
Host Plant Specificity in Web-Building Spiders




Abstract
Simple Summary
Abstract
1. Introduction
2. Spider–Plant Associations
2.1. Cursorial Spiders

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spider Family | Spider Species | Plant Family | Plant Species | Association | Region | Source |
|---|---|---|---|---|---|---|
| Oxyopidae | Peucetia flava | Asteraceae | Trichogoniopsis adenantha | Facultative mutualism; reduced herbivores. | Southeast Brazil | Romero et al. [46] |
| Melastomataceae | Rhyncanthera dichotoma | Commensalism/facultative mutualism; protection and significantly reduced herbivory after rainy season. | Southeast Brazil | Morais-Filho and Romero [39,40,47] | ||
| Solanaceae | Solanum thomasiifolium | Facultative mutualism; likely protection. | Southeast Brazil | Jacobucci et al. [57] | ||
| Fabaceae | Mimosa setosa var. paludosa | Facultative mutualism; reduced exophytic herbivory, but not endophytic herbivory. | Southeast Brazil | Sousa-Lopes et al. [45] | ||
| Peucetia rubrolineata | Asteraceae | Trichogoniopsis adenantha | Facultative mutualism; suppressed herbivory. | Southeast Brazil | Romero et al. [46] | |
| Peucetia viridans | Euphorbia-ceae | Cnidoscolus aconitifolius | Preference/Unknown | Southeast Mexico | Arango et al. [62] | |
| Croton ciliatoglandulifer | Preference/Unknown | West Mexico | Corcuera et al. [63] | |||
| Salticidae | Bagheera kiplingi | Fabaceae | Vachellia spp. (myrmecophytes) | Exploitative/Commensalism | Southeast Mexico, Northwest Costa Rica | Meehan et al. [7] |
| Evarcha culicivora | Euphorbia-ceae | Ricinus communis | Unknown/Commensalism | West Kenya | Cross [64] | |
| Lantana camara | Unknown/Commensalism | West Kenya | Cross [64] | |||
| Pelegrina tillandsiae | Bromeliaceae | Tillandsia usneoides | Obligate commensalism; strict association, but no reported costs or benefits. | Southeast USA | Young and Lockley [24] | |
| Psecas chapoda | Bromeliaceae | Bromelia balansae | Facultative mutualism; the spider aids in nitrogen acquisition. | Northeast Bolivia, Northeast Paraguay, South Brazil, Central-West Brazil | Romero [26]; Romero and Vasconcellos-Neto [29]; Romero et al. [31]; Omena and Romero [56] | |
| Sparrasidae | Delena melanochelis | Myrtaceae | Eucalyptus nitens | Unknown/Commensalism | Australia | Agnarsson and Rayor [65] |
| E. regnans | Unknown/Commensalism | Australia | Agnarsson and Rayor [65] | |||
| Thomisidae | Misumenops argenteus | Lamiaceae | Hyptis suaveolens | Unknown/Commensalism | Southeast Brazil | Romero and Vasconcellos-Neto [27] |
| Asteraceae | Trichogoniopsis adenantha | Facultative mutualism; reduced herbivory | Southeast Brazil | Romero and Vasconcellos-Neto [27,55] | ||
| Misumenops pallidus | Orchideaceae | Chloraea alpina | Commensalism | East Argentina | Quintero et al. [66] | |
| Ranunculaceae | Anemone multifida | Commensalism | East Argentina | Gavini et al. [41] | ||
| Misumenops nepenthicola | Nepentha-ceae | Nepenthes gracilis | Unknown/Commensalism | North Borneo | Karl and Bauer [67] | |
| N. rafflesiana | Unknown/Commensalism | North Borneo | Karl and Bauer [67] | |||
| Synaema marlothi | Roridulaceae | Roridula dentata | Obligate kleptoparasitism | Southern South Africa | Anderson and Midgley [38]; Anderson [68] | |
| Synaema obscuripes | Nepentha-ceae | Nepenthes madagascariensis | Unknown | Southeast Madagascar | Rembold et al. [36] | |
| Thomisus nepenthephilus | Nepentha-ceae | Nepenthes gracilis | Obligate, conditional facilitative mutualism | North Singapore | Lim et al. [34]; Lam and Tan [37] |
2.2. Web-Building Spiders
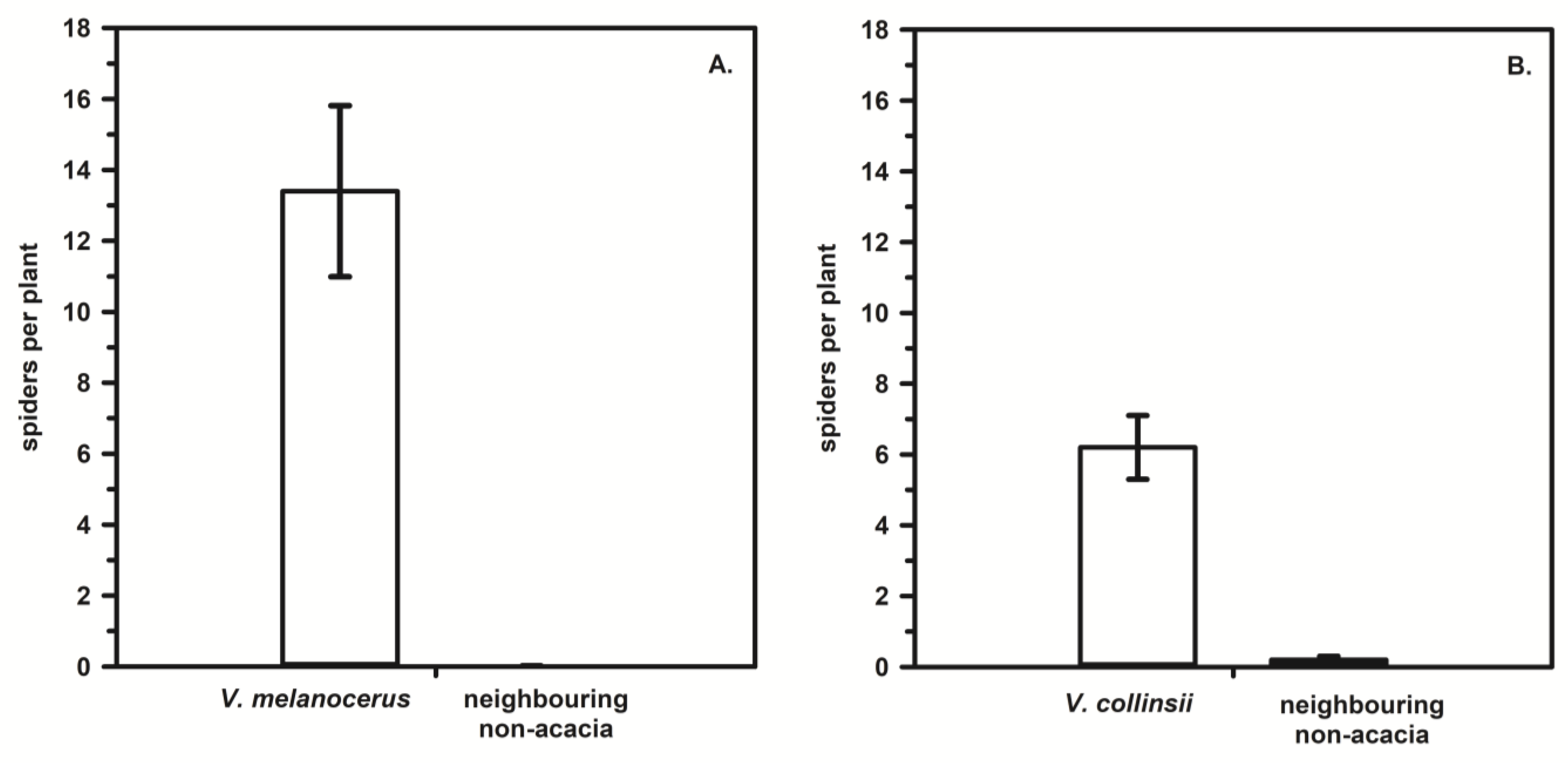
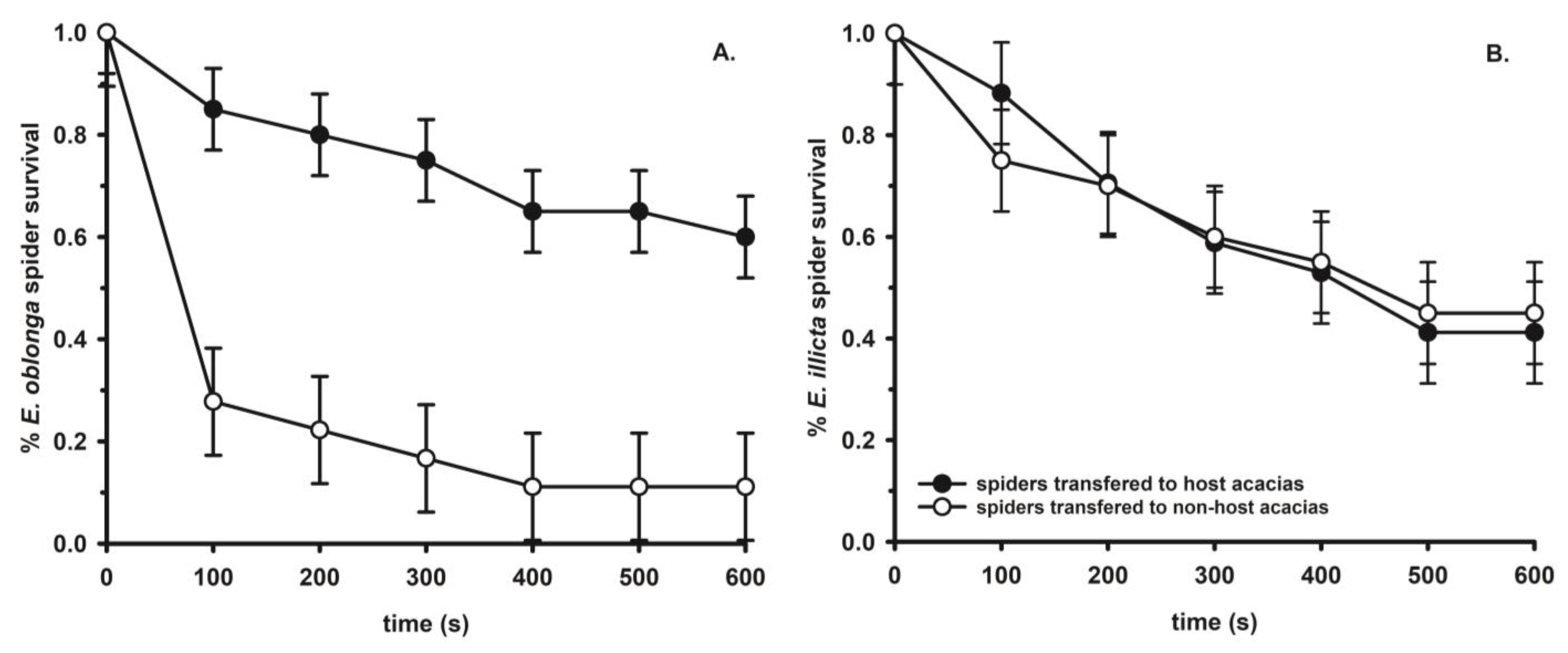
3. Host Plant Specificity in Web-Building Spiders: The Unique Cases of Eustala and Laetesia
3.1. The Araneid Orb-Web Spiders in the Genus Eustala
3.2. The Linyphiid Laetesia Raveni
4. How Might Spiders Identify and Locate Their Host Plant?
4.1. Website Choices and Web Building
4.2. The Use of Chemical Cues and Communication in Spiders
5. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Janzen, D.H. Coevolution of mutualism between ant and acacias in Central America. Evolution 1966, 20, 249–274. [Google Scholar] [CrossRef] [PubMed]
- Weiblen, G.D. How to be a fig wasp. Annu. Rev. Entomol. 2002, 47, 299–330. [Google Scholar] [CrossRef] [PubMed]
- Pauw, A. Floral syndromes accurately predict pollination by a specialized oil-collecting bee (Rediviva peringueyi, Melittidae) in a guild of South African orchids (Coryciinae). Reprod. Biol. 2006, 93, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Vacante, V. The Handbook of Mites of Economic Plants: Identification, Bio-Ecology and Control, 1st ed.; CAB International: Oxford, UK, 2016. [Google Scholar] [CrossRef]
- Gibson, C.W.; Hambler, C.; Brown, V.K. Changes in spider (Araneae) assemblages in relation to succession and grazing management. J. Appl. Ecol. 1992, 29, 132–142. [Google Scholar] [CrossRef]
- Patt, J.M.; Pfannenstiel, R.S. Odor-based recognition of nectar in cursorial spiders. Entomol. Exp. Appl. 2008, 127, 801–809. [Google Scholar] [CrossRef]
- Meehan, C.J.; Olson, E.J.; Reudink, M.W.; Kyser, T.K.; Curry, R.L. Herbivory in a spider through exploitation of an ant-plant mutualism. Curr. Biol. 2009, 19, 892–893. [Google Scholar] [CrossRef]
- Eggs, B.; Sanders, D. Herbivory in spiders: The importance of pollen for orb-weavers. PLoS ONE 2013, 8, e82637. [Google Scholar] [CrossRef]
- Vasconcellos-Neto, J.; Messas, Y.F.; Souza, H.D.S.; Villanueva-Bonilla, G.A.; Romero, G.Q. Spider-plant interactions: An ecological approach. In Behaviour and Ecology of Spiders: Contributions from the Neotropical Region, 1st ed.; Viera, C., Gonzaga, M.O., Eds.; Springer: Cham, Switzerland, 2017; pp. 165–214. [Google Scholar] [CrossRef]
- Hormiga, G.; Scharff, N. The strange case of Laetesia raveni n. sp., a green linyphiid spider from Eastern Australia with a preference for thorny plants (Araneae, Linyphiidae). Zootaxa 2014, 3811, 83–94. [Google Scholar] [CrossRef]
- Hesselberg, Y.; Triana, E. The web of the acacia orb-spider Eustala illicita (Araneae: Araneidae) with notes on its natural history. J. Arachnol. 2010, 38, 2126. [Google Scholar] [CrossRef]
- Garcia, L.C.; Styrsky, J.D. An orb-weaver spider eludes plant-defending acacia ants by hiding in plain sight. Ecol. Entomol. 2013, 28, 190–199. [Google Scholar] [CrossRef]
- Styrsky, J.D. An orb-weaver spider exploits an ant–acacia mutualism for enemy-free space. Ecol. Evol. 2014, 4, 276–283. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef]
- Wang, H.M.; Bai, P.H.; Zhang, J.; Zhang, X.M.; Hui, Q.; Zheng, H.X.; Zhang, X.H. Attraction of bruchid beetles Callosobruchus chinensis (L.) (Coleoptera: Bruchidae) to host plant volatiles. J. Integr. Agric. 2020, 19, 3035–3044. [Google Scholar] [CrossRef]
- Blatrix, R.; Mayer, V. Communication in ant-plant symbioses. In Plant Communication from an Ecological Perspective: Signaling and Communication in Plants, 1st ed.; Baluška, F., Ninkovic, V., Eds.; Springer: Heidelberg/Berlin, Germany, 2010. [Google Scholar]
- Dáttilo, W.F.C.; Izzo, T.J.; Inouye, B.D.; Vasconcelos, H.L.; Bruna, E.M. Recognition of host plant volatiles by Pheidole minutula Mayr (myrmicinae), an Amazonian ant-plant specialist. Biotropica 2009, 41, 642–646. [Google Scholar] [CrossRef]
- Inui, Y.; Itioka, T.; Murase, K.; Yamaoka, R.; Itino, T. Chemical recognition of partner plant species by foundress ant queens in Macaranga-Crematogaster myrmecophytism. J. Chem. Ecol. 2001, 27, 2029–2040. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A. Chemical communication in spiders—A methodological review. J. Arachnol. 2019, 47, 1–27. [Google Scholar] [CrossRef]
- Scott, C.; Gerak, C.; McCann, S.; Gries, G. The role of silk in courtship and chemical communication of the false widow spider, Steatoda grossa (Araneae: Theridiidae). J. Ethol. 2018, 36, 191–197. [Google Scholar] [CrossRef]
- Clark, R.J.; Jackson, R.R.; Cutler, B. Chemical cues from ants influence predatory behavior in Harbocestum pulex (Araneae, Salticidae). J. Arachnol. 2000, 28, 309318. [Google Scholar] [CrossRef]
- Krell, F.K.; Krämer, F. Chemical attraction of crab spiders (Araneae, Thomisidae) to a flower fragrance component. J. Arachnol. 1998, 26, 117–119. [Google Scholar]
- Rao, D. Habitat selection and dispersal. In Behaviour and Ecology of Spiders: Contributions from the Neotropical Region, 1st ed.; Viera, C., Gonzaga, M.O., Eds.; Springer: Cham, Switzerland, 2017; pp. 85–108. [Google Scholar] [CrossRef]
- Young, O.P.; Lockley, T.C. Spiders of Spanish moss in the Delta of Mississippi. J. Arachnol. 1989, 17, 143–148. [Google Scholar]
- Romero, G.Q.; Vasconcellos-Neto, J. Spatial distribution patterns of jumping spiders associated with terrestrial bromeliads. Biotropica 2004, 36, 596–601. [Google Scholar] [CrossRef]
- Romero, G.Q. Geographic range, habitats and host plants of bromeliad-living jumping spider (Salticidae). Biotropica 2006, 38, 522–530. [Google Scholar] [CrossRef]
- Romero, G.Q.; Vasconcellos-Neto, J. Foraging by the flower-dwelling spider, Misumenops argenteus (Thomisidae), at high prey density sites. J. Nat. Hist. 2004, 38, 1287–1296. [Google Scholar] [CrossRef]
- Romero, G.Q.; Vasconcellos-Neto, J. Spatial distribution and microhabitat preference of Psecas chapoda (Peckham & Peckham) (Araneae, Salticidae). J. Arachnol. 2005, 33, 124–134. [Google Scholar]
- Romero, G.Q.; Vasconcellos-Neto, J. Population dynamics, age structure and sex ratio of the bromeliad-dwelling jumping spider, Psecas chapoda (Salticidae). J. Nat. Hist. 2005, 39, 153–163. [Google Scholar] [CrossRef]
- Gonçalves, A.Z.; Mercier, H.; Mazzafera, P.; Romero, G.Q. Spider-fed bromeliads: Seasonal and interspecific variation in plant performance. Ann. Bot. 2011, 107, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Romero, G.Q.; Mazzafera, P.; Vasconcellos-Neto, J.; Trivelin, P.C.O. Bromeliad-living spiders improve host plant nutrition and growth. Ecology 2006, 87, 803–808. [Google Scholar] [CrossRef]
- Romero, G.Q.; Vasconcellos-Neto, J.; Trivelin, P.C.O. Spatial variation in the strength of mutualism between a jumping spider and a terrestrial bromeliad: Evidence from the stable isotope 15N. Acta Oecologica 2008, 33, 380–386. [Google Scholar] [CrossRef]
- Thorogood, C.J.; Bauer, U.; Hiscock, S.J. Convergent and divergent evolution in carnivorous pitcher plant traps. New Phytol. 2018, 217, 1035–1041. [Google Scholar] [CrossRef]
- Lim, R.J.Y.; Lam, W.N.; Tan, H.T.W. Novel pitcher plant–spider mutualism is dependent upon environmental resource abundance. Oecologia 2018, 188, 791–800. [Google Scholar] [CrossRef]
- Choo, J.P.S.; Koh, T.L.; Ng, P.K.L. Pitcher fluid macrofauna: Nematodes and arthropods. In A Guide to the Carnivorous Plants of Singapore, 1st ed.; Tan, H.T.W., Ed.; Singapore Science Centre: Singapore, 1997. [Google Scholar]
- Rembold, K.; Fischer, E.; Striffler, B.F.; Barthlott, W. Crab spider association with the Malagasy pitcher plant Nepenthes madagascariensis. Afr. J. Ecol. 2012, 51, 188–191. [Google Scholar] [CrossRef]
- Lam, W.N.; Tan, H.T.W. The crab spider–pitcher plant relationship is a nutritional mutualism that is dependent on prey-resource quality. J. Anim. Ecol. 2019, 88, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Anderson, B.; Midgely, J.J. It takes two to tango but three is a tangle: Mutualists and cheaters on the carnivorous plant Roridula. Oecologia 2002, 132, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Morais-Filho, J.C.; Romero, G.Q. Microhabitat use by Peucetia flava (Oxyopidae) on the glandular plant Rhyncanthera dichotoma (Melastomataceae). J. Arachnol. 2008, 36, 374–378. [Google Scholar] [CrossRef]
- Morais-Filho, J.C.; Romero, G.Q. Plant glandular trichomes mediate protective mutualism in a spider–plant system. Ecol. Entomol. 2010, 35, 485–494. [Google Scholar] [CrossRef]
- Gavini, S.S.; Quintero, C.; Tadey, M. Ecological role of a flower-dwelling predator in a tri-trophic interaction in northwestern Patagonia. Acta Oecologica 2019, 95, 100–107. [Google Scholar] [CrossRef]
- Vasconcellos-Neto, J.; Romero, G.Q.; Santos, A.J.; Dippenaar-Schoeman, A.S. Associations of spiders of the genus Peucetia (Oxyopidae) with plants bearing glandular hairs. Biotropica 2007, 39, 221–226. [Google Scholar] [CrossRef]
- Kessler, A.; Baldwin, I.T. Plant responses to insect herbivory: The emerging molecular analysis. Annu. Rev. Plant Biol. 2002, 53, 299–328. [Google Scholar] [CrossRef]
- Wagner, G.J.; Wang, E.; Shepherd, R.W. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot. 2004, 93, 3–11. [Google Scholar] [CrossRef]
- Sousa-Lopes, B.; Alves-da-Silva, N.; Alves-Martins, F.; Del-Claro, K. Antiherbivore protection and plant selection by the lynx spider Peucetia flava (Araneae: Oxyopidae) in the Brazilian Cerrado. J. Zool. 2019, 308, 121–127. [Google Scholar] [CrossRef]
- Romero, G.Q.; Souza, J.C.; Vasconcellos-Neto, J. Anti-herbivore protection by mutualistic spiders and the role of plant glandular trichomes. Ecology 2008, 89, 3105–3115. [Google Scholar] [CrossRef] [PubMed]
- Morais-Filho, J.C.; Romero, G.Q. Natural history of Peucetia flava (Araneae, Oxyopidae): Seasonal density fluctuation, phenology and sex ratio on the glandular plant Rhyncanthera dichotoma (Melastomataceae). J. Nat. Hist. 2009, 43, 701–711. [Google Scholar] [CrossRef]
- Gómez-Acevedo, S.L. New reports of spiders in three Mexican ant-acacias. Open J. Ecol. 2021, 11, 32–40. [Google Scholar] [CrossRef]
- Heiling, A.M.; Chittka, L.; Cheng, K.; Herberstein, M.E. Colouration in crab spiders: Substrate choice and prey attraction. J. Exp. Biol. 2005, 208, 1785–1792. [Google Scholar] [CrossRef]
- Llandres, A.L.; Gawryszewski, F.M.; Heiling, A.M.; Herberstein, M.E. The effect of colour variation in predators on the behaviour of pollinators: Australian crab spiders and native bees. Ecol. Entomol. 2011, 36, 72–81. [Google Scholar] [CrossRef]
- Théry, M.; Debut, M.; Gomez, D.; Casas, J. Specific color sensitivities of prey and predator explain camouflage in different visual systems. Behav. Ecol. 2005, 16, 25–29. [Google Scholar] [CrossRef]
- Su, Q.; Qi, L.; Zhang, W.; Yun, Y.; Zhao, Y.; Peng, Y. Biodiversity survey of flower-visiting spiders based on literature review and field study. Environ. Entomol. 2020, 49, 673–682. [Google Scholar] [CrossRef]
- Brechbühl, R.; Casas, J.; Bacher, S. Ineffective crypsis in a crab spider: A prey community perspective. Proc. Roy. Soc. B 2009, 277, 739–746. [Google Scholar] [CrossRef]
- Rose, C.; Schramm, A.; Irish, J.; Bilde, T.; Bird, T.L. Host plant availability and nest-site selection of the social spider Stegodyphus dumicola Pocock, 1898 (Eresidae). Insects 2022, 13, 30. [Google Scholar] [CrossRef]
- Romero, G.Q.; Vasconcellos-Neto, J. The effects of plant structure on the spatial and microspatial distribution of a bromeliad-living jumping spider (Salticidae). J. Anim. Ecol. 2005, 74, 12–21. [Google Scholar] [CrossRef]
- Omena, P.M.; Romero, G.Q. Fine-scale microhabitat selection in a bromeliad-dwelling jumping spider (Salticidae). Biol. J. Linn. Soc. 2008, 94, 653–662. [Google Scholar] [CrossRef]
- Jacobucci, G.B.; Medeiros, L.; Vasconcellos-Neto, J.; Romero, G.Q. Habitat selection and potential antiherbivore effects of Peucetia flava (Oxyopidae) on Solanum thomasiifolium (Solanaceae). J. Arachnol. 2009, 37, 365–367. [Google Scholar] [CrossRef]
- Uetz, G.W. Habitat Structure and Spider Foraging. In Habitat Structure, 1st ed.; Bell, S.S., McCoy, E.D., Mushinsky, H.R., Eds.; Springer: Dordrecht, Switzerland, 1991; pp. 325–348. [Google Scholar] [CrossRef]
- Wise, D.H. Spiders in Ecological Webs, 1st ed.; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar] [CrossRef]
- Souza, A.L.T.D.; Martins, R.P. Distribution of plant-dwelling spiders: Inflorescences versus vegetative branches. Austral Ecol. 2004, 29, 342–349. [Google Scholar] [CrossRef]
- Villanueva-Bonilla, G.A.; Salomão, A.T.; Vasconcellos-Neto, J. Trunk structural traits explain habitat use of a tree-dwelling spider (Selenopidae ) in a tropical forest. Acta Oecologica 2017, 85, 108–115. [Google Scholar] [CrossRef]
- Arango, A.M.; Rico-Gray, V.; Parra-Tabla, V. Population structure, seasonality, and habitat use by the green lynx spider Peucetia viridans (Oxyopidae) inhabiting Cnidoscolus aconitifolius (Euphorbiaceae). J. Arachnol. 2000, 28, 185–194. [Google Scholar] [CrossRef]
- Corcuera, P.; Valverde, P.L.; Jiménez-Salinas, E.; Vite, F.; López-Ortega, G.; Pérez-Hernández, M.A. Distribution of Peucetia viridans (Araneae: Oxyopidae) on Croton ciliatoglandulifer. Environ. Entomol. 2010, 39, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Cross, F.R. Attentional Processes in Mosquito-Eating Jumping Spiders: Search Images and Cross-Modality Priming. Ph.D. Dissertation, University of Canterbury, Canterbury, New Zealand, 2009. [Google Scholar]
- Agnarsson, I.; Rayor, L.S. A molecular phylogeny of the Australian huntsman spiders (Sparassidae, Deleninae): Implications for taxonomy and social behaviour. Mol. Phyl. Evol. 2013, 69, 895–905. [Google Scholar] [CrossRef]
- Quintero, C.; Corley, J.C.; Aizen, M.A. Weak trophic links between a crab-spider and the effective pollinators of a rewardless orchid. Acta Oecologia 2015, 62, 32–39. [Google Scholar] [CrossRef]
- Karl, I.; Bauer, U. Inside the trap: Biology and behavior of the pitcher-dwelling crab spider, Misumenops nepenthicola. Plants People Planet 2020, 2, 290–293. [Google Scholar] [CrossRef]
- Anderson, B. Inferring evolutionary patterns from the biogeographical distributions of mutualists and exploiters. Biol. J. Linn. Soc. 2006, 89, 541–549. [Google Scholar] [CrossRef]
- Tanaka, K. Energetic cost of web construction and its effect on web relocation in the web-building spider Agelena limbata. Oecologia 1989, 81, 459–464. [Google Scholar] [CrossRef] [PubMed]
- Chmiel, K.; Herberstein, M.E.; Elgar, M.A. Web damage and feeding experience influence web site tenacity in the orb-web spider Argiope keyserlingi Karsch. Anim. Behav. 2000, 60, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Whitney, K.D. Experimental evidence that both parties benefit in a facultative plant-spider mutualism. Ecology 2004, 85, 1642–1650. [Google Scholar] [CrossRef]
- Ruch, J.; Heinrich, L.; Bilde, T.; Schneider, J.M. Site selection and foraging in the eresid spider Stegodyphus tentoriicola. J. Insect Behav. 2012, 25, 1–11. [Google Scholar] [CrossRef]
- Nishi, A.H.; Vasconcellos-Neto, J.; Romero, G.Q. The role of multiple partners in a digestive mutualism with a protocarnivorous plant. Ann. Bot. 2013, 111, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.E.; Lohmiller, J.; Joern, A. Importance of vegetation structure to the assembly of an aerial web-building spider community in North American open grassland. J. Arachnol. 2016, 44, 28–35. [Google Scholar] [CrossRef]
- Bomfim, L.D.S.; Bitencourt, J.A.G.; Rodrigues, E.N.L.; Podgaiski, L.R. The role of a rosette-shaped plant (Eryngium horridum, Apiaceae) on grassland spiders along a grazing intensity gradient. Insect Conserv. Divers 2021, 14, 492–503. [Google Scholar] [CrossRef]
- Jolivet, P.; Vasconcellos-Neto, J. Convergence chez les plantes carnivores. La Rech. 1993, 24, 456–458. [Google Scholar]
- Cresswell, J.E. Capture rates and composition of insect prey of the pitcher plant Sarracenia purpurea. Am. Midl. Nat. 1991, 125, 1–9. [Google Scholar] [CrossRef]
- Milne, M.A.; Waller, D.A. Does pitcher plant morphology affect spider residency? Northeast. Nat. 2013, 20, 419–429. [Google Scholar] [CrossRef]
- Fage, L. Au sujet de deux araignées nouvelles trouvées dans les urnes de Nepenthes. Treubia 1930, 12, 23–28. [Google Scholar]
- Lubin, Y.; Hennicke, J.; Schneider, J. Settling decisions of dispersing Stegodyphus lineatus (Eresidae) young. Isr. J. Ecol. Evol. 1998, 44, 217–225. [Google Scholar]
- Cuff, J.P.; Evans, S.A.; Porteous, I.A.; Quiñonez, J.; Evans, D.M. Candy-striped spider leaf and habitat preferences for egg deposition. Agric. For. Entomol. 2022, 24, 422–431. [Google Scholar] [CrossRef]
- Chickering, A.M. The genus Eustala (Araneae, Argiopidae) in Central America. Bull. Mus. Comp. Zool. 1955, 112, 391–518. [Google Scholar]
- Poeta, M.R.M. The orb-weaving spider genus Eustala Simon, 1895 (Araneae, Araneidae): Eight new species, redescriptions, and new records. Zootaxa 2014, 3872, 440–446. [Google Scholar] [CrossRef] [PubMed]
- World Spider Catalog: NMBE. Available online: http://wsc.nmbe.ch (accessed on 29 October 2022).
- Comstock, J.H. The Spider Book, 1st ed.; Doubleday, Page and Company: New York, NY, USA, 1912. [Google Scholar]
- Levi, H.W. The American orb-weaver genera Cyclosa, Metazygia and Eustala north of Mexico (Araneae, Araneidae). Bull. Mus. Comp. Zool. 1997, 148, 61–127. [Google Scholar]
- Messas, Y.F.; Souza, H.S.; Gonzaga, M.O.; Vasconcellos-Neto, J. Spatial distribution and substrate selection by the orb-weaver spider Eustala perfida Mello-Leitão, 1947 (Araneae: Araneidae). J. Nat. Hist. 2014, 48, 2645–2660. [Google Scholar] [CrossRef]
- Souza, H.S.; Messas, Y.F.; Gonzaga, M.O.; Vasconcellos-Neto, J. Substrate selection and spatial segregation by two congeneric species of Eustala (Araneae: Araneidae) in southeastern Brazil. J. Arachnol. 2015, 43, 59–66. [Google Scholar] [CrossRef]
- Ledin, A.E.; Styrsky, J.D.; Styrsky, J.N. Friend or foe? orb-weaver spiders inhabiting ant-acacias capture both herbivorous insects and acacia ant alates. J. Insect Sci. 2020, 20, 1–8. [Google Scholar] [CrossRef]
- Cushing, P.E. Spider-ant associations: An updated review of myrmecomorphy, myrmecophily, and myrmecophagy in spiders. Psyche 2012, 2012, 151989. [Google Scholar] [CrossRef]
- Eberhard, W.G. Aggressive chemical mimicry by a bolas spider. Science 1977, 198, 1173–1175. [Google Scholar] [CrossRef] [PubMed]
- Nishi, A.H. Mutualismo Digestivo Entre Aranhas, Cupins e a Planta Protocarnívora Paepalanthus bromelioides (Eriocaulaceae). Master’s Thesis, São Paulo State University (Universidade Estadual Paulista), São Paulo, Brazil, 2011. [Google Scholar]
- Gastreich, K.R. Trait-mediated indirect effects of a theridiid spider on an ant-plant mutualism. Ecology 1999, 80, 1066–1070. [Google Scholar] [CrossRef]
- Zschokke, S.; Vollrath, F. Unfreezing the behavior of two orb spiders. Physiol. Behav. 1995, 58, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, S.P.; Zschokke, S. Webs of theridiid spiders: Construction, structure and evolution. Biol. J. Linn. Soc. 2003, 78, 293–305. [Google Scholar] [CrossRef]
- Vollrath, F. Analysis and interpretation of orb spider exploration and web-building behavior. Adv. Study. Behav. 1992, 21, 147–199. [Google Scholar] [CrossRef]
- Hesselberg, T. Exploration behaviour and behavioural flexibility in orb-web spiders: A review. Curr. Zool. 2015, 61, 313–327. [Google Scholar] [CrossRef]
- Zschokke, S. Early stages of orb web construction in Araneus diadematus Clerck. Rev. Suisse. Zool. 1996, 2, 709–720. [Google Scholar]
- Hesselberg, T. Web-building flexibility differs in two spatially constrained orb spiders. J. Insect Behav. 2013, 26, 283–303. [Google Scholar] [CrossRef]
- Eberhard, W.G. Behavioral flexibility in orb web construction: Effects of supplies in different silk glands and spider size and weight. J. Arachnol. 1988, 16, 295–302. [Google Scholar]
- Craig, C.L. The ecological and evolutionary interdependence between web architecture and web silk spun by orb web weaving spiders. Biol. J. Linn. Soc. 1987, 30, 135–162. [Google Scholar] [CrossRef]
- Ulrich, J.C. Wind effects of web structure of grass spider Agelenopsis actuosa. Arachnology 2021, 18, 993–997. [Google Scholar] [CrossRef]
- Mulder, T.; Wilkins, L.; Mortimer, B.; Vollrath, F. Dynamic environments do not appear to constrain spider web building behaviour. Sci. Nat. 2021, 108, 20. [Google Scholar] [CrossRef] [PubMed]
- Gunnarsson, B. Fractal dimension of plants and body size distribution in spiders. Fun. Ecol. 1992, 6, 636–641. [Google Scholar] [CrossRef]
- Wolff, J.O.; Little, D.; Herberstein, M.E. Limits of piriform silk adhesion-similar effects of substrate surface polarity on silk anchor performance in two spider species with disparate microhabitat use. Sci. Nat. 2020, 107, 31. [Google Scholar] [CrossRef] [PubMed]
- Hesselberg, T. Ontogenetic changes in web design in two orb-web spiders. Ethology 2010, 116, 535–545. [Google Scholar] [CrossRef]
- Hesselberg, T. The mechanism behind plasticity of web-building behavior in an orb spider facing spatial constraints. J. Arachnol. 2014, 42, 311–314. [Google Scholar] [CrossRef]
- Foelix, R. Biology of Spiders, 1st ed.; Oxford University Press: Oxford, UK, 2011. [Google Scholar]
- Morley, E.L.; Robert, D. Electric fields elicit ballooning in spiders. Curr. Biol. 2018, 28, 2324–2330.e2. [Google Scholar] [CrossRef]
- Cho, M. Aerodynamics and the role of the earth’s electric field in the spiders’ ballooning flight. J. Comp. Physiol. A 2021, 207, 219–236. [Google Scholar] [CrossRef]
- Duffey, E. Aerial dispersal in a known spider population. J. Anim. Ecol. 1956, 25, 85–111. [Google Scholar] [CrossRef]
- Lee, V.M.; Kuntner, M.; Li, D. Ballooning behavior in the golden orbweb spider Nephila pilipes (Araneae: Nephilidae). Front. Ecol. Evol. 2015, 3, 2. [Google Scholar] [CrossRef]
- Weyman, G.S.; Sunderland, K.D.; Jepson, P.C. A review of the evolution and mechanisms of ballooning by spiders inhabiting arable farmland. Ethol. Ecol. Evol. 2002, 14, 307–326. [Google Scholar] [CrossRef]
- Mowery, M.A.; Lubin, Y.; Segoli, M. Invasive brown widow spiders disperse aerially under a broad range of environmental conditions. Ethology 2022, 128, 564–571. [Google Scholar] [CrossRef]
- Larrivée, M.; Buddle, C.M. Diversity of canopy and understorey spiders in north-temperate hardwood forests. Agric. For. Entomol. 2009, 11, 225–237. [Google Scholar] [CrossRef]
- Ruhland, F.; Chiara, V.; Trabalon, M. Age and egg-sac loss determine maternal behaviour and locomotor activity of wolf spiders (Araneae, Lycosidae). Behav. Process. 2016, 132, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Sergi, C.; Schlais, A.; Marshall, M.; Rodríguez, R.L. Western black widow spiders (Latrodectus hesperus) remember prey capture location and size, but only alter behavior for prey caught at particular sites. Ethology 2022, 128, 707–714. [Google Scholar] [CrossRef]
- Rodríguez, R.L.; Gloudeman, M.D. Estimating the repeatability of memories of captured prey formed by Frontinella communis spiders (Araneae: Linyphiidae). Anim. Cogn. 2011, 14, 675–682. [Google Scholar] [CrossRef]
- Nørgaard, T.; Henschel, J.R.; Wehner, R. Long-distance navigation in the wandering desert spider Leucorchestris arenicola: Can the slope of the dune surface provide a compass cue? J. Comp. Physiol. A 2003, 189, 801–809. [Google Scholar] [CrossRef]
- Gaffin, D.D.; Curry, C.M. Arachnid navigation—A review of classic and emerging models. J. Arachnol. 2020, 48, 1–25. [Google Scholar] [CrossRef]
- Wherry, T.; Elwood, R.W. Relocation, reproduction and remaining alive in an orb-web spider. J. Zool. 2009, 279, 57–63. [Google Scholar] [CrossRef]
- Gillespie, R. The mechanism of habitat selection in the long-jawed orb-weaving spider Tetragnatha elongata (Araneae, Tetragnathidae). J. Arachnol. 1987, 15, 81–90. [Google Scholar]
- Beyer, M.; Mangliers, J.; Tuni, C. Silk-borne chemicals of spider nuptial gifts elicit female gift acceptance. Biol. Lett. 2021, 17, 20210386. [Google Scholar] [CrossRef]
- Gerbaulet, M.; Möllerke, A.; Weiss, K.; Chinta, S.; Schneider, J.M.; Schulz, S. Identification of cuticular and web lipids of the spider Argiope bruennichi. J. Chem. Ecol. 2022, 48, 244–262. [Google Scholar] [CrossRef]
- Fischer, A.; Lee, Y.; Dong, T.; Gries, G. Know your foe: Synanthropic spiders are deterred by semiochemicals of European fire ants. R. Soc. Open Sci. 2021, 8, 210279. [Google Scholar] [CrossRef] [PubMed]
- Rypstra, A.L.; Schmidt, J.M.; Reif, B.D.; DeVito, J.; Persons, M.H. Tradeoffs involved in site selection and foraging in a wolf spider: Effects of substrate structure and predation risk. Oikos 2007, 116, 853–863. [Google Scholar] [CrossRef]
- Penfold, S.; Dayananda, B.; Webb, J.K. Chemical cues influence retreat-site selection by flat rock spiders. Behaviour 2017, 154, 149–161. [Google Scholar] [CrossRef]
- Dettner, K.; Liepert, C. Chemical mimicry and camouflage. Annu. Rev. Entomol. 1994, 39, 129–154. [Google Scholar] [CrossRef]
- Allan, R.A.; Capon, R.J.; Brown, W.V.; Elgar, M.A. Mimicry of host cuticular hydrocarbons by salticid spider Cosmophasis bitaeniata that preys on larvae of tree ants Oecophylla smaragdina. J. Chem. Ecol. 2002, 28, 835–848. [Google Scholar] [CrossRef]
- Elgar, M.A.; Allan, R.A. Predatory spider mimics acquire colony-specific cuticular hydrocarbons from their ant model prey. Naturwissenschaften 2004, 91, 143–147. [Google Scholar] [CrossRef]
- Cerveira, A.M.; Jackson, R.R. Inter-population variation and phenotypic plasticity in kairomone use by a poly-specialist spider-eating predator. J. Ethol. 2022, 40, 37–48. [Google Scholar] [CrossRef]
- Pekár, S. Ant-mimicking spider actively selects its mimetic model (Araneae: Gnaphosidae; Hymenoptera: Formicidae). Myrmecol. News. 2020, 30, 131–137. [Google Scholar] [CrossRef]
- Johnson, J.C.; Trubl, P.; Blackmore, V.; Miles, L. Male black widows court well-fed females more than starved females: Silken cues indicate sexual cannibalism risk. Anim. Behav. 2011, 82, 383–390. [Google Scholar] [CrossRef]
- Junker, R.R.; Bretscher, S.; Dötterl, S.; Blüthgen, N. Phytochemical cues affect hunting-site choices of a nursery web spider (Pisaura mirabilis) but not a crab spider (Misumena vatia). J. Arachnol. 2011, 39, 113–117. [Google Scholar] [CrossRef]
- Heiling, A.M.; Cheng, K.; Herberstein, M.E. Exploitation of floral signals by crab spiders (Thomisus spectabilis, Thomisidae). Behav. Ecol. 2004, 15, 321–326. [Google Scholar] [CrossRef]
- Tedore, C.; Johnsen, S. Immunological dependence of plant-dwelling animals on the medicinal properties of their plant substrates: A preliminary test of a novel evolutionary hypothesis. Arthropod. Plant Interact. 2015, 9, 437–446. [Google Scholar] [CrossRef]
- Li, Z.; Su, Q.; Zhao, Y.; Yun, Y.; Peng, Y. Volatile scents preference of crab spiders (Ebrechtella tricuspidata) for host flowers. ResearchSquare PPR482028. arXiv 2022. [Google Scholar] [CrossRef]
- Blamires, S.J. Plasticity in extended phenotypes: Orb web architectural responses to variations in prey parameters. J. Exp. Biol. 2010, 213, 3207–3212. [Google Scholar] [CrossRef]
- Haberkern, A.M.; Fernandez-Fournier, P.; Avilés, L. Spinning in the rain: Interactions between spider web morphology and microhabitat use. Biotropica 2020, 52, 480–487. [Google Scholar] [CrossRef]
- Hironaka, Y.; Abé, H. Nesting habits of the Japanese foliage spider, Cheiracanthium japonicum (Araneae: Miturgidae): Host plant preference based on the physical traits of plant leaves. J. Nat. Hist. 2012, 46, 2665–2676. [Google Scholar] [CrossRef]
- Itescu, Y. Are island-like systems biologically similar to islands? a review of the evidence. Ecography 2018, 42, 1298–1314. [Google Scholar] [CrossRef]
- Slatkin, M. Gene flow and the geographic structure of natural populations. Science 1987, 236, 787–792. [Google Scholar] [CrossRef]
- Lee, J.W.; Jiang, L.; Su, Y.C.; Tso, I.M. Is Central mountain range a geographic barrier to the giant wood spider Nephila pilipes (Araneae: Tetragnathidae) in Taiwan? a population genetic approach. Zool. Stud. 2004, 43, 112–122. [Google Scholar]
- Bell, J.R.; Bohan, D.A.; Shaw, E.M.; Weyman, G.S. Ballooning dispersal using silk: World fauna, phylogenies, genetics and models. Bull. Entomol. Res. 2005, 95, 69–114. [Google Scholar] [CrossRef] [PubMed]
- Amador-Vargas, S.; Orribarra, V.S.; Portugal-Loayza, A.; Fernández-Marín, H. Association patterns of swollen-thorn acacias with three ant species and other organisms in a dry forest of Panama. Biotropica 2021, 53, 560–566. [Google Scholar] [CrossRef]
- Coronado-Rivera, J.; Del Valle, M.S.; Amador-Vargas, S. True bugs living on ant-defended acacias: Evasion strategies and ant species preferences, in Costa Rica and Panama. Rev. Biol. Trop. 2020, 68, 415–425. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hesselberg, T.; Boyd, K.M.; Styrsky, J.D.; Gálvez, D. Host Plant Specificity in Web-Building Spiders. Insects 2023, 14, 229. https://doi.org/10.3390/insects14030229
Hesselberg T, Boyd KM, Styrsky JD, Gálvez D. Host Plant Specificity in Web-Building Spiders. Insects. 2023; 14(3):229. https://doi.org/10.3390/insects14030229
Chicago/Turabian StyleHesselberg, Thomas, Kieran M. Boyd, John D. Styrsky, and Dumas Gálvez. 2023. "Host Plant Specificity in Web-Building Spiders" Insects 14, no. 3: 229. https://doi.org/10.3390/insects14030229
APA StyleHesselberg, T., Boyd, K. M., Styrsky, J. D., & Gálvez, D. (2023). Host Plant Specificity in Web-Building Spiders. Insects, 14(3), 229. https://doi.org/10.3390/insects14030229