
Behavioural and Electrophysiological Response of Anastrepha fraterculus (Diptera: Tephritidae) to a γ-Lactone Synthetic Semiochemical

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Individuals
2.2. Synthetic Semiochemicals
2.3. Experimental Procedures
2.3.1. Electroantennography
2.3.2. Field Cage Experiments
2.4. Data Analysis
2.4.1. Electroantennography
2.4.2. Field Cage Experiments
3. Results
3.1. Electroantennography
3.2. Field Cage Experiments
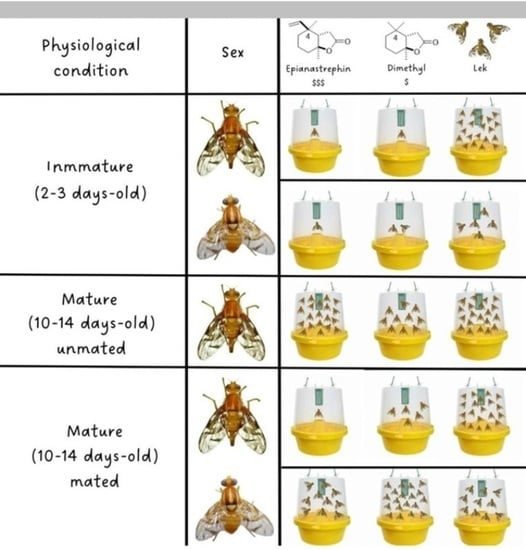
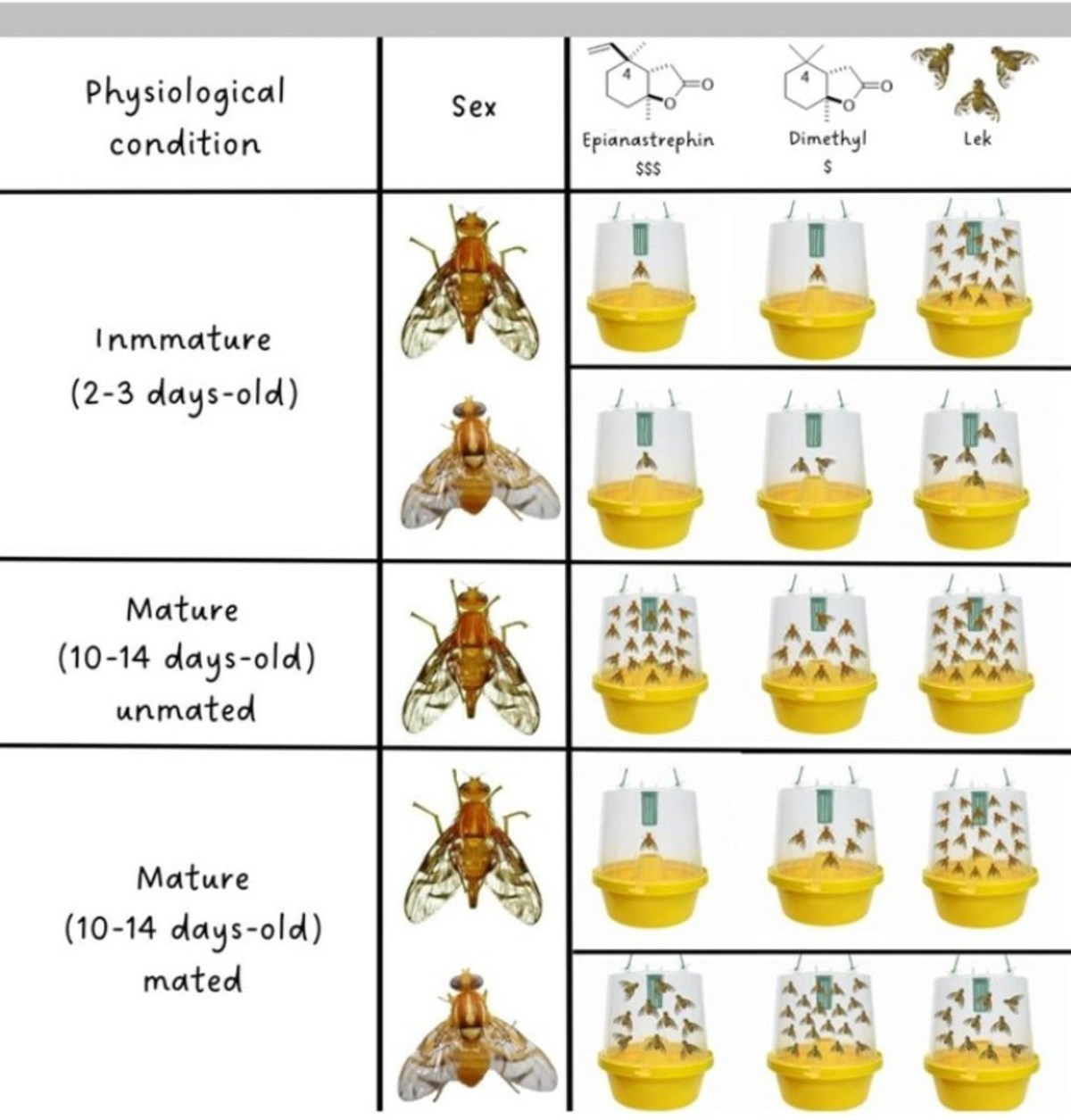
3.2.1. Immature Females and Males
3.2.2. Virgin Females
3.2.3. Mated Females and Males
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gadenne, C.; Barrozo, R.B.; Anton, S. Plasticity in insect olfaction: To smell or not to smell? Ann. Rev. Entomol. 2016, 61, 317–333. [Google Scholar] [CrossRef]
- Epsky, N.D.; Heath, R.R. Pheromone production by male Anastrepha suspensa (Diptera: Tephritidae) under natural light cycles in greenhouse studies. Environ. Entomol. 1993, 22, 464–469. [Google Scholar] [CrossRef]
- Heath, R.R.; Epsky, N.D.; Guzman, A.; Dueben, B.D.; Manukian, A.; Meyer, W.L. Development of a dry plastic insect trap with food-based synthetic attractant for the Mediterranean and Mexican fruit flies (Diptera: Tephritidae). J. Econ. Entomol. 1995, 88, 1307–1315. [Google Scholar] [CrossRef]
- Canal, N.A.; Chacon, M.D.; Galeano, P.E.; Gomez, S.G.; Gomez, M.J.; Ramirez, J.H.; Osorio, A.; Castaneda, M.R. Response of Anastrepha species (Diptera: Tephritidae) to synthetic attractants in Colombia. In IAEA-TECDOC, Development of Improved Attractants and Their Integration into Fruit Fly SIT Management Programmes; IAEA: Vienna, Austria, 2007; Volume 1574, pp. 129–141. [Google Scholar]
- Vargas, R.I.; Haviland, D.; Faber, B.; Kabashima, J.; Grafton-Cardwell, B.; Morse, J.G. Improving trapping systems for early detection and eradication of fruit flies in California. Citrograph 2013, 4, 28–34. [Google Scholar]
- Lima-Mendonça, A.; de Lima Mendonça, A.; Goulart Sant’Ana, A.; Do Nascimento, R. Semioquimicos de moscas das frutas do género Anastrepha. Química Nova 2014, 37, 293–301. [Google Scholar]
- Epsky, N.D.; Kendra, P.E.; Schnell, E.Q. History and development of food-based attractants. In Trapping and the Detection, Control and Regulation of Tephritid Fruit Flies; Shelly, T.E., Epsky, N., Jang, E.B., Reyes-Flores, J., Vargas, R.I., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 75–118. [Google Scholar]
- Levi-Zada, A.; Byers, J.A. Circadian rhythms of insect pheromone titer, calling, emission, and response: A review. Sci. Nat. 2021, 108, 35. [Google Scholar] [CrossRef] [PubMed]
- Scolari, F.; Valerio, F.; Benelli, G.; Papadopoulos, N.T.; Vaníčková, L. Tephritid Fruit Fly Semiochemicals: Current Knowledge and Future Perspectives. Insects 2021, 12, 408. [Google Scholar] [CrossRef]
- International Atomic Energy Agency. Trapping Guidelines for Area-Wide Fruit Fly Programmes; IAEA: Vienna, Austria, 2003. [Google Scholar]
- Beroza, M.; Green, N.; Gertler, S.; Steiner, L.; Miyashita, D. Insect attractants, new attractants for the Mediterranean fruit fly. J. Agric. Food Chem. 1961, 9, 361–365. [Google Scholar] [CrossRef]
- Vargas, R.I.; Stark, J.D.; Kido, M.H.; Ketter, H.M.; Whitehand, L.C. Methyl eugenol and cue-lure traps for suppression of male oriental fruit flies and melon flies (Diptera: Tephritidae) in Hawaii: Effects of lure mixtures and weathering. J. Econ. Entomol. 2000, 93, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.H.; Nishida, R.; Jang, E.B.; Shelly, T.E. Pheromones, male lures, and trapping of tephritid fruit flies. In Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies; Shelly, T.E., Epsky, N., Jang, E.B., Reyes-Flores, J., Vargas, R.I., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 15–74. [Google Scholar]
- Tasnin, S.; Merkel, K.; Clarke, A.R. Effects of advanced age on olfactory response of male and female Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae). J. Insect Physiol. 2020, 122, 104024. [Google Scholar] [CrossRef]
- Walse, S.S.; Kuzmich, D. Diastereoselective Synthesis of (±)- Epianastrephin, (±)-Anastrephin and Analogs Thereof. U.S. Patent No. 10,011,580, 3 July 2018. [Google Scholar]
- Vlad, P.F.; Gorincioi, E.C.; Aricu, A.N.; Coltsa, M.N. A Short, Efficient Synthesis of (-)(3as, 7as)-Trans-Tetrahydroactinidiolide. Nat. Prod. Lett. 1999, 13, 1–4. [Google Scholar] [CrossRef]
- Kaneko, H.; Hoshino, K. Isolation from Cigar Tobacco Leaves of Tetrahydroactinidiolide: (2-Hydroxy-2, 6, 6-trimethylcyclohexylacetic Acid γ-Lactone). Agric. Biol. Chem. 1969, 33, 969–970. [Google Scholar] [CrossRef]
- Milet-Pinheiro, P.; Navarro, D.; De Aquino, N.C.; Ferreira, L.L.; Tavares, R.F.; da Silva, R.D.C.C.; Lima Mendoca, A.; Do Nascimento, R. Identification of male-borne attractants in Anastrepha fraterculus (Diptera:Tephritidae). Chemoecology 2014, 25, 115–122. [Google Scholar] [CrossRef]
- Da Rosa, J.M.; Arioli, C.J.; Dos Santos, J.P.; Menezes-Netto, A.C.; Botton, M. Evaluation of food lures for capture and monitoring of Anastrepha fraterculus (Diptera: Tephritidae) on temperate fruit trees. J. Econom. Entomol. 2017, 110, 995–1001. [Google Scholar] [CrossRef]
- De Meyer, M.D.; Clarke, A.R.; Vera, M.T.; Hendrichs, J. Resolution of cryptic species complexes of tephritid pests to enhance SIT application and facilitate international trade. ZooKeys 2015, 540, 1–557. [Google Scholar] [CrossRef] [PubMed]
- Selivon, D.; Perondini, A.L.P.; Hernández-Ortiz, V.; doVal, F.C.; Camacho, A.; Gomes, F.R.; Prezotto, L.F. Genetical, Morphological, Behavioral, and Ecological Traits Support the Existence of Three Brazilian Species of the Anastrepha fraterculus Complex of Cryptic Species. Front. Ecol. Evol. 2022, 10, 359. [Google Scholar] [CrossRef]
- Malavasi, A.; Morgante, J.S.; Prokopy, R.J. Distribution and activities of Anastrepha fraterculus (Diptera: Tephritidae) flies on host and nonhost trees. Ann. Entomol. Soc. Am. 1983, 76, 286–292. [Google Scholar] [CrossRef]
- Cladera, J.L.; Vilardi, J.C.; Juri, M.; Paulin, L.E.; Giardini, M.C.; Gómez Cendra, P.V.; Segura, D.F.; Lanzavecchia, S.B. Genetics and biology of Anastrepha fraterculus: Research supporting the use of the sterile insect technique (SIT) to control this pest in Argentina. BMC Genet. 2014, 15, S12. [Google Scholar] [CrossRef]
- Guillén, D.; Sánchez, R. Expansion of the national fruit fly control programme in Argentina. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007. [Google Scholar]
- Hernández-Ortiz, V.; Bartolucci, A.F.; Morales-Valles, P.; Frías, D.; Selivon, D. Cryptic species of the Anastrepha fraterculus complex (Diptera: Tephritidae): A multivariate approach for the recognition of South American morphotypes. Ann. Entomol. Soc. Am. 2012, 105, 305–318. [Google Scholar] [CrossRef]
- Hernández-Ortiz, V.; Canal, N.A.; Tigrero Salas, J.O.; Ruiz-Hurtado, F.M.; Dzul-Cauich, J.F. Taxonomy and phenotypic relationships of the Anastrepha fraterculus complex in the mesoamerican and pacific neotropical dominions (Diptera, Tephritidae). ZooKeys 2015, 540, 95–124. [Google Scholar] [CrossRef]
- Norrbom, A. Host plant database for Anastrepha and Toxotrypana (Diptera: Tephritidae: Toxotrypanini). In Diptera Data Dissemination Disk (CD-ROM); USDAAPHIS: Washington, DC, USA, 2004; p. 2. [Google Scholar]
- Hernández-Ortiz, B.J.; Barradas-Juanz, N.; Díaz-Castelazo, C. A Review of the Natural Host Plants of the Anastrepha fraterculus Complex in the Americas. In Area-Wide Management of Fruit Fly Pests; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2020; pp. 90–114. [Google Scholar]
- Segura, D.F.; Petit-Marty, N.; Sciurano, R.B.; Vera, M.T.; Calcagno, G.; Allinghi, A.; Cendra, P.G.; Cladera, J.; Vilardi, J. Lekking behavior of Anastrepha fraterculus (Diptera: Tephritidae). Fla. Entomol. 2007, 90, 154–162. [Google Scholar] [CrossRef]
- Lima, I.S.; House, P.E.; Nascimento, R.R.D. Volatile substances from male Anastrepha fraterculus Wied. (Diptera: Tephritidae): Identification and behavioural activity. J. Braz. Chem. Soc. 2001, 12, 196–201. [Google Scholar] [CrossRef]
- Cáceres, C.; Segura, D.F.; Vera, M.T.; Wornoayporn, V.; Cladera, J.L.; Teal, P.; Sapountzis, P.; Bowtzis, K.; Zacharopoulou, A.; Robinson, A.S. Incipient speciation revealed in Anastrepha fraterculus (Diptera; Tephritidae) by studies on mating compatibility, sex pheromones, hybridization, and cytology. Biol. J. Linn. Soc. 2009, 97, 152. [Google Scholar] [CrossRef]
- Břízová, R.; Mendonça, A.L.; Vaníčková, L.; Mendonça, A.L.; Da Silva, C.E.; Tomčala, A.; Paranhos, B.A.J.; Dias, V.S.; Joachim-Bravo, I.S.; Hoskovec, M.; et al. Pheromone analyses of the Anastrepha fraterculus (Diptera: Tephritidae) cryptic species complex. Fla. Entomol. 2013, 96, 1107–1115. [Google Scholar] [CrossRef]
- Vaníčková, L.; Břízová, R.; Pompeiano, A.; Ferreira, L.L.; de Aquino, N.C.; de Farias Tavares, R.; Rodriguez, L.D.; Mendonça, A.D.L.; Canal, N.A.; Nascimento, R.R.D. Characterisation of the chemical profiles of Brazilian and Andean morphotypes belonging to the Anastrepha fraterculus complex (Diptera, Tephritidae). ZooKeys 2015, 540, 193. [Google Scholar] [CrossRef]
- Bachmann, G.E. Factores que Afectan el Éxito de Apareamiento de Machos de la Mosca Sudamericana de la Fruta, Anastrepha fraterculus (Diptera: Tephritidae). Ph.D. Thesis, Universidad de Buenos Aires, Buenos Aires, Argentina, 2016. [Google Scholar]
- Jin, S.; Zhou, X.; Gu, F.; Zhong, G.; Yi, X. Olfactory plasticity: Variation in the expression of chemosensory receptors in Bactrocera dorsalis in different physiological states. Front. Physiol. 2017, 8, 672. [Google Scholar] [CrossRef] [PubMed]
- Kuzmich, D.; Kawagoe, Z.A.; Walse, S.S. PVC Formulation of Anastrepha suspensa Pheromones Suitable for Field Studies. Engineering 2020, 7, 1646–1650. [Google Scholar] [CrossRef]
- Crawley, R.J. The R Book; Jhon Wiley & Sons: Chichester, UK, 2007. [Google Scholar]
- Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Casanoves, F.; Di Rienzo, J.A.; Robledo, C.W. Infostat User´s Guide; Editorial Brujas: Córdoba, Argentina, 2018. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat; versión 2020; Centro de Transferencia InfoStat, FCA; Universidad Nacional de Córdoba: Córdoba, Argentina, 2020; Available online: http://www.infostat.com.ar (accessed on 5 January 2023).
- Wu, A.; Li, X.; Yan, X.; Fan, W.; Hao, C. Electroantennogram responses of Plutella xylostella (L.), to sex pheromone components and host plant volatile semiochemicals. J. App. Entomol. 2020, 144, 396–406. [Google Scholar] [CrossRef]
- Guo, H.; Smith, D.P. Time-Dependent Odorant Sensitivity Modulation in Insects. Insects 2022, 13, 354. [Google Scholar] [CrossRef]
- Shen, H.H. How do mosquitoes smell us? The answers could help eradicate disease. Proc. Nat. Acad. Sci. USA 2017, 114, 2096–2098. [Google Scholar] [CrossRef]
- Su, C.Y.; Wang, J.W. Modulation of neural circuits: How stimulus context shapes innate behavior in Drosophila. Curr. Opin. Neurobiol. 2014, 29, 9–16. [Google Scholar] [CrossRef]
- Córdova-García, G.; Esquivel, C.J.; Pérez-Staples, D.; Ruiz-May, E.; Herrera-Cruz, M.; Reyes-Hernández, M.; Abraham, S.; Aluja, M.; Sirot, L. Characterization of reproductive proteins in the Mexican fruit fly points towards the evolution of novel functions. Proc. R. Soc. B 2022, 289, 20212806. [Google Scholar] [CrossRef] [PubMed]
- Jang, E.B.; McInnis, D.O.; Lance, D.; Carvalho, L. Mating-induced changes in olfactory-mediated behaviour of laboratory-reared normal, sterile and wild female Mediterranean fruit flies (Diptera: Tephritidae) mated to conespecific males. Ann. Entomol. Soc. Am. 1998, 91, 139–144. [Google Scholar] [CrossRef]
- Devescovi, F.; Hurtado, J.; Taylor, P.W. Mating-induced changes in responses of female Queensland fruit fly to male pheromones and fruit: A mechanism for mating-induced sexual inhibition. J. Insect Physiol. 2021, 129, 104195. [Google Scholar] [CrossRef]
- Córdova-García, G.; Sirot, L.; Abraham, S.; Díaz-Fleischer, F.; Flores-Estevez, N.; López-Ortega, M.; Pérez-Staples, D. Mating, but not male accessory gland products, changes female response to olfactory cues in Anastrepha fruit flies. Front. Physiol. 2021, 12, 714247. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.V. Interacciones Tritróficas Entre Moscas de la Fruta (Diptera, Tephritidae), sus Hospedantes y Parasitoides en las Principales Regiones de Producción Frutícola del Uruguay. Ph.D. Thesis, Universidad de la República, Montevideo, Uruguay, 2020. [Google Scholar]
- Campanini, E.B.; Congrains, C.; Torres, F.R.; de Brito, R.A. Odorant-binding proteins expression patterns in recently diverged species of Anastrepha fruit flies. Sci. Rep. 2017, 7, 2194. [Google Scholar] [CrossRef]
- Sims, C.; Birkett, M.A.; Withall, D.M. Enantiomeric Discrimination in Insects: The Role of OBPs and ORs. Insects 2022, 13, 368. [Google Scholar] [CrossRef]
- Juárez, M.L.; Pimper, L.E.; Bachmann, G.E.; Conte, C.A.; Ruiz, M.J.; Goane, L.; Medina Pereyra, P.; Castro, F.; Cladera, J.L.; Fernández, P.C.; et al. Gut bacterial diversity and physiological traits of Anastrepha fraterculus Brazilian-1 morphotype males are affected by antibiotic treatment. BMC Microbiol. 2019, 19, 283. [Google Scholar] [CrossRef]
- Belliard, S.A.; Bachmann, G.E.; Fernández, P.C.; Hurtado, J.; Vera, M.T.; Segura, D.F. Identification of host plant volatile stimulants of Anastrepha fraterculus male courtship behavior. Front. Ecol. Evol. 2022, 10, 908. [Google Scholar] [CrossRef]
- Robacker, D.C.; Hart, W.G. (Z)-3-nonenol, (Z,Z)-3,6-nonadienol and (S,S)-(–)- epianastrephin: Male produced pheromones of the Mexican fruit fly. Entomol. Exp. Appl. 1985, 39, 103–108. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | df | F-Value | p-Value |
|---|---|---|---|
| (Intercept) | 1 | 459.12 | <0.0001 |
| Sex | 1 | 3.44 | 0.0659 |
| Physiological condition | 2 | 6.97 | 0.0013 |
| Semiochemical | 1 | 11.34 | 0.0010 |
| Sex x Physiological condition | 2 | 3.66 | 0.0285 |
| Physiological condition x semiochemical | 2 | 0.10 | 0.9017 |
| Sex x Semiochemical | 1 | 0.08 | 0.7737 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goane, L.; Carrizo, B.N.; Ruiz, M.J.; Bachmann, G.E.; Milla, F.H.; Segura, D.F.; Kuzmich, D.; Walse, S.; Vera, M.T. Behavioural and Electrophysiological Response of Anastrepha fraterculus (Diptera: Tephritidae) to a γ-Lactone Synthetic Semiochemical. Insects 2023, 14, 206. https://doi.org/10.3390/insects14020206
Goane L, Carrizo BN, Ruiz MJ, Bachmann GE, Milla FH, Segura DF, Kuzmich D, Walse S, Vera MT. Behavioural and Electrophysiological Response of Anastrepha fraterculus (Diptera: Tephritidae) to a γ-Lactone Synthetic Semiochemical. Insects. 2023; 14(2):206. https://doi.org/10.3390/insects14020206
Chicago/Turabian StyleGoane, Lucía, Beatriz N. Carrizo, María Josefina Ruiz, Guillermo E. Bachmann, Fabian H. Milla, Diego F. Segura, Dan Kuzmich, Spencer Walse, and María Teresa Vera. 2023. "Behavioural and Electrophysiological Response of Anastrepha fraterculus (Diptera: Tephritidae) to a γ-Lactone Synthetic Semiochemical" Insects 14, no. 2: 206. https://doi.org/10.3390/insects14020206
APA StyleGoane, L., Carrizo, B. N., Ruiz, M. J., Bachmann, G. E., Milla, F. H., Segura, D. F., Kuzmich, D., Walse, S., & Vera, M. T. (2023). Behavioural and Electrophysiological Response of Anastrepha fraterculus (Diptera: Tephritidae) to a γ-Lactone Synthetic Semiochemical. Insects, 14(2), 206. https://doi.org/10.3390/insects14020206