The Impact of a Cypovirus on Parental and Filial Generations of Lymantria dispar L.

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and DsCPV-1
2.2. Infection of a Parental L. dispar Generation with DsCPV-1
2.3. The Impact of DsCPV-1 Infection on a Filial L. dispar Generation
2.4. qPCR for Assessing the Possible Vertical Transmission of DsCPV-1
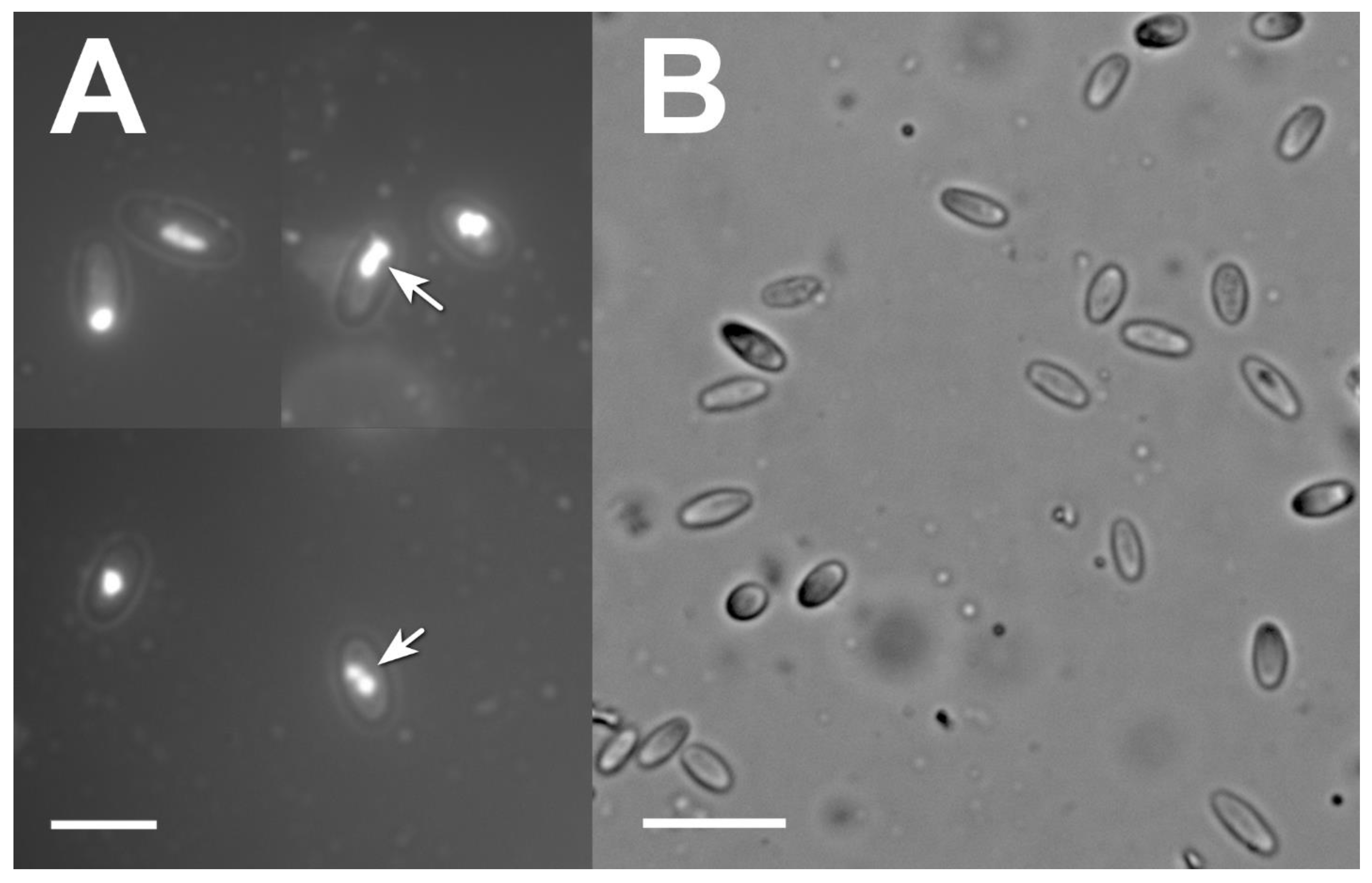
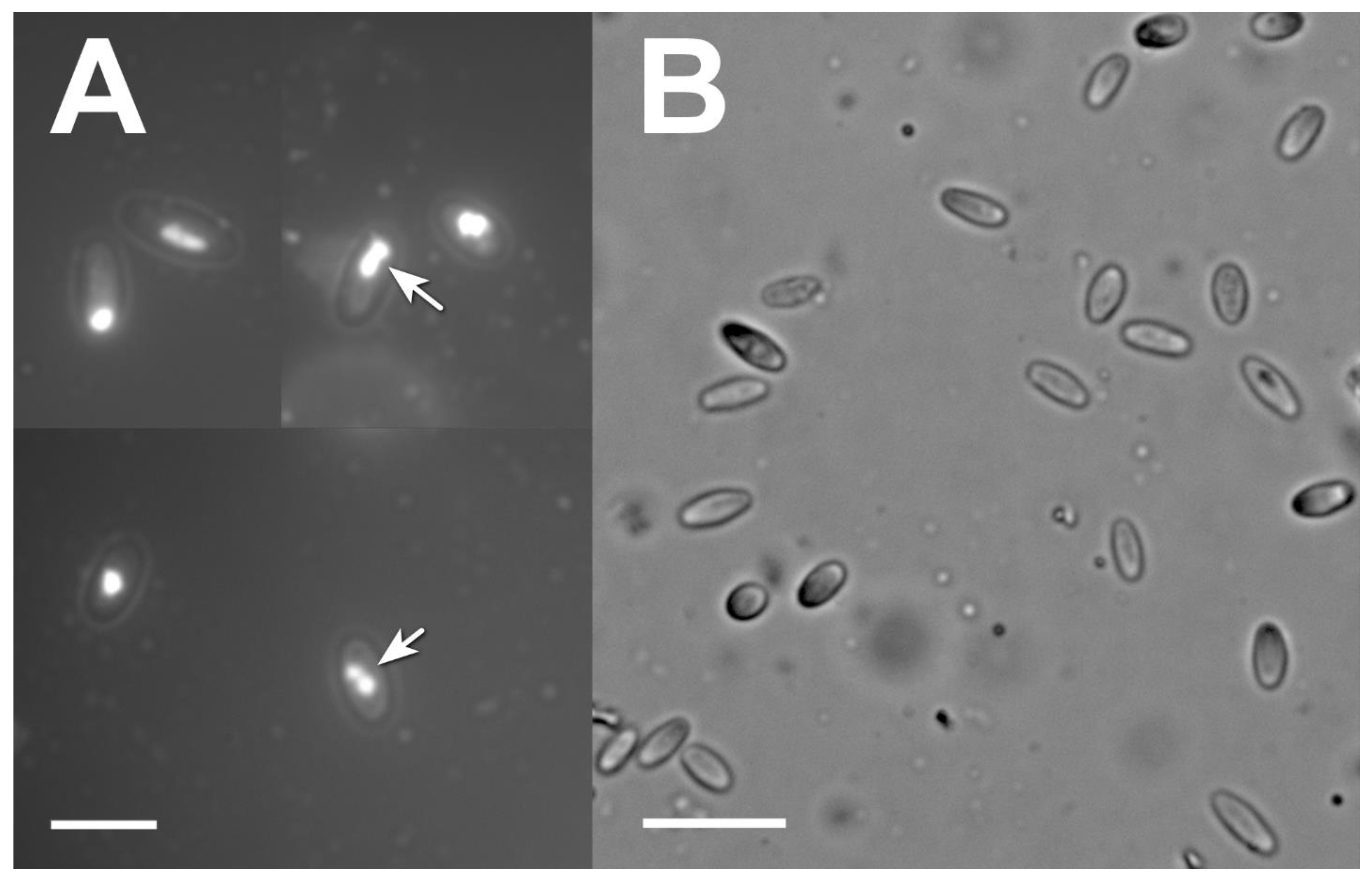
2.5. Microsporidia Detection and Identification
2.6. Statistical Analysis
3. Results
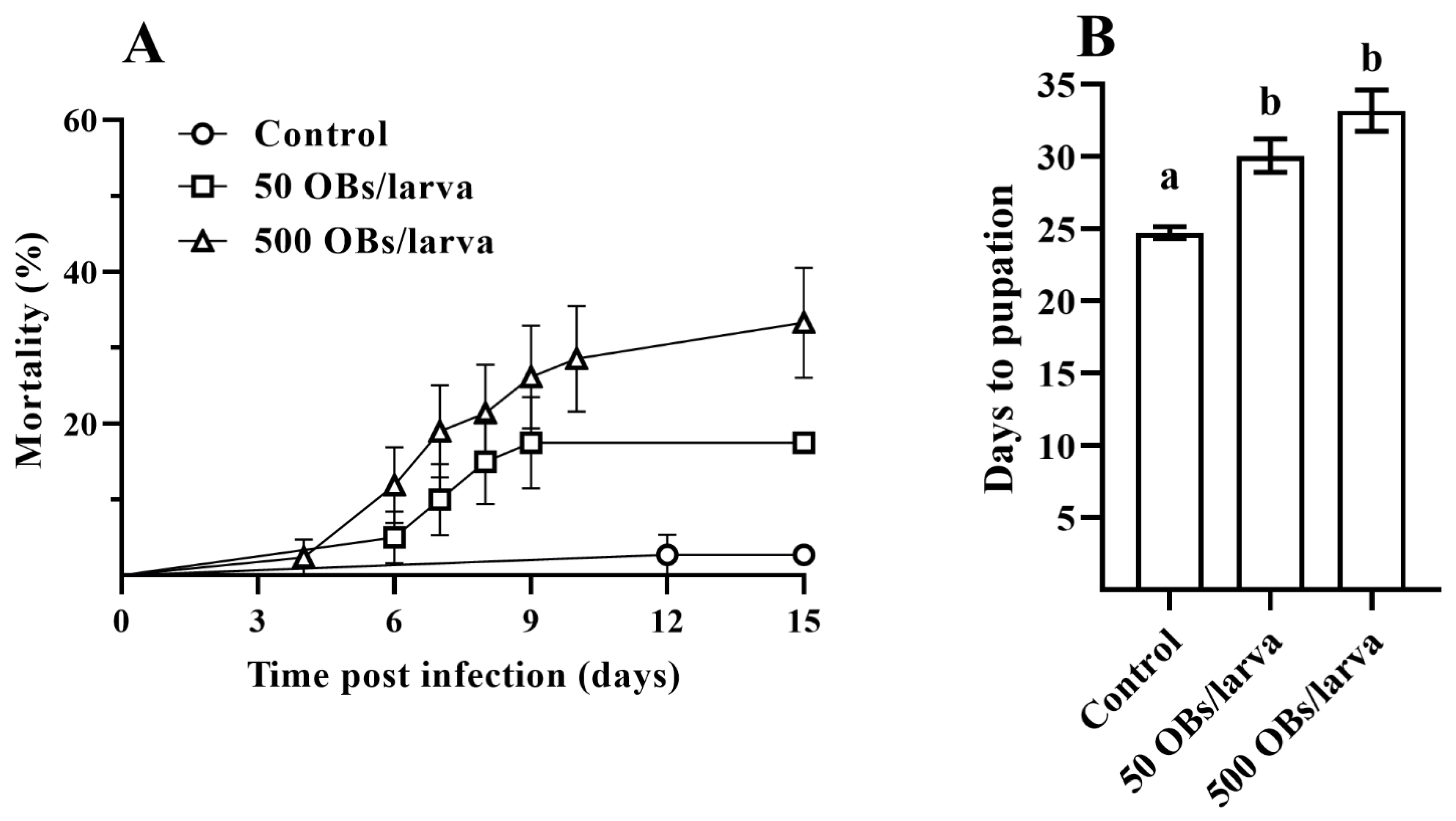
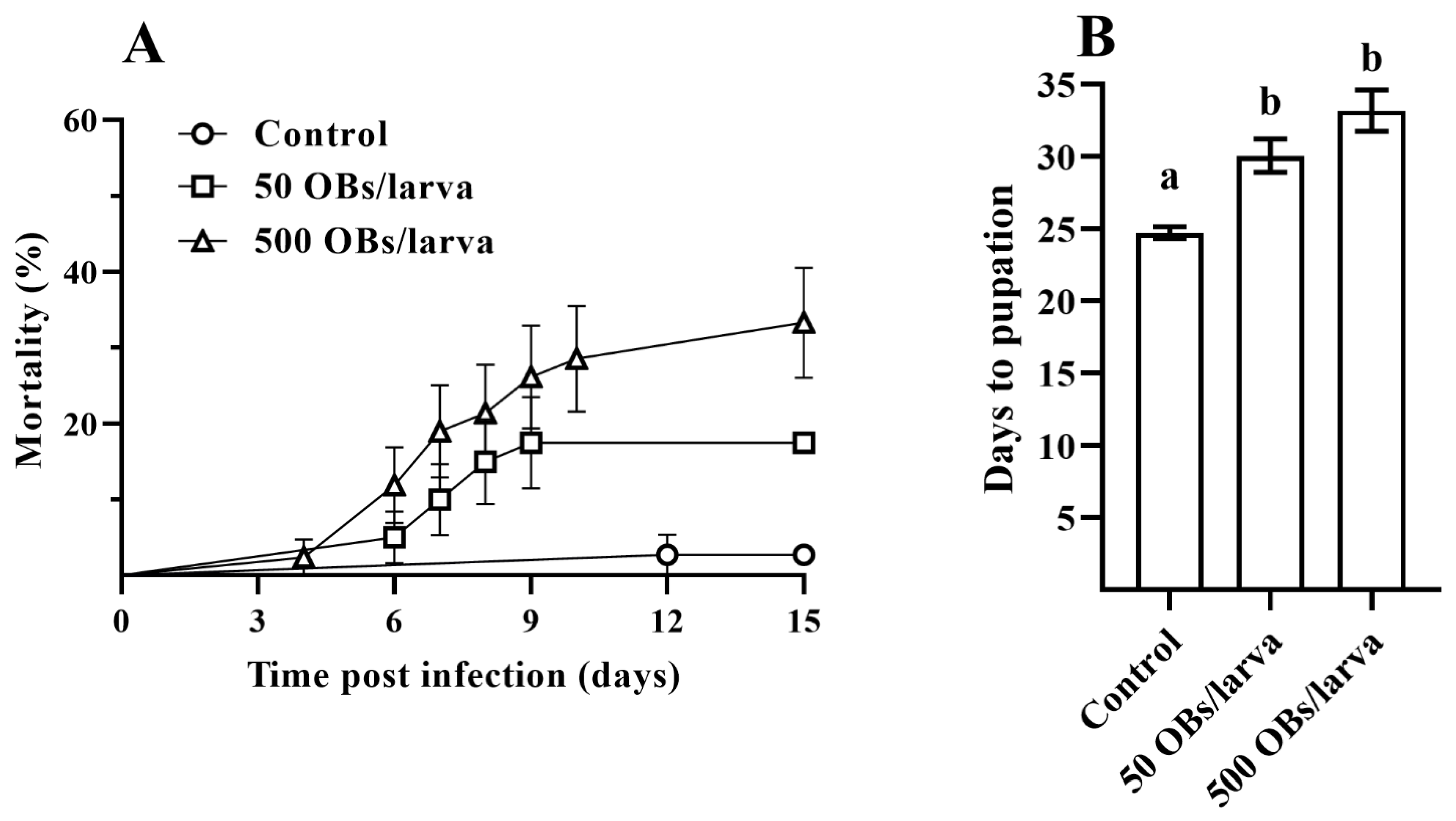
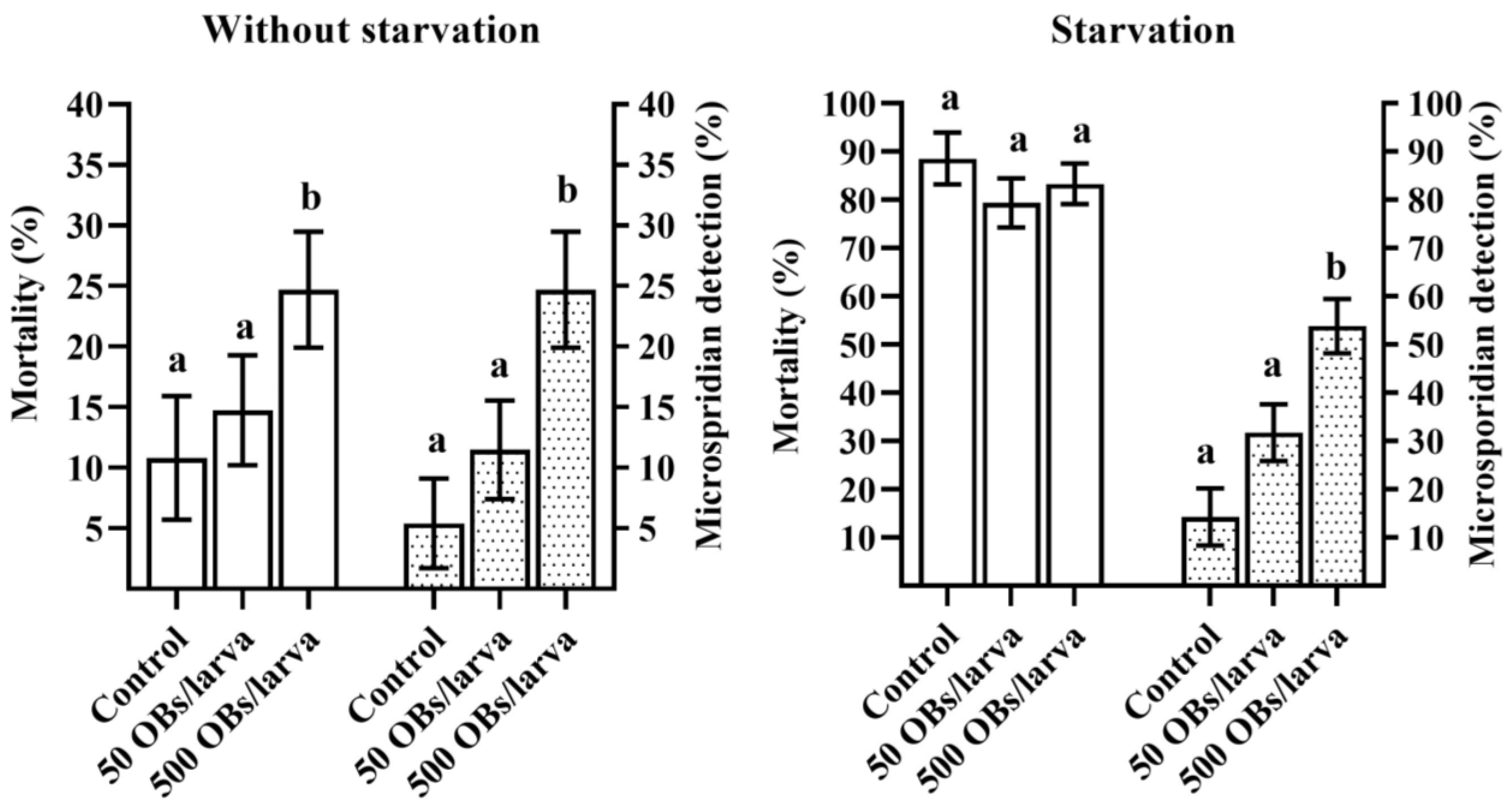
3.1. The Pathogenicity of DsCPV-1 and Life History-Traits of the Parental L. dispar Generation
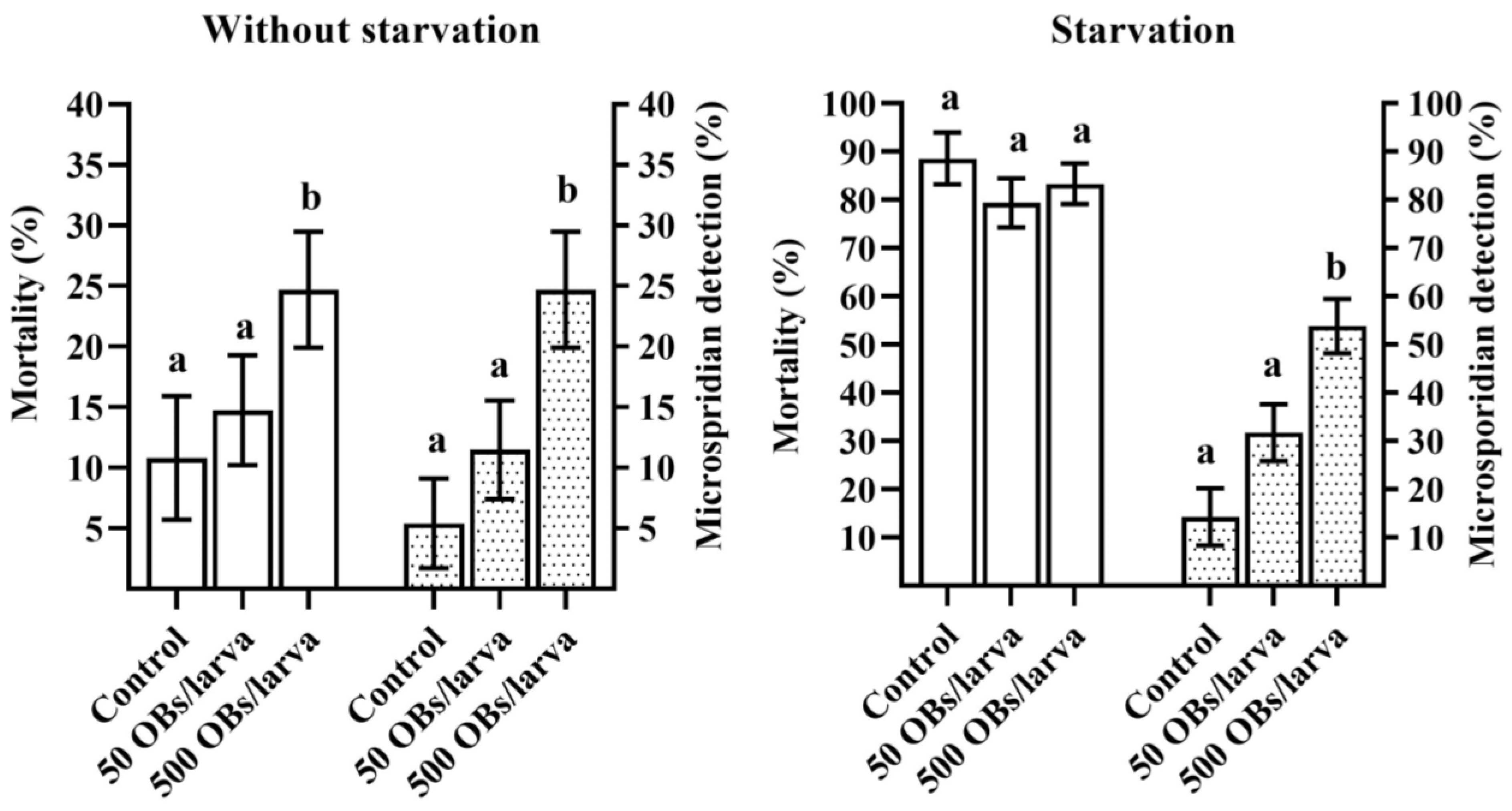
3.2. The Impact of DsCPV-1 Infection on the L. dispar Filial Generation
3.3. Microsporidia Detection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lacey, L.A. Chapter 1—Entomopathogens used as microbial control agents. In Microbial Control of Insect and Mite Pests; Academic Press: Cambridge, MA, USA, 2017; pp. 3–12. [Google Scholar]
- Hajek, A.E.; Gardescu, S.; Delalibera, I. Summary of classical biological control introductions of entomopathogens and nematodes for insect control. BioControl 2021, 66, 167–180. [Google Scholar] [CrossRef]
- Solter, L.F.; Hajek, A.E.; Lacey, L.A. Chapter 2—Exploration for entomopathogens. In Microbial Control of Insect and Mite Pests; Academic Press: Cambridge, MA, USA, 2017; pp. 13–23. [Google Scholar]
- Slack, J.; Arif, B.M. The Baculoviruses occlusion-derived virus: Virion structure and function. Adv. Virus Res. 2006, 69, 99–165. [Google Scholar]
- Lefkowitz, E.J.; Dempsey, D.M.; Hendrickson, R.C.; Orton, R.J.; Siddell, S.G.; Smith, D.B. Virus Taxonomy: The Database of the International Committee on Taxonomy of Viruses (ICTV). Nucleic Acids Res. 2018, 46, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Belloncik, S. Cytoplasmic polyhedrosis viruses—Reoviridae. Adv. Virus Res. 1989, 37, 173–209. [Google Scholar] [PubMed]
- Zhan, Z.; Guan, L.; Wang, J.; Liu, Z.; Guo, Y.; Xiao, Y.; Wang, H.; Jin, L. Isolation and genomic characterization of a cypovirus from the Oleander hawk moth, Daphnis nerii. J. Invertebr. Pathol. 2019, 163, 43–47. [Google Scholar] [CrossRef]
- Pavlushin, S.V.; Ilinsky, Y.Y.; Belousova, I.A.; Bayborodin, S.I.; Lunev, E.A.; Kechin, A.A.; Khrapov, E.A.; Filipenko, M.L.; Toshchakov, S.V.; Martemyanov, V.V. Appearances are deceptive: Three RNA viruses co-infected with the nucleopolyhedrovirus in host Lymantria dispar. Virus Res. 2021, 297, 198371. [Google Scholar] [CrossRef]
- Belloncik, S.; Mori, H. Cypoviruses. In The Insect Viruses; Miller, L.K., Ball, L.A., Eds.; Springer: Boston, MA, USA, 1998; pp. 337–369. [Google Scholar]
- Green, T.B.; White, S.; Rao, S.; Mertens, P.P.C.; Adler, P.H.; Becnel, J.J. Biological and molecular studies of a cypovirus from the black fly Simulium ubiquitum (Diptera: Simuliidae). J. Invertebr. Pathol. 2007, 95, 26–32. [Google Scholar] [CrossRef]
- Williams, T.; Virto, C.; Murillo, R.; Caballero, P. Covert Infection of Insects by Baculoviruses. Front. Microbiol. 2017, 8, 1337. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Sun, J.Z.; Weidner, E.H.; Lamotte, L.R. Stressors and rearing diseases of Trichoplusia ni: Evidence of vertical transmission of NPV and CPV. J. Invertebr. Pathol. 1999, 74, 149–155. [Google Scholar] [CrossRef]
- Virto, C.; Navarro, D.; Tellez, M.M.; Murillo, R.; Williams, T.; Caballero, P. Chemical and biological stress factors on the activation of nucleopolyhedrovirus infections in covertly infected Spodoptera exigua. J. Appl. Entomol. 2017, 141, 384–392. [Google Scholar] [CrossRef]
- Boukouvala, M.C.; Kavallieratos, N.G.; Skourti, A.; Pons, X.; Alonso, C.L.; Eizaguirre, M.; Fernandez, E.B.; Solera, E.D.; Fita, S.; Bohinc, T.; et al. Lymantria dispar (L.) (Lepidoptera: Erebidae): Current status of biology, ecology, and management in Europe with notes from North America. Insects 2022, 13, 854. [Google Scholar] [CrossRef]
- Alalouni, U.; Schädler, M.; Brandl, R. Natural enemies and environmental factors affecting the population dynamics of the gypsy moth. J. Appl. Entomol. 2013, 10, 721–738. [Google Scholar] [CrossRef]
- Hajek, A.E.; van Frankenhuyzen, K. Use of entomopathogens against forest pests. In Microbial Control of Insect and Mite Pests; Academic Press: Cambridge, MA, USA, 2017; pp. 313–330. [Google Scholar]
- Elkinton, J.S. Gypsy moth. In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 435–439. [Google Scholar]
- Martemyanov, V.V.; Akhanaev, Y.B.; Belousova, I.A.; Pavlusin, S.V.; Yakimova, M.E.; Kharlamova, D.D.; Ageev, A.A.; Golovina, A.N.; Astapenko, S.A.; Kolosov, A.V.; et al. A new cypovirus-1 strain as a promising agent for Lepidopteran pest control. Microbiol. Spectr. 2023, 11, e0385522. [Google Scholar] [CrossRef]
- Hughes, P.R.; van Beek, N.A.M.; Wood, H.A. A modified droplet feeding method for rapid assay of Bacillus thuringiensis and baculoviruses in Noctuid larvae. J. Invertebr. Pathol. 1986, 48, 187–192. [Google Scholar] [CrossRef]
- Malysh, J.M.; Kononchuk, A.G.; Frolov, A.N. Detection of microsporidia infecting beet webworm Loxostege sticticalis (Pyraloidea: Crambidae) in European part of Russia in 2006–2008. Plant Prot. News 2019, 2, 45–51. [Google Scholar] [CrossRef]
- Weiss, L.M.; Vossbrinck, C.R. Molecular biology, molecular phylogeny, and molecular diagnostic approaches to the microsporidia. In The Microsporidia and Microsporidiosis; Wittner, M., Weiss, L.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 1999; pp. 129–171. [Google Scholar]
- Vogelstein, B.; Gillespie, D. Preparative and analytical purification of DNA from Agarose. Proc. Natl. Acad. Sci. USA 1979, 76, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R Foundation for Statistical Computing: A Language and Environment for Statistical Computing; The R Project for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.r-project.org/ (accessed on 14 May 2023).
- Hlina, B.L.; Birceanu, O.; Robinson, C.S.; Dhiyebi, H.; Wilkie, M.P. The relationship between thermal physiology and lampricide sensitivity in larval sea lamprey (Petromyzon Marinus). J. Great Lakes Res. 2021, 47, 272–284. [Google Scholar] [CrossRef]
- Rothman, L.D.; Myers, J.H. Debilitating effects of viral diseases on host Lepidoptera. J. Invertebr. Pathol. 1996, 67, 1–10. [Google Scholar] [CrossRef]
- Echeverry, F.; Bergeron, J.; Kaupp, W.; Guertin, C.; Arella, M. Sequence analysis and expression of the polyhedrin gene of choristoneura fumiferana cytoplasmic polyhedrosis virus (CdCPV). Gene 1997, 198, 399–406. [Google Scholar] [CrossRef]
- Fossiez, F.; Belloncik, S.; Arella, M. Nucleotide sequence of the polyhedrin gene of Euxoa scandens cytoplasmic polyhedrosis virus (EsCPV). Virology 1989, 169, 462–465. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.S.; Possee, R.D.; King, L.A. Activation and detection of a latent baculovirus resembling Mamestra brassicae nuclear polyhedrosis virus in M. brassicae insects. Virology 1993, 194, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Murillo, R.; Hussey, M.S.; Possee, R.D. Evidence for covert baculovirus infections in a Spodoptera exigua laboratory culture. J. Gen. Virol. 2011, 92, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Fuxa, J.R.; Richter, A.R.; Ameen, A.O.; Hammock, B.D. Vertical transmission of TnSNPV, TnCPV, AcMNPV, and possibly recombinant NPV in Trichoplusia ni. J. Invertebr. Pathol. 2002, 79, 44–50. [Google Scholar] [CrossRef]
- Yang, M.M.; Zhai, W.J.; Li, B.C.; Zhang, W.; Zhang, Z.Q.; Li, M.L.; Zhang, Y.A. Covert LdMNPV detected in Lymantria dispar larvae during a survey of potential hosts for DekiNPV production. Int. J. Pest Manag. 2015, 61, 26–29. [Google Scholar] [CrossRef]
- Maddox, J.V.; Baker, M.D.; Jeffords, M.R.; Kuras, M.; Linde, A.; Solter, L.F.; McManus, M.L.; Vávra, J.; Vossbrinck, C.R. Nosema portugal, N. sp., isolated from gypsy moths (Lymantria dispar L.) collected in Portugal. J. Invertebr. Pathol. 1999, 73, 1–14. [Google Scholar] [CrossRef]
- Pilarska, D.; Takov, D.; Hyliš, M.; Radek, R.; Fiala, I.; Solter, L.; Linde, A. Natural occurrence of microsporidia infecting Lepidoptera in Bulgaria. Acta Parasitol. 2017, 62, 858–869. [Google Scholar] [CrossRef]
- McManus, M.L.; Solter, L. Microsporidian pathogens in European gypsy moth populations. In Proc: Ecology, Survey, and Management of Forest Insects; USDA Forest Service: Washington, DC, USA, 2003; pp. 44–51. [Google Scholar]
- Bakhvalov, S.A.; Ilinykh, A.V.; Zhimerikhin, V.N.; Martemyanov, V.V. Dynamics of black arches Lymantria monacha L. and gypsy moth L. dispar L. (Lymantriidae, Lepidoptera): Role of food resources and virus infection. Evraz. Entomol. Zh. 2002, 1, 101–108. [Google Scholar]
- Kononchuk, A.G.; Martemyanov, V.V.; Ignatieva, A.N.; Belousova, I.A.; Inoue, M.N.; Tokarev, Y.S. Susceptibility of the gypsy moth Lymantria dispar (Lepidoptera: Erebidae) to Nosema pyrausta (Microsporidia: Nosematidae). Insects 2021, 12, 447. [Google Scholar] [CrossRef]
- Goertz, D.; Hoch, G. Vertical transmission and overwintering of microsporidia in the gypsy moth, Lymantria dispar. J. Invertebr. Pathol. 2008, 99, 43–48. [Google Scholar] [CrossRef]
- Bauer, L.S.; Miller, D.L.; Maddox, J.V.; McManus, M.L. Interactions between a Nosema sp. (Microspora: Nosematidae) and nuclear polyhedrosis virus infecting the gypsy moth, Lymantria dispar (Lepidoptera: Lymantriidae). J. Invertebr. Pathol. 1998, 72, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Pavlushin, S.V.; Belousova, I.A.; Chertkova, E.A.; Akhanaev, Y.B.; Martemyanov, V.V.; Glupov, V.V. Effect of starvation as a population stress-factor on the activation of covert baculovirus infection in the gypsy moth. Biol. Bull. Rev. 2021, 11, 86–91. [Google Scholar] [CrossRef]
- Akhanaev, Y.B.; Belousova, I.A.; Lebedeva, D.A.; Pavlushin, S.V.; Martemyanov, V.V. A comparison of the vertical transmission of high-and low-virulence nucleopolyhedrovirus strains in Lymantria dispar L. Insects 2020, 11, 455. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akhanaev, Y.B.; Pavlushin, S.V.; Kharlamova, D.D.; Odnoprienko, D.; Subbotina, A.O.; Belousova, I.A.; Ignatieva, A.N.; Kononchuk, A.G.; Tokarev, Y.S.; Martemyanov, V.V. The Impact of a Cypovirus on Parental and Filial Generations of Lymantria dispar L. Insects 2023, 14, 917. https://doi.org/10.3390/insects14120917
Akhanaev YB, Pavlushin SV, Kharlamova DD, Odnoprienko D, Subbotina AO, Belousova IA, Ignatieva AN, Kononchuk AG, Tokarev YS, Martemyanov VV. The Impact of a Cypovirus on Parental and Filial Generations of Lymantria dispar L. Insects. 2023; 14(12):917. https://doi.org/10.3390/insects14120917
Chicago/Turabian StyleAkhanaev, Yuriy B., Sergey V. Pavlushin, Daria D. Kharlamova, Daria Odnoprienko, Anna O. Subbotina, Irina A. Belousova, Anastasia N. Ignatieva, Anastasia G. Kononchuk, Yuri S. Tokarev, and Vyacheslav V. Martemyanov. 2023. "The Impact of a Cypovirus on Parental and Filial Generations of Lymantria dispar L." Insects 14, no. 12: 917. https://doi.org/10.3390/insects14120917
APA StyleAkhanaev, Y. B., Pavlushin, S. V., Kharlamova, D. D., Odnoprienko, D., Subbotina, A. O., Belousova, I. A., Ignatieva, A. N., Kononchuk, A. G., Tokarev, Y. S., & Martemyanov, V. V. (2023). The Impact of a Cypovirus on Parental and Filial Generations of Lymantria dispar L. Insects, 14(12), 917. https://doi.org/10.3390/insects14120917