
Insect Freeze-Tolerance Downunder: The Microbial Connection



Simple Summary
Abstract
1. Introduction

2. Ice-Nucleating Agents (Ice+)
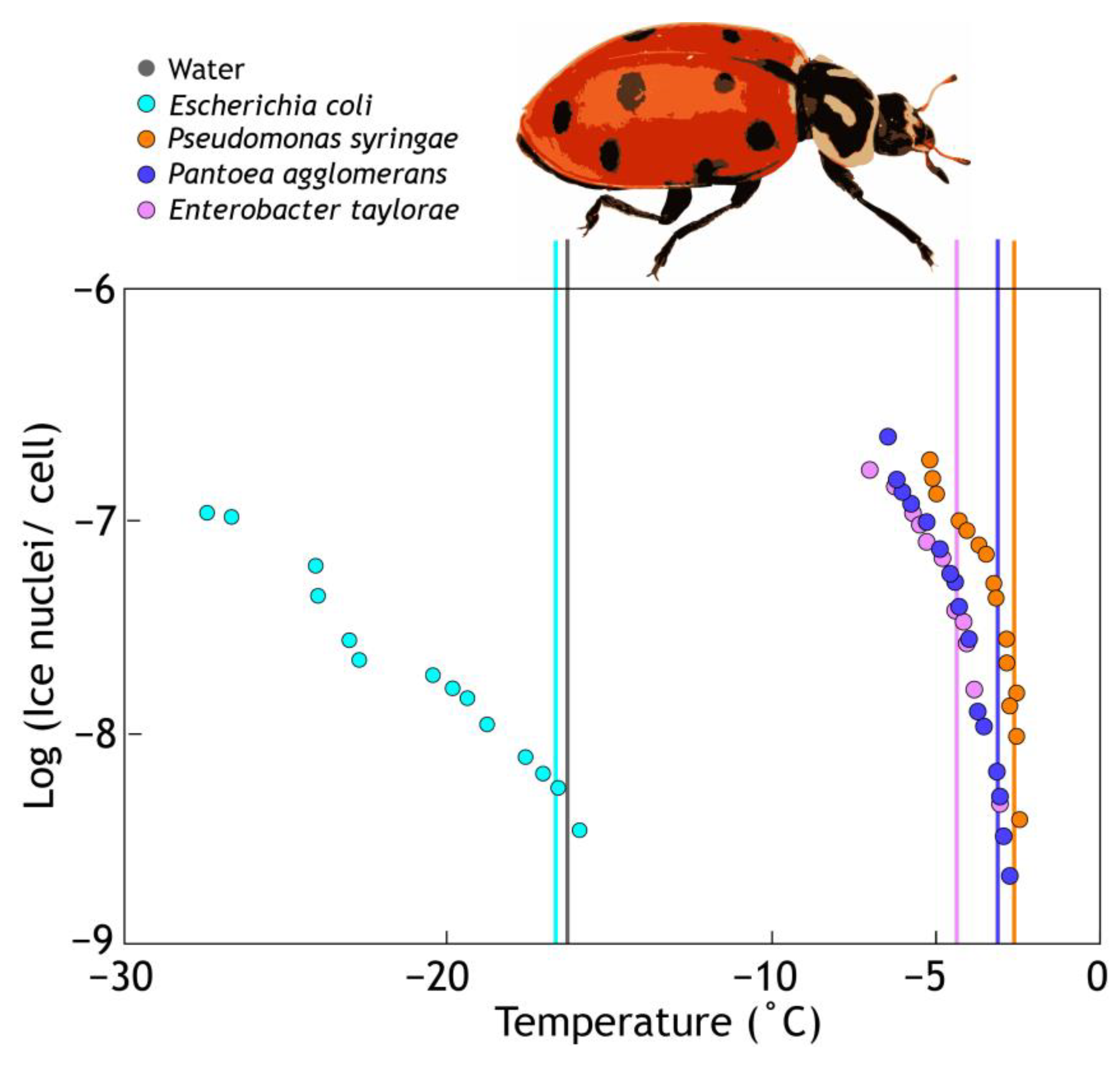
3. Sources of Ice+ Particles

4. Extended Genotype
5. Transmission of Insect Microbes
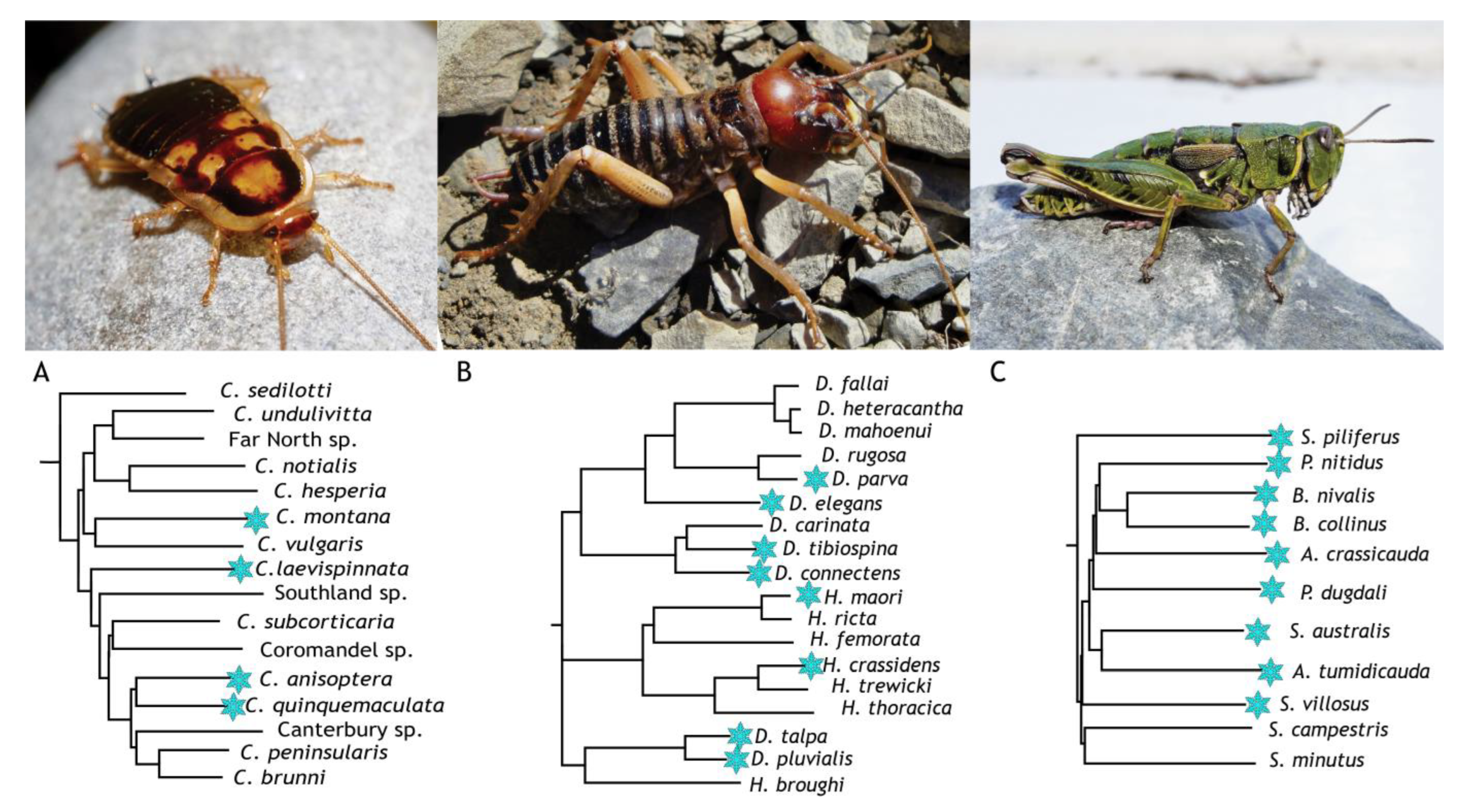
6. Freeze-Tolerant Insects in Aotearoa/New Zealand

6.1. Celatoblatta quinquemaculata—The Otago Alpine Cockroach
6.2. Hemideina maori—The Mountain Tree Wētā
6.3. Sigaus australis—The Southern Alpine Grasshopper
7. Exploring the Microbiome

Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lisovski, S.; Ramenofsky, M.; Wingfield, J.C. Defining the degree of seasonality and its significance for future research. Integ. Comp. Biol. 2017, 57, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef]
- Tougeron, K. Diapause research in insects: Historical review and recent work perspectives. Entomol. Exp. Appl. 2019, 167, 27–36. [Google Scholar] [CrossRef]
- Fielding, D.J. Diapause traits of Melanoplus sanguinipes and Melanoplus borealis (Orthoptera: Acrididae). Ann. Entomol. Soc. Am. 2008, 101, 439–448. [Google Scholar] [CrossRef]
- Sinclair, B.J. Insect cold tolerance: How many kinds of frozen? Eur. J. Entomol. 1999, 96, 157–164. [Google Scholar]
- Graether, S.; Kuiper, M.; Gagné, S.M.; Walker, V.K.; Jia, Z.; Sykes, B.D.; Davies, P.L. β-Helix structure and ice-binding properties of a hyperactive antifreeze protein from an insect. Nature 2000, 406, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Toxopeus, J.; Sinclair, B.J. Mechanisms underlying insect freeze tolerance. Biol. Rev. 2018, 93, 1891–1914. [Google Scholar] [CrossRef] [PubMed]
- Poikela, N.; Tyukmaeva, V.; Hoikkala, A.; Kankare, M. Multiple paths to cold tolerance: The role of environmental cues, morphological traits and the circadian clock gene vrille. BMC Ecol. Evol. 2021, 21, 117. [Google Scholar] [CrossRef]
- Roberts, R.M. Seasonal strategies in insects. N. Z. Entomol. 1978, 6, 350–356. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Addo-Bediako, A.; Chown, S.L. Climatic variability and the evolution of insect freeze tolerance. Biol. Rev. 2003, 78, 181–195. [Google Scholar] [CrossRef]
- Franks, F. Water: A Matrix of Life, 2nd ed.; Royal Society of Chemistry: London, UK, 2000. [Google Scholar]
- Lock, G.S.H. The Growth and Decay of Ice; Cambridge University Press Inc.: Cambridge, UK, 1990. [Google Scholar]
- Lide, D.R. CRC Handbook of Chemistry and Physics, 84th ed.; CRC Press: Boca Raton, FL, USA, 2003; ISBN 9780849304842. [Google Scholar]
- Heverly, J.R. Supercooling and crystallization. Trans. Am. Geophys. Union 1949, 30, 205–210. [Google Scholar] [CrossRef]
- Lundheim, R. Physiological and ecological significance of biological ice nucleators. Philos. Trans. R. Soc. Lond. B 2002, 357, 937–943. [Google Scholar] [CrossRef] [PubMed]
- Mazur, P. A biologist’s view of the relevance of thermodynamics and physical chemistry to cryobiology. Cryobiology 2010, 60, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Salt, R.W. A comparison of injury and survival of larvae of Cephus cinctus Nort. after intracellular and extracellular freezing. Can. J. Zool. 1961, 39, 349–357. [Google Scholar] [CrossRef]
- Worland, M.R.; Wharton, D.A.; Byars, S.G. Intracellular freezing and survival in the freeze tolerant alpine cockroach C. quinquemaculata. J. Insect Physiol. 2004, 50, 225–232. [Google Scholar] [CrossRef]
- Duman, J.G.; Walters, K.R.; Sformo, T.; Carrasco, M.A.; Nickell, P.K.; Lin, X.; Barns, B.M. Antifreeze and ice-nucleator proteins. In Low Temperature Biology of Insects; Denlinger, D.L., Lee, R.E., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 59–90. [Google Scholar]
- Sformo, T.; Walters, K.; Jeannet, K.; Wowk, B.; Fahy, G.M.; Barnes, B.M.; Duman, J.G. Deep supercooling, vitrification and limited survival to −100 °C in the Alaskan beetle Cucujus clavipes puniceus (Coleoptera: Cucujidae) larvae. J. Exp. Biol. 2010, 213, 502–509. [Google Scholar] [CrossRef]
- Zachariassen, K.; Hammel, H. Nucleating agents in the haemolymph of insects tolerant to freezing. Nature 1976, 262, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E.; Zachariassen, K.E.; Baust, J.G. Effect of cryoprotectants on the activity of hemolymph nucleating agents in physical solutions. Cryobiology 1981, 18, 511–514. [Google Scholar] [CrossRef]
- Hawes, T.C.; Wharton, D.A. Freeze fitness in alpine Tiger moth caterpillars and their parasitoids. Oecologia 2011, 167, 39–48. [Google Scholar] [CrossRef]
- Paschke, D. Cold Tolerance Mechanisms of the New Zealand Alpine Wētā, Hemideina maori. Ph.D. Thesis, University of Otago, Dunedin, New Zealand, 2017. [Google Scholar]
- Lee, R.E. A primer on insect cold-tolerance. In Low Temperature Biology of Insects; Denlinger, D.L., Lee, R.E., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 3–34. [Google Scholar]
- Chown, S.L.; Nicolson, S.W. Insect Physiological Ecology: Mechanisms and Patterns; Oxford University Press Inc.: New York, NY, USA, 2004. [Google Scholar]
- Layne, J.R.; Edgar, C.L.; Mewith, R.E. Cold hardiness of the woolly bear caterpillar (Pyrrharctia isabella Lepidoptera: Arctiidae). Am. Midl. Nat. 1999, 141, 293–304. [Google Scholar] [CrossRef]
- Rozsypal, J.; Košťál, V. Supercooling and freezing as eco-physiological alternatives rather than mutually exclusive strategies: A case study in Pyrrhocoris apterus. J. Insect Physiol. 2018, 111, 53–62. [Google Scholar] [CrossRef] [PubMed]
- de Araujo, G.G.; Rodrigues, F.; Gonçalves, F.L.T.; Galante, D. Survival and ice nucleation activity of Pseudomonas syringae strains exposed to simulated high-altitude atmospheric conditions. Sci. Rep. 2019, 9, 7768. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E.; Lee, M.R.; Strong-Gunderson, J.M. Insect cold-hardiness and ice nucleating active microorganisms including their potential use for biological control. J. Insect Physiol. 1993, 39, 1–12. [Google Scholar] [CrossRef]
- Wharton, D.A. Supercooling and freezing tolerant animals. In Supercooling; Wilson, P., Ed.; IntechOpen: London, UK, 2012. [Google Scholar]
- Ali, F.; Wharton, D.A. Ice-active substances from the infective juveniles of the freeze tolerant entomopathogenic nematode, Steinernema feltiae. PLoS ONE 2016, 11, e0156502. [Google Scholar] [CrossRef]
- Denton, M.; Marshall, C. Laws of form revisited. Nature 2001, 410, 417. [Google Scholar] [CrossRef] [PubMed]
- Storey, K.B.; Storey, J.M. Molecular biology of freezing tolerance. Compr. Physiol. 2013, 3, 1283–1308. [Google Scholar]
- Lee, R.E.; Strong-Gunderson, J.M.; Lee, M.R.; Grove, K.S.; Riga, T.J. Isolation of ice nucleating active bacteria from insects. J. Exp. Zool. 1991, 257, 124–127. [Google Scholar] [CrossRef]
- Wharton, D.A.; Selvanesan, L.; Marshall, C.J. Ice-active proteins from New Zealand snow tussocks, Chionochloa macra and C. rigida. Cryo-Letters 2010, 31, 239–248. [Google Scholar]
- Lee, R.E.; Costanso, J.P.; Mugnano, J.A. Regulation of supercooling and ice nucleation in insects. Eur. J. Entomol. 1996, 93, 405–418. [Google Scholar]
- Wilson, P.W.; Heneghan, A.F.; Haymet, A.D.J. Ice nucleation in nature: Supercooling point (SCP) measurements and the role of heterogeneous nucleation. Cryobiology 2003, 46, 88–98. [Google Scholar] [CrossRef]
- Hill, T.C.J.; Moffett, B.F.; Demott, P.J.; Georgakopoulos, D.G.; Stump, W.L.; Franc, D. Measurement of ice nucleation-active bacteria on plants and in precipitation by quantitative PCR. Appl. Environ. Microbiol. 2014, 80, 1256–1267. [Google Scholar] [CrossRef] [PubMed]
- Failor, K.C.; Schmale, D.G.; Vinatzer, B.A.; Monteil, C.L. Ice nucleation active bacteria in precipitation are genetically diverse and nucleate ice by employing different mechanisms. ISME J. 2017, 11, 2740–2753. [Google Scholar] [CrossRef]
- Qui, Y.; Hudait, A.; Molinero, V. How size and aggregation of ice-binding proteins control their ice nucleation efficiency. J. Am. Chem. Soc. 2019, 141, 7439–7452. [Google Scholar]
- Vasebi, Y.; Llontop, M.E.M.; Hanlon, R.; Schmale, D.G.; Schnell, R.; Vinatzer, B.A. Comprehensive characterization of an aspen (Populus tremuloides) leaf litter sample that maintained ice nucleation activity for 48 years. Biogeosciences 2019, 16, 1675–1683. [Google Scholar] [CrossRef]
- O’Sullivan, D.; Murray, B.J.; Ross, J.F.; Webb, M.E. The adsorption of fungal ice-nucleating proteins on mineral dusts: A terrestrial reservoir of atmospheric ice-nucleating particles. Atmos. Chem. Phys. 2016, 16, 7879–7887. [Google Scholar] [CrossRef]
- Kunert, A.T.; Pöhlker, M.L.; Tang, K.; Krevert, C.S.; Wieder, C.; Speth, K.R.; Hanson, L.E.; Morris, C.E.; Schmale, D.G., III; Pöschl, U.; et al. Macromolecular fungal ice nuclei in Fusarium: Effects of physical and chemical processing. Biogeosciences 2019, 16, 4647–4659. [Google Scholar] [CrossRef]
- Wilson, M.; Lindow, S.E. Ecological similarity and coexistence of epiphytic ice-nucleating (Ice+) Pseudomonas syringae strains and a non-ice-nucleating (ice−) biological control agent. Appl. Environ. Microbiol. 1994, 60, 3128–3137. [Google Scholar] [CrossRef]
- Duman, J.G.; Neven, L.G.; Beals, J.M.; Olson, K.R.; Castellino, F.J. Freeze-tolerance adaptations, including haemolymph protein and lipoprotein nucleators, in the larvae of the cranefly Tipula trivittata. J. Insect Physiol. 1985, 31, 1–8. [Google Scholar] [CrossRef]
- Forbes, J.; Bissoyi, A.; Eickhoff, L.; Reicher, N.; Hansen, T.; Bon, C.G.; Walker, V.K.; Koop, T.; Rudich, Y.; Braslavsky, I.; et al. Water-organizing motif continuity is critical for potent ice nucleation protein activity. Nat. Commun. 2022, 13, 5019. [Google Scholar] [CrossRef]
- Gupta, R.; Deswal, R. Antifreeze proteins enable plants to survive in freezing conditions. J. Biosci. 2014, 39, 1–14. [Google Scholar] [CrossRef]
- Ganguly, M.; Dib, S.; Ariya, P. A Purely inorganic highly efficient ice nucleating particle. ACS Omega 2018, 3, 3384–3395. [Google Scholar] [CrossRef] [PubMed]
- Cascajo-Castresana, M.; David, R.O.; Iriarte-Alonso, M.A.; Bittner, A.M.; Marcolli, C. Protein aggregates nucleate ice: The example of apoferritin. Atmos. Chem. Phys. 2020, 20, 3291–3315. [Google Scholar] [CrossRef]
- Dawkins, R. The Extended Phenotype; Oxford University Press: Oxford, UK, 1982; 307p. [Google Scholar]
- Hendry, L.P.; Bruijning, M.; Forsberg, S.K.G.; Ayroles, J.F. The microbiome extends host evolutionary potential. Nat. Commun. 2021, 12, 5141. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Prabhu, A.; Aroney, S.T.N.; Rinke, C. Insights into plastic biodegradation: Community composition and functional capabilities of the superworm (Zophobas morio) microbiome in styrofoam feeding trials. Microb. Genom. 2022, 8, 000916. [Google Scholar]
- Savio, C.; Mugo-Kamiri, L.; Upfold, J.K. Bugs in bugs: The role of probiotics and prebiotics in maintenance of health in mass-reared insects. Insects 2022, 13, 376. [Google Scholar] [CrossRef]
- Hammer, T.J.; Moran, N.A. Links between metamorphosis and symbiosis in holometabolous insects. Philos. Trans. R. Soc. B 2019, 374, 20190068. [Google Scholar] [CrossRef]
- Nakano, M.; Morgan-Richards, M.; Trewick, S.A.; Clavijo-McCormick, A. Chemical ecology and olfaction in short-horned grasshoppers (Orthoptera: Acrididae). J Chem. Ecol. 2022, 48, 121–140. [Google Scholar] [CrossRef]
- Ferguson, L.V.; Dhakal, P.; Lebenzon, J.E.; Heinrichs, D.E.; Bucking, C.; Sinclair, B.J. Seasonal shifts in the insect gut microbiome are concurrent with changes in cold tolerance and immunity. Funct. Ecol. 2018, 32, 2357–2368. [Google Scholar] [CrossRef]
- Sommer, F.; Ståhlman, M.; Ilkayeva, O.; Arnemo, J.; Kindberg, J.; Josefsson, J.; Newgard, C.; Fröbert, O.; Bäckhed, F. The gut microbiota modulates energy metabolism in the hibernating Brown Bear Ursus arctos. Cell Rep. 2016, 14, 1655–1661. [Google Scholar] [CrossRef]
- Wharton, D.A.; Pow, B.; Kristensen, M.; Ramløv, H.; Marshall, C.J. Ice-active proteins and cryoprotectants from the New Zealand alpine cockroach, Celatoblatta quinquemaculata. J. Insect Physiol. 2009, 55, 27–31. [Google Scholar] [CrossRef]
- Worland, M.R.; Sinclair, B.J.; Wharton, D.A. Ice nucleator activity in a New Zealand alpine cockroach C. quinquemaculata (Dictyoptera: Blattidae). Cryo-Letters 1997, 18, 327–334. [Google Scholar]
- Sinclair, B.J.; Worland, M.R.; Wharton, D.A. Ice nucleation and freezing tolerance in New Zealand alpine and lowland weta, Hemideina spp. (Orthoptera; Stenopelmatidae). Physiol. Entomol. 1999, 24, 56–63. [Google Scholar] [CrossRef]
- Broderick, N.; Lemaitre, B. Gut-associated microbes of Drosophila melanogaster. Gut Microbes 2021, 3, 307–321. [Google Scholar] [CrossRef] [PubMed]
- Worland, M.R.; Block, W. Ice-nucleating bacteria from the guts of two sub-Antarctic beetles Hydromedion sparsutum and Perimylops antarcticus (Perimylopidae). Cryobiology 1999, 38, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Tsumuki, H.; Konno, H.; Maeda, T.; Okamoto, Y. An ice-nucleating active fungus isolated from the gut of the rice stem borer, Chilo suppressalis Walker (Lepidoptera: Pyralidae). J. Insect Physiol. 1992, 38, 119–125. [Google Scholar] [CrossRef]
- Strong-Gunderson, J.M.; Lee, R.E.; Lee, M.R.; Riga, T.J. Ingestion of ice nucleating active bacteria increases the supercooling point of the lady beetle Hippodamia convergens. J. Insect Physiol. 1990, 36, 153–157. [Google Scholar] [CrossRef]
- Gupta, A.; Nair, S. Dynamics of insect-microbiome interaction influence host and microbial symbiont. Front. Microbiol. 2020, 11, 1357. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, T.; Kikuchi, Y.; Nikoh, N.; Shimada, M.; Fukatsu, T. Strict Host-Symbiont Cospeciation and reductive genome evolution in insect gut bacteria. PLoS Biol. 2006, 4, e337. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hosokawa, T.; Nikoh, N.; Meng, X.-Y.; Kamagata, Y.; Fukatsu, T. Host-symbiont co-speciation and reductive genome evolution in gut symbiotic bacteria of acanthosomatid stinkbugs. BMC Biol. 2009, 7, 2. [Google Scholar] [CrossRef]
- Franco, F.P.; Túler, A.C.; Gallan, D.Z.; Gonçalves, F.G.; Favaris, A.P.; Peñaflor, M.F.G.V.; Leal, W.S.; Moura, D.S.; Bento, J.M.S.; Silva-Filho, M.C. Fungal phytopathogen modulates plant and insect response to promote it dissemination. ISME J. 2021, 15, 3522–3533. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N. A The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef] [PubMed]
- Nikoh, N.; Hosokawa, T.; Oshima, K.; Hattori, M.; Fukatsu, T. Reductive evolution of bacterial genome in insect gut environment. Genome Biol. Evol. 2011, 3, 702–714. [Google Scholar] [CrossRef] [PubMed]
- Lavy, O.; Gophna, U.; Gefen, E.; Ayali, A. Locust bacterial symbionts: An update. Insects 2020, 11, 655. [Google Scholar] [CrossRef] [PubMed]
- Rosas, T.; García-Ferris, C.; Domínguez-Santos, R.; Llop, P.; Latorre, A.; Moya, A. Rifampicin treatment of Blattella germanica evidences a fecal transmission route of their gut microbiota. FEMS Microbiol. Ecol. 2018, 94, fiy002. [Google Scholar] [CrossRef] [PubMed]
- Trewick, S.A.; Wallis, G.P.; Morgan-Richards, M. The invertebrate life of New Zealand: A phylogeographic approach. Insects 2011, 2, 297–325. [Google Scholar] [CrossRef] [PubMed]
- Buckley, T.R.; Hoare, R.J.B.; Leschen, R.A.B. Key question on the evolution and biogeography of New Zealand alpine insects. J. R. Soc. N. Z. 2022. [Google Scholar] [CrossRef]
- Block, W.; Wharton, D.; Sinclair, B. Cold tolerance of a New Zealand alpine cockroach, Celatoblatta quinquemaculata (Dictyoptera, Blattidae). Physiol. Entomol. 1998, 23, 1–6. [Google Scholar] [CrossRef]
- Ramløv, H.; Bedford, J.; Leader, J.; Ramurv, H. Freezing tolerance of the New Zealand alpine weta, Hemideina maori Hutton. J. Therm. Biol. 1992, 17, 51–54. [Google Scholar] [CrossRef]
- Hawes, T.C. Cryobiology canalization of freeze tolerance in an alpine grasshopper. Cryobiology 2015, 71, 356–359. [Google Scholar] [CrossRef]
- Hawes, T.C.; Wharton, D.A. Tolerance of freezing in caterpillars of the New Zealand Magpie moth (Nyctemera annulate). Physiol. Entomol. 2011, 35, 296–300. [Google Scholar] [CrossRef]
- Dennis, A.B.; Dunning, L.T.; Sinclair, B.J.; Buckley, T.R. Parallel molecular routes to cold adaptation in eight genera of New Zealand stick insects. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Twort, V.G.; Newcomb, R.D.; Buckley, T.R. New Zealand tree and giant wētā (Orthoptera) transcriptomics reveal divergent selection patterns in metabolic loci. Genome Biol. Evol. 2019, 11, 1293–1306. [Google Scholar] [CrossRef] [PubMed]
- Koot, E.; Morgan-Richards, M.; Trewick, S.A. An alpine grasshopper radiation older than the hills, on the Southern Alps (Kā Tiritiri o te Moana) of New Zealand (Aotearoa). Mol. Phylogen. Evol. 2020, 147, 106783. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, B.J. Field ecology of freeze tolerance: Interannual variation in cooling rates, freeze-thaw and thermal stress in the microhabitat of the alpine cockroach Celatoblatta quinquemaculata. Oikos 2001, 93, 286–293. [Google Scholar] [CrossRef]
- Sinclair, B.J. Seasonal variation in freezing tolerance of the New Zealand alpine cockroach Celatoblatta quinquemaculata. Ecol. Entomol. 1997, 22, 462–467. [Google Scholar] [CrossRef]
- Wharton, D.A. Cold tolerance of New Zealand alpine insects. J. Insect Physiol. 2011, 57, 1090–1095. [Google Scholar] [CrossRef]
- Johns, P.M. The cockroaches of New Zealand. Rec. Canterb. Mus. 1966, 8, 93–136. [Google Scholar]
- Zervos, S. Protrellus dalei n. sp., Blatticola barryi n. sp., and Suifunema mackenziei n. sp., thelastomatid nematodes from New Zealand cockroaches. N. Z. J. Zool. 1987, 14, 239–250. [Google Scholar] [CrossRef]
- Schauer, C.; Thompson, C.L.; Brune, A. The bacterial community in the gut of the cockroach Shelfordella lateralis reflects the close evolutionary relatedness of cockroaches and termites. Appl. Environ. Microbiol. 2012, 78, 2758–2767. [Google Scholar] [CrossRef]
- Sabree, Z.L.; Kambhamptis, S.; Moran, N.A. Nitrogen recycling and nutritional provisioning by Blattabacterium, the cockroach endosymbiont. Proc. Natl. Acad. Sci. USA 2014, 106, 19521–19526. [Google Scholar] [CrossRef]
- Sinclair, B.J.; Renault, D. Intracellular ice formation in insects: Unresolved after 50 years? Comp. Biochem. Physiol. Mol. Integr. Physiol. 2010, 155, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Thorne, M.A.S.; Seybold, A.; Marshall, C.; Wharton, D. Molecular snapshot of an intracellular freezing event in an Antarctic nematode. Cryobiology 2017, 75, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Ramløv, H.; Westh, P. Ice formation in the freeze tolerant alpine weta Hemideina maori Hutton (Orthoptera, Stenopelmatidae). Cryo-Letters 1993, 14, 169–176. [Google Scholar]
- Trewick, S.A.; Morgan-Richards, M. After the deluge: Mitochondrial DNA indicates Miocene radiation and Pliocene adaptation of tree and giant weta (Orthoptera: Anostostomatidae). J. Biogeog. 2005, 32, 295–309. [Google Scholar] [CrossRef]
- Trewick, S.A.; Wallis, G.P.; Morgan-Richards, M. Phylogeographic pattern correlates with Pliocene mountain-building in the alpine scree weta (Orthoptera, Anostostomatidae). Mol. Ecol. 2000, 9, 657–666. [Google Scholar] [CrossRef] [PubMed]
- King, K.J.; Sinclair, B.J. Water loss in tree weta (Hemideina): Adaptation to the montane environment and a test of the melanisation-desiccation resistance hypothesis. J. Exp. Biol. 2015, 218, 1995–2004. [Google Scholar] [CrossRef]
- Hance, T.; Boivin, G. Effect of parasitism by Anaphes sp. (Hyme- noptera: Mymaridae) on the cold hardiness of Listronotus oregonensis (Coleoptera: Curculionidae) eggs. Can. J. Zool. 1993, 71, 759–764. [Google Scholar] [CrossRef]
- Nowierski, R.M.; Fitzgerald, B.C. Supercooling capacity of Eurasian and North American populations of parasitoids of the Russian wheat aphid, Diuraphis noxia. Biocontrol 2002, 47, 279–292. [Google Scholar] [CrossRef]
- Tyrell, C.; Wharton, D.A.; Ramløv, H.; Moller, H. Cold tolerance of an endoparasitic nematode within a freezing-tolerant orthopteran host. Parasitology 1994, 109, 367–372. [Google Scholar] [CrossRef]
- Ward, D.F.; Thomas, F.; Poulin, R. Fluctuating asymmetry and parasitism in six New Zealand insects. Acta Oecol. 1998, 19, 409–412. [Google Scholar] [CrossRef]
- Hawes, T.C. Integumental buffering in an alpine grasshopper. Physiol. Entomol. 2014, 39, 280–284. [Google Scholar] [CrossRef]
- Xiong, H. Ice Activities in New Zealand Chionochloa Species. Ph.D. Thesis, University of Otago, Dunedin, New Zealand, 2018. [Google Scholar]
- White, E.G.; Watson, R.N. A food consumption study of three New Zealand alpine grasshopper species. N. Z. Agric. Res. 1972, 15, 867–877. [Google Scholar] [CrossRef]
- Lukas, M.; Schwidetzky, R.; Eufemio, R.; Bonn, M.; Meister, K. Towards understanding bacterial ice nucleation. J. Physl. Chem. B 2022, 126, 1861–1867. [Google Scholar] [CrossRef] [PubMed]
- Waite, D.W.; Dsouza, M.; Biswas, K.; Ward, D.F.; Deines, P.; Taylor, M.W. Microbial community structure in the gut of the New Zealand insect Auckland tree weta (Hemideina thoracica). Arch. Microbiol. 2015, 197, 603–612. [Google Scholar] [CrossRef]
- Huson, D.H.; Auch, A.F.; Qi, J.; Schuster, S.C. MEGAN analysis of metagenomic data. Genome Res. 2007, 17, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Marshall, C.J.; Basu, K.; Davies, P.L. Ice-shell purification of ice-binding proteins. Cryobiology 2016, 72, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Marcelino, V.R.; Wille, M.; Hurt, A.C.; González-Acuña, D.; Klaassen, M.; Schlub, T.E.; Eden, J.S.; Shi, M.; Iredell, J.R.; Sorrell, T.C.; et al. Meta-transcriptomics reveals a diverse antibiotic resistance gene pool in avian microbiomes. BMC Biol. 2019, 17, 31. [Google Scholar] [CrossRef]
- Muñoz-Benavent, M.; Pérez-Cobas, A.E.; García-Ferris, C.; Moya, A.; Latorre, A. Insects’ potential: Understanding the functional role of their gut microbiome. J. Pharm. Biomed. Anal. 2021, 194, 113787. [Google Scholar] [CrossRef]
- Culviner, P.H.; Guegler, C.K.; Laub, M.T. A simple, cost-effective, and robust method for rRNA depletion in RNA-Sequencing studies. MBio 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Song, S.J.; Sanders, J.G.; Baldassarre, D.T.; Chaves, J.A.; Johnson, N.S.; Piaggio, A.J.; Stuckey, M.J.; Nováková, E.; Metcalf, J.L.; Chomel, B.B.; et al. Is there convergence of gut microbes in blood-feeding vertebrates? Phil. Trans. R. Soc. B Biol. Sci. 2019, 374, 1777. [Google Scholar] [CrossRef]
- Cookson, A.L.; Biggs, P.J.; Marshall, J.C.; Reynolds, A.; Collis, R.M.; French, N.P.; Brightwell, G. Culture independent analysis using gnd as a target gene to assess Escherichia coli diversity and community structure. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Purcell, R.V.; Visnovska, M.; Biggs, P.J.; Schmeier, S.; Frizelle, F.A. Distinct gut microbiome patterns associate with consensus molecular subtypes of colorectal cancer. Sci. Rep. 2017, 7, 11590. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Species | Temperature of Crystallization °C | Lower Lethal Temperature °C | % of Body Water Frozen | Initiation of Freezing |
|---|---|---|---|---|---|
| Cockroach | Celatoblatta quinquemaculata | −5.4 | −8.9 | 74 | gut |
| Wētā | Hemideina maori | −3.8 | −10 | 82 | hindgut |
| Grasshopper | Sigaus australis | −4 | −11 | Not known | Not known |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morgan-Richards, M.; Marshall, C.J.; Biggs, P.J.; Trewick, S.A. Insect Freeze-Tolerance Downunder: The Microbial Connection. Insects 2023, 14, 89. https://doi.org/10.3390/insects14010089
Morgan-Richards M, Marshall CJ, Biggs PJ, Trewick SA. Insect Freeze-Tolerance Downunder: The Microbial Connection. Insects. 2023; 14(1):89. https://doi.org/10.3390/insects14010089
Chicago/Turabian StyleMorgan-Richards, Mary, Craig J. Marshall, Patrick J. Biggs, and Steven A. Trewick. 2023. "Insect Freeze-Tolerance Downunder: The Microbial Connection" Insects 14, no. 1: 89. https://doi.org/10.3390/insects14010089
APA StyleMorgan-Richards, M., Marshall, C. J., Biggs, P. J., & Trewick, S. A. (2023). Insect Freeze-Tolerance Downunder: The Microbial Connection. Insects, 14(1), 89. https://doi.org/10.3390/insects14010089