; Wink, M. Attraction, Deterrence or Intoxication of Bees (Apis mellifera) by Plant Allelochemicals. Chemoecology 1993, 4, 8–18. [Google Scholar] [CrossRef]
- Cunha Pereira, R.; Faria Barbosa, W.; Pereira Lima, M.A.; Vieira, J.O.L.; Carvalho Guedes, R.N.; Rodrigues da Silva, B.K.; Dias Barbosa, G.M.; Lemes Fernandes, F. Toxicity of Botanical Extracts and Their Main Constituents on the Bees Partamona helleri and Apis mellifera. Ecotoxicology 2020, 29, 246–257. [Google Scholar] [CrossRef]
- Atkins, E.L.; Anderson, L.D. Toxicity of Bees on Contaminated Surfaces. The Series. Separate Containers Are Used for Toxicological Studies with Honeybees. The Pesticide Evaluation Is given in Detail, and Toxicity of Pesticide Dusts to Honeybees 1 Formation Concerning the Effect O. J. Econ. Entomol. 1954, 47, 969–972. [Google Scholar] [CrossRef]
- Anderson, L.D.; Atkins, E.L. Toxicity of Pesticides to Honey Bees in Laboratory and Field Tests in Southern California, 1955–1956. J. Econ. Entomol. 1958, 51, 103–108. [Google Scholar] [CrossRef]
- Anderson, L.D.; Tuft, T.O. Toxicity of Several New Insecticides to Honey Bees. J. Econ. Entomol. 1952, 45, 466–469. [Google Scholar] [CrossRef]
- Efrom, C.F.S.; Redaelli, L.R.; Meirelles, R.N.; Ourique, C.B. Side-Effects of Pesticides Used in the Organic System of Production on Apis mellifera Linnaeus, 1758. Brazilian Arch. Biol. Technol. 2012, 55, 47–53. [Google Scholar] [CrossRef]
- Umpiérrez, M.L.; Paullier, J.; Porrini, M.; Garrido, M.; Santos, E.; Rossini, C. Potential Botanical Pesticides from Asteraceae Essential Oils for Tomato Production: Activity against Whiteflies, Plants and Bees. Ind. Crops Prod. 2017, 109, 686–692. [Google Scholar] [CrossRef]
- Santos, A.C.C.; Cristaldo, P.F.; Araújo, A.P.A.; Melo, C.R.; Lima, A.P.S.; Santana, E.D.R.; de Oliveira, B.M.S.; Oliveira, J.W.S.; Vieira, J.S.; Blank, A.F.; et al. Apis mellifera (Insecta: Hymenoptera) in the Target of Neonicotinoids: A One-Way Ticket? Bioinsecticides Can Be an Alternative. Ecotoxicol. Environ. Saf. 2018, 163, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Xavier, V.M.; Message, D.; Picanço, M.C.; Chediak, M.; Santana Júnior, P.A.; Ramos, R.S.; Martins, J.C. Acute Toxicity and Sublethal Effects of Botanical Insecticides to Honey Bees. J. Insect Sci. 2015, 15, 137. [Google Scholar] [CrossRef]
- Ruffinengo, S.; Eguaras, M.; Floris, I.; Faverin, C.; Bailac, P.; Ponzi, M. LD50 and Repellent Effects of Essential Oils from Argentinian Wild Plant Species on Varroa destructor. J. Econ. Entomol. 2005, 98, 651–655. [Google Scholar] [CrossRef]
- da Silva, I.M.; Zanuncio, J.C.; Brügger, B.P.; Soares, M.A.; Zanuncio, A.J.V.; Wilcken, C.F.; Tavares, W.d.S.; Serrão, J.E.; Sediyama, C.S. Selectivity of the Botanical Compounds to the Pollinators Apis mellifera and Trigona hyalinata (Hymenoptera: Apidae). Sci. Rep. 2020, 10, 4820. [Google Scholar] [CrossRef]
- Potrich, M.; da Silva, R.T.L.; Maciel, R.M.A.; Costa-Maia, F.M.; Lozano, E.R.; Rossi, R.M.; Martins, J.R.; Dallacort, S. Are Plant Extracts Safe for Honey Bees (Apis mellifera)? J. Apic. Res. 2020, 59, 844–851. [Google Scholar] [CrossRef]
- Nicodemo, D.; Nogueira Couto, R.H. Use of Repellents for Honeybees (Apis mellifera L.) in Vitro in the Yellow Passion-Fruit (Passiflora edulis Deg) Crop and in Confined Beef Cattle Feeders. J. Venom. Anim. Toxins Incl. Trop. Dis. 2004, 10, 77–85. [Google Scholar] [CrossRef]
- Miranda, J.; Navickiene, H.M.D.; Nogueira-Couto, R.; de Bortoli, S.A.; Kato, M.; Bolzani, V.D.; Furlan, M. Susceptibility of Apis mellifera (Hymenoptera: Apidae) to Pellitorine, an Amide Isolated from Piper tuberculatum (Piperaceae). Apidologie 2003, 34, 409–415. [Google Scholar] [CrossRef]
- Challa, G.K.; Firake, D.M.; Behere, G.T. Bio-Pesticide Applications May Impair the Pollination Services and Survival of Foragers of Honey Bee, Apis cerana Fabricius in Oilseed Brassica. Environ. Pollut. 2019, 249, 598–609. [Google Scholar] [CrossRef] [PubMed]
- Ratnakar, V.; Rao, S.R.K.; Sridevi, D.; Vidyasagar, B. Contact Toxicity of Certain Conventional Insecticides to European Honeybee, Apis mellifera Linnaeus. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3359–3365. [Google Scholar] [CrossRef]
- Elzen, P.J.; Elzen, G.W.; Lester, G.E. Compatibility of an Organically Based Insect Control Program with Honey Bee (Hymenoptera: Apidae) Pollination in Cantaloupes. J. Econ. Entomol. 2004, 97, 1513–1516. [Google Scholar] [CrossRef] [PubMed]
- Rembold, H.; Sharma, G.K.; Czoppelt, C. Evidence of Growth Disruption in Insects without Feeding Inhibition by Neem Seed Fractions. J. Plant Dis. Prot. 1980, 87, 290–297. [Google Scholar]
- Rembold, H.; Sharma, G.K.; Czoppelt, C.; Schmutterer, H. Azadirachtin: A Potent Insect Growth Regulator of Plant Origin. Z. Angew. Entomol. 1982, 93, 12–17. [Google Scholar] [CrossRef]
- Schmutterer, H.; Holst, H. Untersuchungen Über Die Wirkung Des Angereicherten Und Formulierten Niemsamenextrakts AZT-VR-K Auf Die Honigbiene Apis mellifera L. J. Appl. Entomol. 1987, 103, 208–213. [Google Scholar] [CrossRef]
- Naumann, K.; Isman, M.B.; Currie, R.W. Evaluation of the Repellent Effects of a Neem Insecticide on Foraging Honey Bees and Other Pollinators. Can. Entomol. 1994, 126, 225–230. [Google Scholar] [CrossRef]
- Naumann, K.; Isman, M.B. Toxicity of a Neem (Azadirachta indica A. Juss) Insecticide to Larval Honey Bees. Am. Bee J. 1996, 136, 518–520. [Google Scholar]
- Hassona, N.M.; Kordy, A.M. Relationship between Toxicity of Certain Pesticides to the Honey Bee, Apis mellifera L. (Hymenoptera: Apidea) Foragers and Their Haemolymph Amino Acids. Middle East J. Appl. Sci. 2015, 5, 19–25. [Google Scholar]
- Zhao, K.; Wu, H.; Hou, R.; Wu, J.; Wang, Y.; Huang, S.; Cheng, D.; Xu, H.; Zhang, Z. Effects of Sublethal Azadirachtin on the Immune Response and Midgut Microbiome of Apis cerana cerana (Hymenoptera: Apidae). Ecotoxicol. Environ. Saf. 2022, 229, 113089. [Google Scholar] [CrossRef] [PubMed]
- Ladurner, E.; Bosch, J.; Kemp, W.P.; Maini, S. Assessing Delayed and Acute Toxicity of Five Formulated Fungicides to Osmia lignaria Say and Apis mellifera. Apidologie 2005, 36, 449–460. [Google Scholar] [CrossRef]
- Naiara Gomes, I.; Ingred Castelan Vieira, K.; Moreira Gontijo, L.; Canto Resende, H. Honeybee Survival and Flight Capacity Are Compromised by Insecticides Used for Controlling Melon Pests in Brazil. Ecotoxicology 2020, 29, 97–107. [Google Scholar] [CrossRef]
- Amaral, R.L.; Venzon, M.; Filho, S.M.; Lima, M.A.P. Does Ingestion of Neem-Contaminated Diet Cause Mortality of Honey Bee Larvae and Foragers? J. Apic. Res. 2016, 54, 405–410. [Google Scholar] [CrossRef]
- Kaur, G.; Singh, R.; Singh, A. Impact of Neem Oil on Developmental Stages of Honey Bee Apis mellifera L. Indian J. Entomol. 2022, 84, 783–787. [Google Scholar] [CrossRef]
- Shawki, M.A.-A.; Táborský, V.; Kamler, F.; Kazda, J. Effect of Two NeemAzalTM Formulations on Honeybees under Semi-Field Conditions. Plant Prot. Sci. 2005, 41, 63–72. [Google Scholar] [CrossRef]
- Thompson, H.M.; Wilkins, S.; Battersby, A.H.; Waite, R.J.; Wilkinson, D. The Effects of Four Insect Growth-Regulating (IGR) Insecticides on Honeybee (Apis mellifera L.) Colony Development, Queen Rearing and Drone Sperm Production. Ecotoxicology 2005, 14, 757–769. [Google Scholar] [CrossRef]
- Tschoeke, P.H.; Oliveira, E.E.; Dalcin, M.S.; Silveira-Tschoeke, M.C.A.C.; Sarmento, R.A.; Santos, G.R. Botanical and Synthetic Pesticides Alter the Flower Visitation Rates of Pollinator Bees in Neotropical Melon Fields. Environ. Pollut. 2019, 251, 591–599. [Google Scholar] [CrossRef]
- Calderone, N.W.; Nasr, M.E. Evaluation of a Formic Acid Formulation for the Fall Control of Varroa jacobsoni (Acari: Varroidae) in Colonies of the Honey Bee Apis mellifera (Hymenoptera: Apidae) in a Temperate Climate. J. Econ. Entomol. 1999, 92, 526–533. [Google Scholar] [CrossRef]
- Ginsburg, J.M.; Schmit, J.B. A Comparison between Rotenone and Pyrethrins as Contact Insecticides. J. Econ. Entomol. 1932, 25, 918–922. [Google Scholar] [CrossRef]
- Stevenson, J.H. The Acute Toxicity of Unformulated Pesticides to Worker Honey Bees (Apis mellifera L.). Plant Pathol. 1978, 27, 38–40. [Google Scholar] [CrossRef]
- Appel, A.G. Knockdown Efficiency and Materials’ Compatibility of Wasp and Hornet Spray Formulations to Honey Bees (Hymenoptera: Apidae). J. Econ. Entomol. 1990, 83, 1925–1931. [Google Scholar] [CrossRef]
- Ginsburg, J.M. Test To Determine Toxicity Of Pyrethrum Vapors To Honeybees. J. Agric. Res. 1930, 40, 1053–1057. [Google Scholar]
- Caron, D.M. Effects of Some ULV Mosquito Abatement Insecticides on Honey Bees. J. Econ. Entomol. 1979, 72, 148–151. [Google Scholar] [CrossRef]
- Oliveira, C.R.; Domingues, C.E.C.; de Melo, N.F.S.; Roat, T.C.; Malaspina, O.; Jones-Costa, M.; Silva-Zacarin, E.C.M.; Fraceto, L.F. Nanopesticide Based on Botanical Insecticide Pyrethrum and Its Potential Effects on Honeybees. Chemosphere 2019, 236, 124282. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D. Bumblebees: Behaviour, Ecology, and Conservation; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Cameron, S.A.; Lozier, J.D.; Strange, J.P.; Koch, J.B.; Cordes, N.; Solter, L.F.; Griswold, T.L. Patterns of Widespread Decline in North American Bumble Bees. Proc. Natl. Acad. Sci. USA 2011, 108, 662–667. [Google Scholar] [CrossRef]
- Rollin, O.; Vray, S.; Dendoncker, N.; Michez, D.; Dufrêne, M.; Rasmont, P. Drastic Shifts in the Belgian Bumblebee Community over the Last Century. Biodivers. Conserv. 2020, 29, 2553–2573. [Google Scholar] [CrossRef]
- Rasmont, P.; Coppée, A.; Michez, D.; De Meulemeester, M.T. An Overview of the Bombus terrestris (L. 1758) Subspecies (Hymenoptera: Apidae). Ann. Société Entomol. Fr. 2008, 44, 243–250. [Google Scholar] [CrossRef]
- Velthuis, H.H.W.; Van Doorn, A. A Century of Advances in Bumblebee Domestication and the Economic and Environmental Aspects of Its Commercialization for Pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef]
- Sterk, G.; Cuylaerts, J.; Kolokytha, P. Lethal and Sublethal Effects of Several Formulations of Azadirachtin on IPM Impact R&D Colonies of the Bumblebee Bombus terrestris (Hymenoptera: Apidae). Hazards Pestic. Bees 2018, 462, 39–45. [Google Scholar] [CrossRef]
- Sterk, G.; Hannegraaf, J.; Kolokytha, P. Effects of Chemical and Biological Plant Protection Products on R&D Colonies of the Buff-Tailed Bumblebee Bombus terrestris. In Proceedings of the Hazards of Pesticides to Bees, 14th International Symposium of the ICP-PR Bee Protection Group, Bern, Switzerland, 23–25 October 2019. [Google Scholar]
- Demirozer, O.; Uzun, A.; Yanik, G.; Bulus, I.Y.; Gosterit, A. Investigation of the Efficacy of Some Biopesticides by Food Exposure on Bombus terrestris L. (Hymenoptera: Apidae). J. Apic. Res. 2022, 1–5. [Google Scholar] [CrossRef]
- Koskor, E.; Muljar, R.; Drenkhan, K.; Karise, R.; Bender, A.; Viik, E.; Luik, A.; Mänd, M. The Chronic Effect of the Botanical Insecticide Neem EC on the Pollen Forage of the Bumble Bee Bombus terrestris L. Agron. Res. 2009, 7, 341–346. [Google Scholar]
- Kim, S.; Lee, J.K.; Song, Y.J.; Kang, S.C.; Kim, B.; Choi, I.J.; Lee, D.H. Evaluating Natural Compounds as Potential Insecticides against Three Economically Important Pests, Bemisia tabaci (Hemiptera: Aleyrodidae), Frankliniella occidentalis (Thysanoptera: Thripidae), and Myzus persicae (Hemiptera: Aphididae), on Greenhouse Swee. Appl. Biol. Chem. 2018, 61, 313–323. [Google Scholar] [CrossRef]
- Rasmussen, C.; Thomas, J.C.; Engel, M.S. A New Genus of Eastern Hemisphere Stingless Bees (Hymenoptera: Apidae), with a Key to the Supraspecific Groups of Indomalayan and Australasian Meliponini. Am. Mus. Novit. 2017, 3888, 33. [Google Scholar] [CrossRef]
- Cortopassi-Laurino, M.V.L.I.-F.; Roubik, D.W.; Dollin, A.; Heard, T.; Aguilar, I.; Venturieri, C.G.; Eardley, C.; Nogueira-Neto, P. Global Meliponiculture: Challenges and Opportunities. Apidologie 2006, 37, 275–292. [Google Scholar] [CrossRef]
- Christopher Brown, J.; Albrecht, C. The Effect of Tropical Deforestation on Stingless Bees of the Genus Melipona (Insecta: Hymenoptera: Apidae: Meliponini) in Central Rondonia, Brazil. J. Biogeogr. 2001, 28, 623–634. [Google Scholar] [CrossRef]
- Samejima, H.; Marzuki, M.; Nagamitsu, T.; Nakasizuka, T. The Effects of Human Disturbance on a Stingless Bee Community in a Tropical Rainforest. Biol. Conserv. 2004, 120, 577–587. [Google Scholar] [CrossRef]
- Reyes-Gonzalez, A.; Mora, F.; Porter-Boland, L.; MRamírez, I.M.; Del-Val, E. Stingless Bees (Apidae: Meliponini) at Risk in Western Mexico. Biotropica 2022, 54, 829–838. [Google Scholar] [CrossRef]
- Toledo-Hernández, E.; Peña-Chora, G.; Hernández-Velázquez, V.M.; Lormendez, C.C.; Toribio-Jiménez, J.; Romero-Ramírez, Y.; León-Rodríguez, R. The Stingless Bees (Hymenoptera: Apidae: Meliponini): A Review of the Current Threats to Their Survival. Apidologie 2022, 53, 8. [Google Scholar] [CrossRef]
- Barbosa, W.F.; Smagghe, G.; Guedes, R.N.C. Pesticides and Reduced-Risk Insecticides, Native Bees and Pantropical Stingless Bees: Pitfalls and Perspectives. Pest Manag. Sci. 2015, 71, 1049–1053. [Google Scholar] [CrossRef]
- Tomé, H.V.; Ramos, G.S.; Araújo, M.F.; Santana, W.C.; Santos, G.R.; Guedes, R.N.; Maciel, C.D.; Newland, P.L.; Oliveira, E.E. Agrochemical Synergism Imposes Higher Risk to Neotropical Bees than to Honeybees. R. Soc. Open Sci. 2017, 18, 160866. [Google Scholar] [CrossRef]
- Correia-Oliveira, M.E.; Poderoso, J.C.M.; Ferreira, A.F.; De Olinda, R.A.; Ribeiro, G.T. Impact of Aqueous Plant Extracts on Trigona spinipes (Hymenoptera: Apidae). Sociobiology 2012, 59, 849–858. [Google Scholar]
- Matos, W.B.; Santos, A.C.C.; Lima, A.P.S.; Santana, E.D.R.; Silva, J.E.; Blank, A.F.; Araújo, A.P.A.; Bacci, L. Potential Source of Ecofriendly Insecticides: Essential Oil Induces Avoidance and Cause Lower Impairment on the Activity of a Stingless Bee than Organosynthetic Insecticides, in Laboratory. Ecotoxicol. Environ. Saf. 2021, 209, 111764. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.V.; Farias, E.d.S.; Santos, A.A.; Filomeno, C.A.; dos Santos, I.B.; Barbosa, L.C.A.; Picanço, M.C. Selection of an Essential Oil from Corymbia and Eucalyptus Plants against Ascia monuste and Its Selectivity to Two Non-Target Organisms. Crop Prot. 2018, 110, 207–213. [Google Scholar] [CrossRef]
- Seixas, P.T.L.; Demuner, A.J.; Alvarenga, E.S.; Barbosa, L.C.A.; Marques, A.; Farias, E.d.S.; Picanço, M.C. Bioactivity of Essential Oils from Artemisia against Diaphania hyalinata and Its Selectivity to Beneficial Insects. Sci. Agric. 2018, 75, 519–525. [Google Scholar] [CrossRef]
- Xavier, V.M.; Message, D.; Picanço, M.C.; Bacci, L.; Silva, G.A.; Da Silva Benevenute, J. Impact of Botanical Insecticides on Indigenous Stingless Bees (Hymenoptera: Apidae). Sociobiology 2010, 56, 713–725. [Google Scholar]
- Barbosa, W.F.; Tomé, H.V.V.; Bernardes, R.C.; Siqueira, M.A.L.; Smagghe, G.; Guedes, R.N.C. Biopesticide-Induced Behavioral and Morphological Alterations in the Stingless Bee Melipona quadrifasciata. Environ. Toxicol. Chem. 2015, 34, 2149–2158. [Google Scholar] [CrossRef]
- Tome, H.V.V.; Barbosa, W.F.; Corrêa, A.S.; Gontijo, L.M.; Martins, G.F.; Guedes, R.N.C. Reduced-Risk Insecticides in Neotropical Stingless Bee Species: Impact on Survival and Activity. Ann. Appl. Biol. 2015, 167, 186–196. [Google Scholar] [CrossRef]
- Oliveira, C.V.P.; Sinópolis Gigliolli, A.A.; Galhardo, D.; Moreira, D.R.; Ronqui, L.; Santos, S.A.; Arnault Toledo, V.A.; Ruvolo-Takasusuki, M.C.C. Effects Of Biopesticides In Tetragonisca angustula Latreille (Hymenoptera: Meliponinae) Pollinators. Arq. Ciências Veterinárias Zool. UNIPAR 2020, 23, e2301. [Google Scholar] [CrossRef]
- Viana, T.A.; Barbosa, W.F.; Lourenço, A.P.; Santana, W.C.; Campos, L.O.; Martins, G.F. Changes in Innate Immune Response and Detoxification in Melipona quadrifasciata (Apinae: Meliponini) on Oral Exposure to Azadirachtin and Spinosad. Apidologie 2021, 52, 252–261. [Google Scholar] [CrossRef]
- Tesoriero, D.; Maccagnani, B.; Santi, F.; Celli, G. Toxicity of Three Pesticides on Larval Instars of Osmia cornuta: Preliminary Results. Bull. Insectology 2003, 56, 169–171. [Google Scholar]
- White, J.B.; Park, Y.L.; West, T.P.; Tobin, P.C. Assessment of Potential Fumigants to Control Chaetodactylus krombeini (Acari: Chaetodactylidae) Associated with Osmia cornifrons (Hymenoptera: Megachilidae). J. Econ. Entomol. 2009, 102, 2090–2095. [Google Scholar] [CrossRef]
- Isman, M.B.; Grieneisen, M.L. Botanical Insecticide Research: Many Publications, Limited Useful Data. Trends Plant Sci. 2014, 19, 140–145. [Google Scholar] [CrossRef]
- Santana, C.B.; Souza, J.G.L.; Toledo, A.G.; Alves, L.F.A.; Alves, D.S.; Corrêa, J.M.; Pinto, F.G.S. Antimicrobial and Insecticidal Effects of Essential Oil and Plant Extracts of Myrcia oblongata Dc in Pathogenic Bacteria and Alphitobius diaperinus. Brazilian J. Biol. 2022, 82, e233425. [Google Scholar] [CrossRef]
- Bibiano, C.S.; Alves, D.S.; Freire, B.C.; Vilela Bertolucci, S.K.; Carvalho, G.A. Toxicity of Essential Oils and Pure Compounds of Lamiaceae Species against Spodoptera frugiperda (Lepidoptera: Noctuidae) and Their Safety for the Nontarget Organism Trichogramma pretiosum (Hymenoptera: Trichogrammatidae). Crop Prot. 2022, 158, 106011. [Google Scholar] [CrossRef]
- Stenger, L.D.; Abati, R.; Pawlak, I.G.; Varpechoski, G.O.; De Souza Vismara, E.; Barbosa, L.R.; Wagner Júnior, A.; Lozano, E.R.; Potrich, M. Toxicity of Essential Oil of Eugenia uniflora (L.) to Thaumastocoris peregrinus (Hemiptera: Thaumastocoridae) and Selectivity to the Parasitoid Cleruchoides noackae (Lin & Hubert) (Hymenoptera: Mymaridae). Crop Prot. 2021, 147, 105693. [Google Scholar] [CrossRef]
- Altoe, M.D.; Lima, J.D.A.; Potrich, M.; Battisti, L.; Lozano, E.R. Insecticidal and Repellent Effects of Essential Oil Eugenia uniflora L. (Myrtaceae) on Sitophilus zeamais Mots. (Coleoptera: Curculionidae). Int. J. Trop. Insect Sci. 2022, 7. [Google Scholar] [CrossRef]
- Sgolastra, F.; Medrzycki, P.; Bortolotti, L.; Renzi, M.T.; Tosi, S.; Bogo, G.; Teper, D.; Porrini, C.; Molowny-Horas, R.; Bosch, J. Synergistic Mortality between a Neonicotinoid Insecticide and an Ergosterol-Biosynthesis-Inhibiting Fungicide in Three Bee Species. Pest Manag. Sci. 2017, 73, 1236–1243. [Google Scholar] [CrossRef]
- Tosi, S.; Nieh, J.C. Lethal and Sublethal Synergistic Effects of a New Systemic Pesticide, Flupyradifurone (Sivantow), on Honeybees. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190433. [Google Scholar] [CrossRef] [PubMed]
- Siviter, H.; Bailes, E.J.; Martin, C.D.; Oliver, T.R.; Koricheva, J.; Leadbeater, E.; Brown, M.J.F. Agrochemicals Interact Synergistically to Increase Bee Mortality. Nature 2021, 596, 389–392. [Google Scholar] [CrossRef] [PubMed]


| Sublethal Effects | Honeybees | Bumblebees | Stingless Bees | Solitary Bees |
|---|---|---|---|---|
| Anti-feeding | Azadirachtin (3,2) | Azadirachtin (2,1) | ||
| Brood development | Azadirachtin (3,2) | Azadirachtin (2,2) | ||
| Foraging activity | Annonin (1,1), Azadirachtin (8,5) | Azadirachtin (1,1) | Azadirachtin (1,1) | |
| Gene expression | EOs (1,1) | Azadirachtin (2,2) | ||
| Immunity | Azadirachtin (1,1) | |||
| Locomotory/flight activity | Azadirachtin (2,2), EOs (7,2) | Azadirachtin (7,3), EOs (9,4) | ||
| Morphology and physiology | Azadirachtin (4,4), EOs (4,1), Pyrethrum (2,2) | Azadirachtin (1,1) | Azadirachtin (7,4) | |
| Overwintering | Azadirachtin (1,1) | |||
| Repellence | Azadirachtin (4,1), EOs (10,8) | EOs (1,1) | ||
| Reproduction | Azadirachtin (1,1) |
| Research Gaps | Future Directions |
|---|---|
| Develop specific protocols to assess the lethal and sublethal effects of biopesticides on bees from different species with the inclusion of a positive control Increase the knowledge of species sensitivity distribution regarding chemical and biological pesticides for Apoidea, including in the studies the ground-nesting solitary bees Develop new and better field and semi-field protocols for both synthetic and biopesticides |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catania, R.; Lima, M.A.P.; Potrich, M.; Sgolastra, F.; Zappalà, L.; Mazzeo, G. Are Botanical Biopesticides Safe for Bees (Hymenoptera, Apoidea)? Insects 2023, 14, 247. https://doi.org/10.3390/insects14030247
Catania R, Lima MAP, Potrich M, Sgolastra F, Zappalà L, Mazzeo G. Are Botanical Biopesticides Safe for Bees (Hymenoptera, Apoidea)? Insects. 2023; 14(3):247. https://doi.org/10.3390/insects14030247
Chicago/Turabian StyleCatania, Roberto, Maria Augusta Pereira Lima, Michele Potrich, Fabio Sgolastra, Lucia Zappalà, and Gaetana Mazzeo. 2023. "Are Botanical Biopesticides Safe for Bees (Hymenoptera, Apoidea)?" Insects 14, no. 3: 247. https://doi.org/10.3390/insects14030247
APA StyleCatania, R., Lima, M. A. P., Potrich, M., Sgolastra, F., Zappalà, L., & Mazzeo, G. (2023). Are Botanical Biopesticides Safe for Bees (Hymenoptera, Apoidea)? Insects, 14(3), 247. https://doi.org/10.3390/insects14030247