
Wolbachia in Black Spiny Whiteflies and Their New Parasitoid Wasp in Japan: Evidence of the Distinct Infection Status on Aleurocanthus camelliae Cryptic Species Complex


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
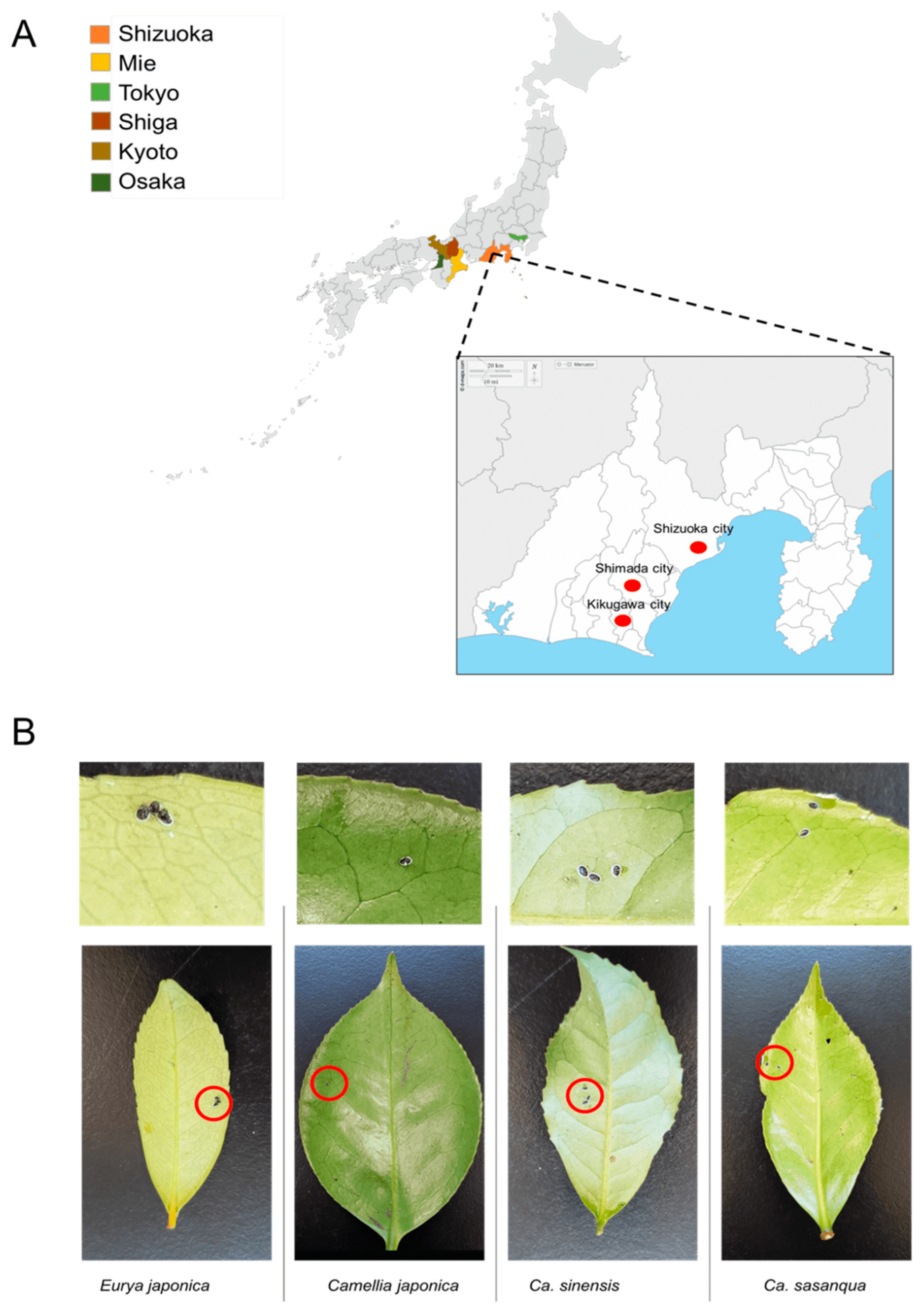
2.1. Sample Collection
2.2. DNA Extraction
2.3. Morphomolecular Identification
2.4. Nested PCR for Determining Wolbachia Infections and MLST Sequencing
2.5. Bacteriophage Detection and Wolbachia Phenotypic Characteristic Determination
2.6. DNA Sequencing and Phylogenetic Analysis
2.7. Genetic Differentiation, Network Analysis, and Recombination Test of Wolbachia
3. Results
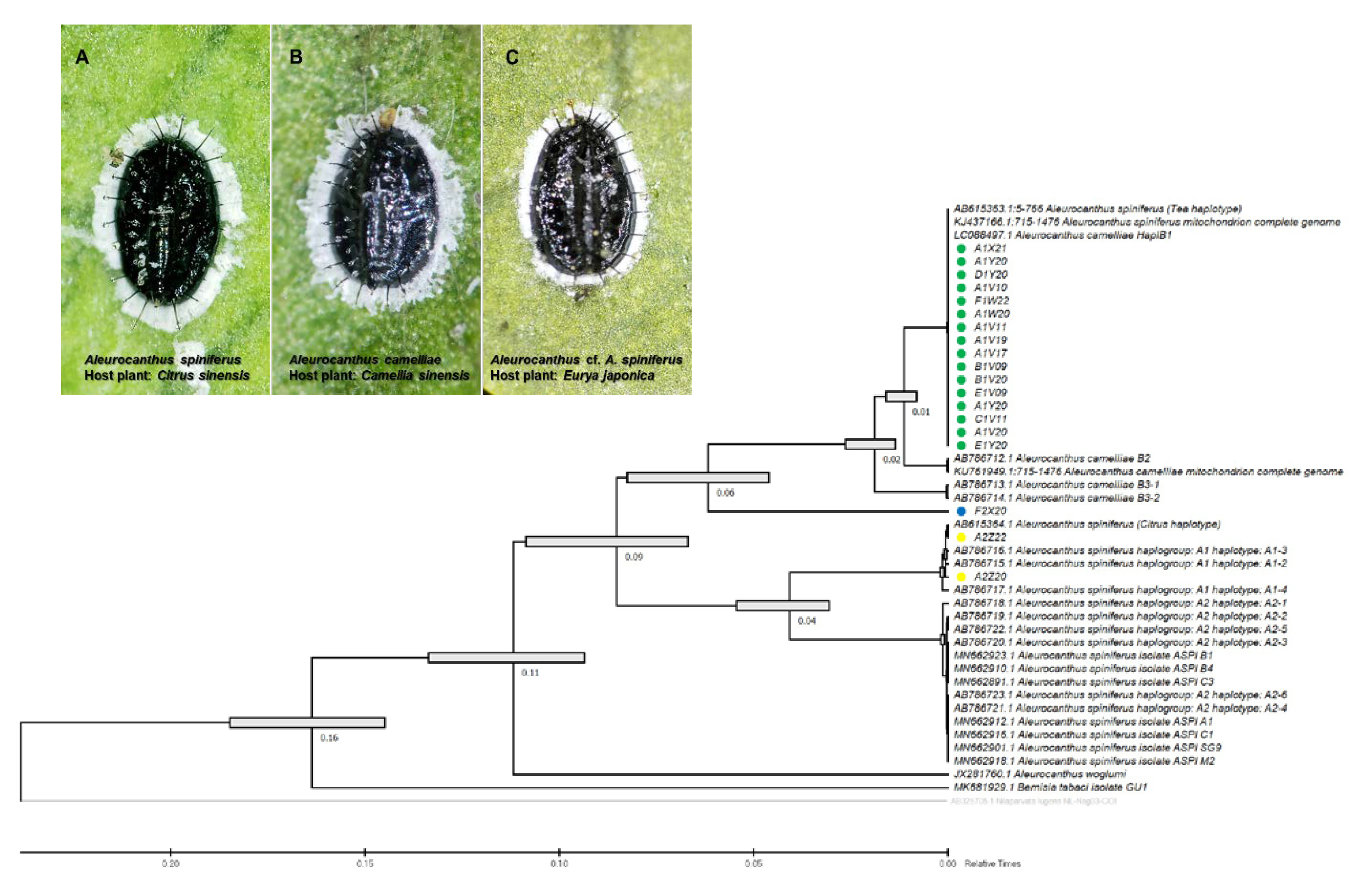
3.1. Morphomolecular Identification
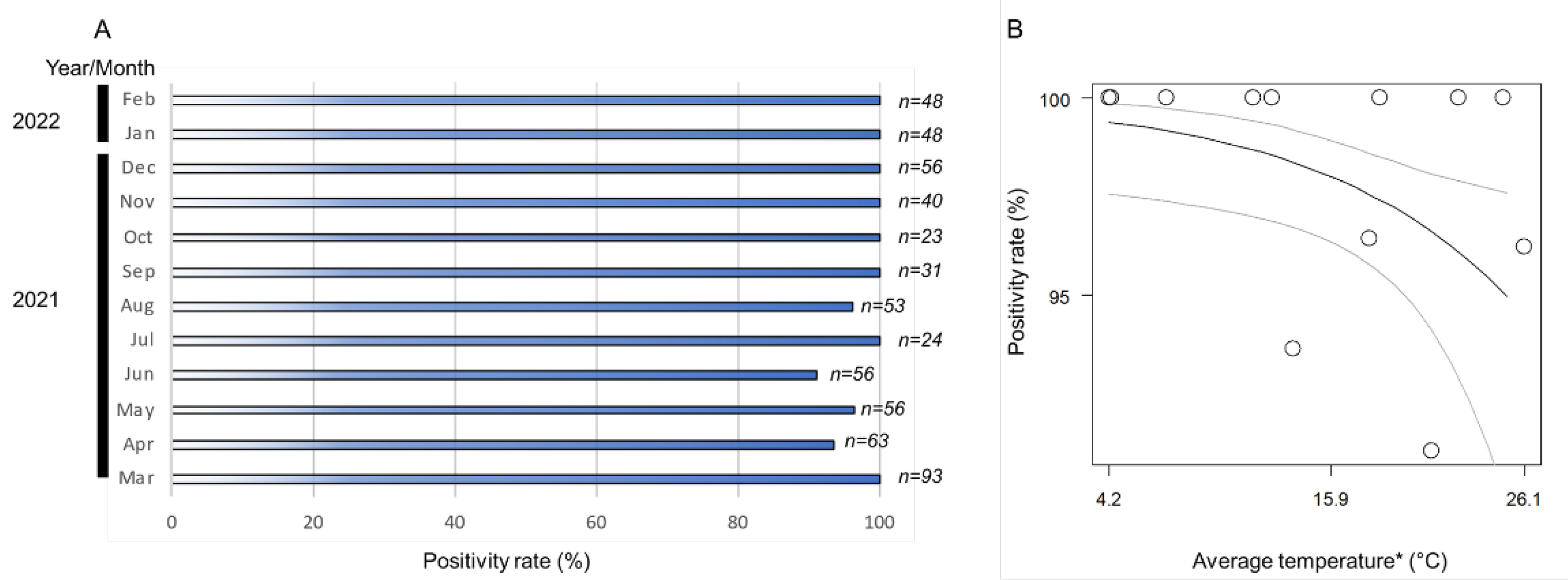
3.2. Positivity and Infection Rates of Wolbachia
3.3. Genetic Diversity of Wolbachia
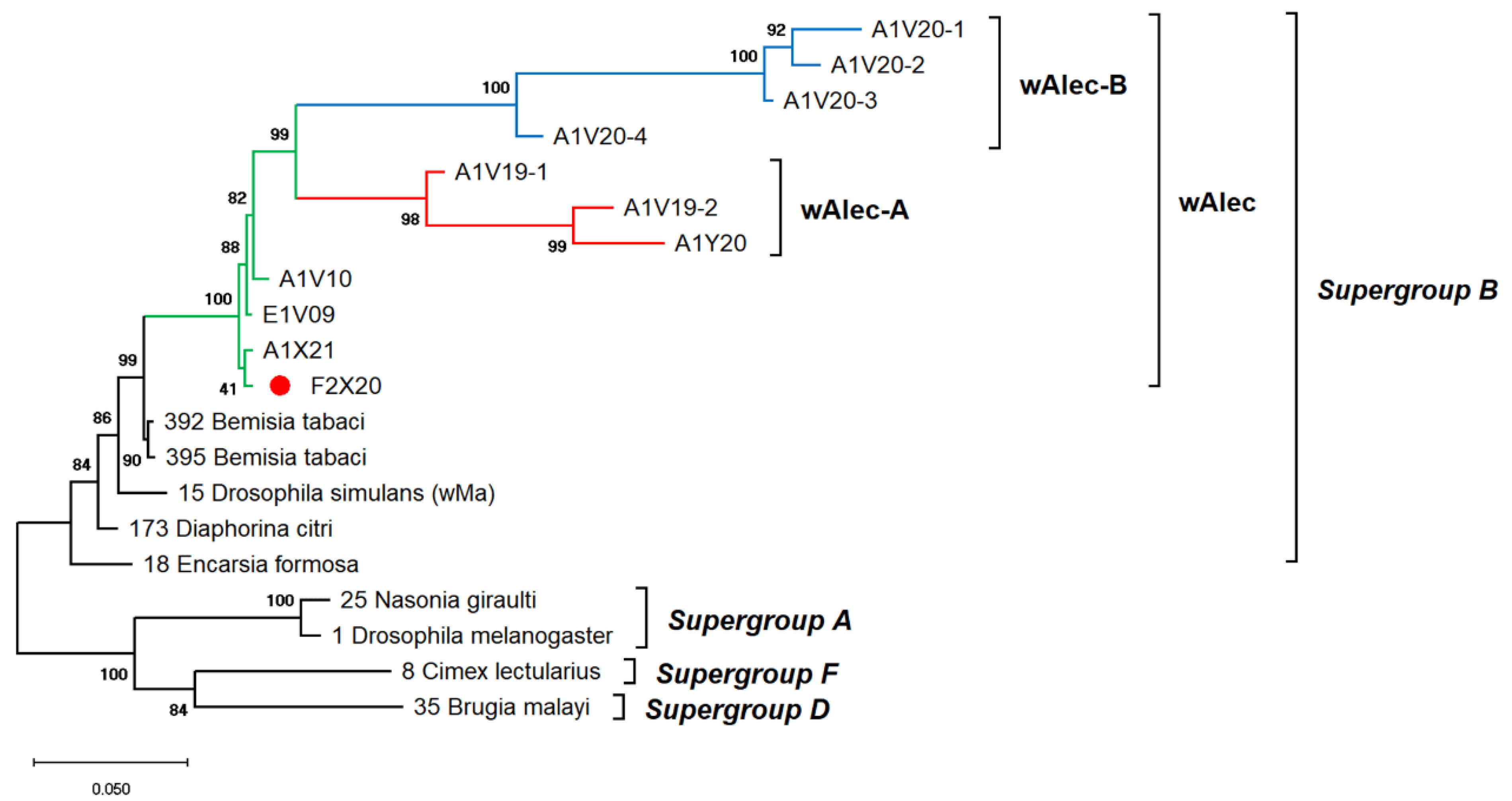
3.4. Phage WO Detection and Wolbachia Phenotypic Screening
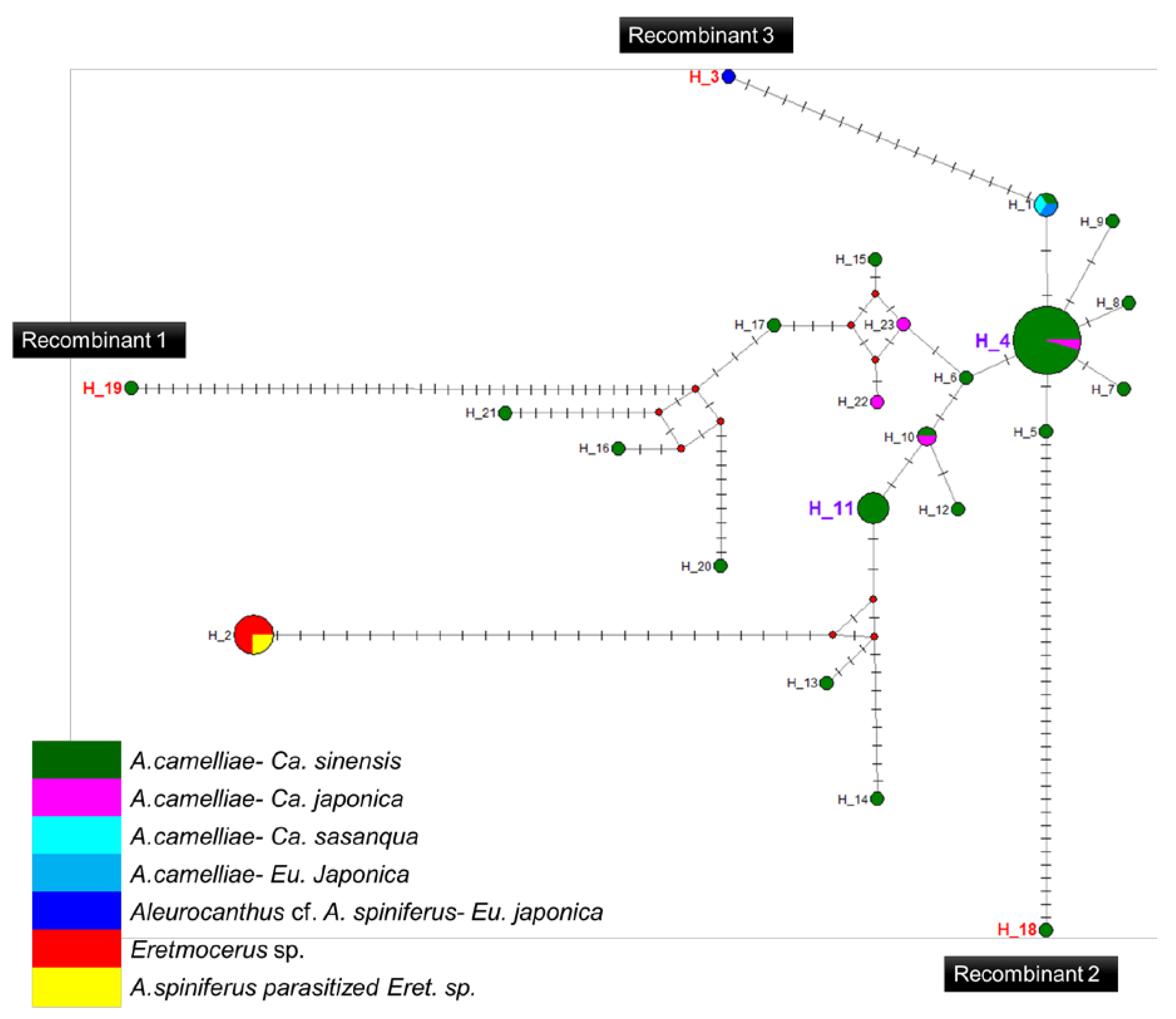
3.5. Recombination and Haplotype Diversity of Wolbachia
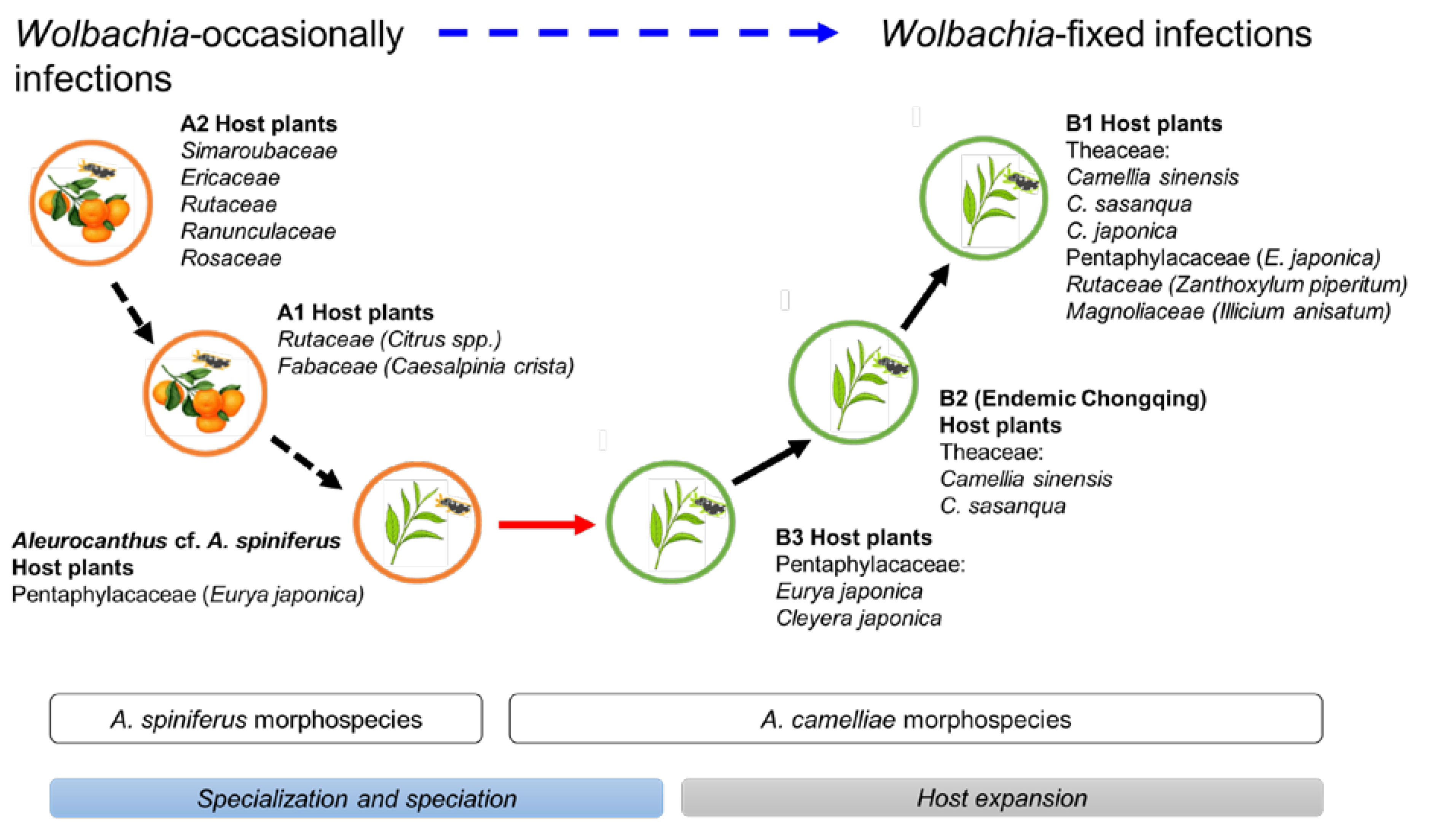
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Werren, J.H. Biology of Wolbachia. Annu. Rev. Entomol. 1997, 42, 587–609. [Google Scholar] [CrossRef] [PubMed]
- Saridaki, A.; Bourtzis, K. Wolbachia: More than just a bug in insects genitals. Curr. Opin. Microbiol. 2010, 13, 67–72. [Google Scholar] [CrossRef]
- Rokas, I. Wolbachia as a speciation agent. Trends Ecol. Evol. 2000, 15, 44–45. [Google Scholar] [CrossRef]
- Gill, A.C.; Darby, A.C.; Makepeace, B.L. Iron necessity: The secret of Wolbachia’s success? PLoS Negl. Trop. Dis. 2014, 8, e3224. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, D.D.; Katju, V.; Jaenike, J. Wolbachia and the evolution of reproductive isolation between Drosophila recens and Drosophila subquinaria. Evolution 1999, 53, 1157–1164. [Google Scholar] [CrossRef]
- Cariou, M.; Duret, L.; Charlat, S. The global impact of Wolbachia on mitochondrial diversity and evolution. J. Evol. Biol. 2017, 30, 2204–2210. [Google Scholar] [CrossRef]
- Bartoňová, A.S.; Konvička, M.; Marešová, J.; Wiemers, M.; Ignatev, N.; Wahlberg, N.; Schmitt, T.; Faltýnek Fric, Z. Wolbachia affects mitochondrial population structure in two systems of closely related Palaearctic blue butterflies. Sci. Rep. 2021, 11, 3019. [Google Scholar] [CrossRef]
- Avtzis, D.N.; Doudoumis, V.; Bourtzis, K. Wolbachia infections and mitochondrial diversity of two Chestnut feeding Cydia species. PLoS ONE 2014, 9, e112795. [Google Scholar] [CrossRef]
- Xiao, J.H.; Wang, N.X.; Murphy, R.W.; Cook, J.; Jia, L.Y.; Huang, D.W. Wolbachia infection and dramatic intraspecific mitochondrial DNA divergence in a Fig wasp. Evolution 2012, 66, 1907–1916. [Google Scholar] [CrossRef] [PubMed]
- Schuler, H.; Köppler, K.; Daxböck-Horvath, S.; Rasool, B.; Krumböck, S.; Schwarz, D.; Hoffmeister, T.S.; Schlick-Steiner, B.C.; Steiner, F.M.; Telschow, A.; et al. The hitchhiker’s guide to Europe: The infection dynamics of an ongoing Wolbachia invasion and mitochondrial selective sweep in Rhagoletis cerasi. Mol. Ecol. 2016, 25, 1595–1609. [Google Scholar] [CrossRef]
- Kodama, Y. Studies on Aleurocanthus spiniferus Quaint; Kagoshima Prefectural Office of Internal Affairs: Kagoshima, Japan, 1931. (In Japanese) [Google Scholar]
- Clausen, C.P. Introduced Parasites and Predators of Arthropod Pests and Weeds: A World Review; US Department of Agriculture Handbook No. 480; Agricultural Research Service, US Department of Agriculture: Washington, DC, USA, 1978; ISBN 001-000-03739-1.
- Lu, M.; Hulcr, J.; Sun, J. The role of symbiotic microbes in insect invasions. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 487–505. [Google Scholar] [CrossRef]
- Xue, X.; Li, S.J.; Ahmed, M.Z.; de Barro, P.J.; Ren, S.X.; Qiu, B.L. Inactivation of Wolbachia reveals its biological roles in whitefly host. PLoS ONE 2012, 7, e48148. [Google Scholar] [CrossRef] [PubMed]
- Kanmiya, K.; Ueda, S.; Kasai, A.; Yamashita, K.; Sato, Y.; Yoshiyasu, Y. Proposal of new specific status for tea-infesting populations of the nominal Citrus spiny whitefly Aleurocanthus spiniferus (Homoptera: Aleyrodidae). Zootaxa 2011, 2797, 25–44. [Google Scholar] [CrossRef]
- Jansen, M.; Porcelli, F. Aleurocanthus camelliae (Hemiptera: Aleyrodidae), a species possibly new for the European fauna of a genus in great need of revision. Tijdschr. Entomol. 2018, 161, 63–78. [Google Scholar] [CrossRef]
- Rizzo, D.; Suma, P.; Rossi, E.; Farina, P.; da Lio, D.; Bartolini, L.; Salemi, C.; Farina, A.; Rapisarda, C. First record of Aleurocanthus camelliae Kanmiya & Kasai, 2011 (Hemiptera, Aleyrodidae) from Italy, on Ornamental Camellia spp. plants. EPPO Bull. 2021, 51, 333–339. [Google Scholar] [CrossRef]
- Adi, M.; Susanti, D. Short communication: First record of Aleurocanthus camelliae (Homoptera: Aleyrodidae) in Indonesia, an invasive pest on various medicinal plants. TOI 2020, 13, 94–100. [Google Scholar] [CrossRef]
- Uesugi, R.; Sato, Y.; Han, B.Y.; Huang, Z.D.; Yara, K.; Furuhashi, K. Molecular evidence for multiple phylogenetic groups within two species of invasive Spiny whiteflies and their parasitoid wasp. Bull. Entomol. Res. 2016, 106, 328–340. [Google Scholar] [CrossRef]
- Nugnes, F.; Laudonia, S.; Jesu, G.; Jansen, M.G.M.; Bernardo, U.; Porcelli, F. Aleurocanthus spiniferus (Hemiptera: Aleyrodidae) in some European countries: Diffusion, hosts, molecular characterization, and natural enemies. Insects 2020, 11, 42. [Google Scholar] [CrossRef]
- Cioffi, M.; Cornara, D.; Corrado, I.; Gerardus, M.; Jansen, M.; Porcelli, F. The status of Aleurocanthus spiniferus from its unwanted introduction in Italy to date. Bull. Insectol. 2013, 66, 273–281. [Google Scholar]
- Kasai, A.; Yamashita, K.; Yoshiyasu, Y. Tea-infesting population of the Citrus spiny whitefly, Aleurocanthus spiniferus (Homoptera: Aleyrodidae), does not accept Citrus leaves as host plants. Jpn. J. Appl. Entomol. Zool. 2010, 54, 140–143. [Google Scholar] [CrossRef]
- Uesugi, R.; Sato, Y. Differentiation of the Tea-infesting population of Citrus spiny whitefly Aleurocanthus spiniferus (Homoptera: Aleyrodidae) from the Citrus-infesting population in Japan on the basis of differences in the mitochondrial Cytochrome c oxidase subunit I gene. Jpn. J. Appl. Entomol. Zool. 2011, 55, 155–161. [Google Scholar] [CrossRef][Green Version]
- Pandey, N.; Singh, A.; Rana, V.S.; Rajagopal, R. Molecular characterization and analysis of bacterial diversity in Aleurocanthus woglumi (Hemiptera: Aleyrodidae). Environ. Entomol. 2013, 42, 1257–1264. [Google Scholar] [CrossRef] [PubMed]
- Bubici, G.; Prigigallo, M.I.; Garganese, F.; Nugnes, F.; Jansen, M.; Porcelli, F. First report of Aleurocanthus spiniferus on Ailanthus altissima: Profiling of the insect microbiome and MicroRNAs. Insects 2020, 11, 161. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Takatsuka, J.; Shimazu, M. Characterization of Paecilomyces cinnamomeus from the Camellia whitefly, Aleurocanthus camelliae (Hemiptera: Aleyrodidae), infesting Tea in Japan. J. Invertebr. Pathol. 2012, 110, 14–23. [Google Scholar] [CrossRef][Green Version]
- Truett, G.E.; Heeger, P.; Mynatt, R.L.; Truett, A.A.; Walker, J.A.; Warman, M.L. Preparation of PCR-quality Mouse genomic DNA with hot sodium hydroxide and tris (HotSHOT). BioTechniques 2000, 29, 52–54. [Google Scholar] [CrossRef]
- Gillespie, P.S. A review of the whitefly Genus Aleurocanthus Quaintance & Baker (Hemiptera: Aleyrodidae) in Australia. Zootaxa 2012, 3252, 1–42. [Google Scholar]
- Folmer, O.F.; Black, M.B.; Hoeh, W.R.; v Lutz, R.; Vrijenhoek, R.C. DNA primers for amplification of mitochondrial Cytochrome c oxidase subunit I from diverse Metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Mol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Zhou, W.; Rousset, F.; O’Neil, S. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proc. Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef]
- Ji, H.L.; Qi, L.D.; Hong, X.Y.; Xie, H.F.; Li, Y.X. Effects of host sex, plant species, and putative host species on the prevalence of Wolbachia in natural populations of Bemisia tabaci (Hemiptera: Aleyrodidae): A modified nested PCR study. J. Econ. Entomol. 2015, 108, 210–218. [Google Scholar] [CrossRef]
- Werren, J.H.; Windsor, D.M. Wolbachia infection frequencies in insects: Evidence of a global equilibrium? Proc. Biol. Sci. 2000, 267, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Baldo, L.; Hotopp, J.C.D.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.Y.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef] [PubMed]
- Masui, S.; Kuroiwa, H.; Sasaki, T.; Inui, M.; Kuroiwa, T.; Ishikawa, H. Bacteriophage WO and virus-like particles in Wolbachia, an endosymbiont of Arthropods. Biochem. Biophys. Res. Commun. 2001, 283, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Su, C.Y.; Zhu, D.H.; Yang, X.H. Design and testing of effective primers for amplification of the orf7 gene of phage WO associated with Andricus hakonensis. Insects 2021, 12, 713. [Google Scholar] [CrossRef]
- LePage, D.P.; Metcalf, J.A.; Bordenstein, S.R.; On, J.; Perlmutter, J.I.; Shropshire, J.D.; Layton, E.M.; Funkhouser-Jones, L.J.; Beckmann, J.F.; Bordenstein, S.R. Prophage WO genes recapitulate and enhance Wolbachia-induced Cytoplasmic incompatibility. Nature 2017, 543, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Pichon, S.; Bouchon, D.; Liu, C.; Chen, L.; Garrett, R.A.; Grève, P. The expression of one ankyrin Pk2 allele of the WO Prophage is correlated with the Wolbachia feminizing effect in Isopods. BMC Microbiol. 2012, 12, 55. [Google Scholar] [CrossRef]
- Walker, T.; Klasson, L.; Sebaihia, M.; Sanders, M.J.; Thomson, N.R.; Parkhill, J.; Sinkins, S.P. Ankyrin repeat do-main-encoding genes in the WPip Strain of Wolbachia from the Culex pipiens Group. BMC Biol. 2007, 5, 39. [Google Scholar] [CrossRef]
- Shropshire, J.D.; On, J.; Layton, E.M.; Zhou, H.; Bordenstein, S.R. One prophage WO gene rescues Cytoplasmic incompatibility in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2018, 115, 4987–4991. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Tamura, K.; Battistuzzi, F.U.; Billing-Ross, P.; Murillo, O.; Filipski, A.; Kumar, S. Estimating divergence times in large molecular phylogenies. Proc. Natl. Acad. Sci. USA 2012, 109, 19333–19338. [Google Scholar] [CrossRef] [PubMed]
- Watterson, G.A. On the number of segregating sites in genetical models without recombination. Theor. Popul. Biol. 1975, 7, 256–276. [Google Scholar] [CrossRef]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; ISBN 9780231886710. [Google Scholar]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Fu, Y.X.; Li, W.H. Statistical tests of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar] [CrossRef]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Tseng, S.P.; Wetterer, J.K.; v Suarez, A.V.; Lee, C.Y.; Yoshimura, T.; Shoemaker, D.W.; Yang, C.S. Genetic diversity and Wolbachia infection patterns in a globally distributed invasive ant. Front. Genet. 2019, 10, 838. [Google Scholar] [CrossRef]
- Martin, D.P.; Varsani, A.; Roumagnac, P.; Botha, G.; Maslamoney, S.; Schwab, T.; Kelz, Z.; Kumar, V.; Murrell, B. RDP5: A computer program for analyzing recombination in, and removing signals of recombination from, nucleotide sequence datasets. Virus Evol. 2021, 7, veaa087. [Google Scholar] [CrossRef]
- Martin, D.; Rybicki, E. RDP: Detection of recombination amongst aligned sequences. Bioinformatics 2000, 16, 562–563. [Google Scholar] [CrossRef]
- Padidam, M.; Sawyer, S.; Fauquet, C.M. Possible emergence of new Gemini viruses by frequent recombination. Virology 1999, 265, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.P.; Posada, D.; Crandall, K.A.; Williamson, C. A modified Bootscan algorithm for automated identification of recombinant sequences and recombination breakpoints. AIDS Res. Hum. Retrovir. 2005, 21, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.M. Analyzing the mosaic structure of genes. J. Mol. Evol. 1992, 34, 126–129. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A. Evaluation of methods for detecting recombination from DNA sequences: Computer simulations. Proc. Natl. Acad. Sci. USA 2001, 98, 13757–13762. [Google Scholar] [CrossRef]
- Gibbs, M.J.; Armstrong, J.S.; Gibbs, A.J. Sister-scanning: A Monte Carlo procedure for assessing signals in recombinant sequences. Bioinformatics 2000, 16, 573–582. [Google Scholar] [CrossRef]
- Weiller, G.F. Phylogenetic Profiles: A graphical method for detecting genetic recombinations in homologous sequences. Mol. Biol. Evol. 1998, 15, 326–335. [Google Scholar] [CrossRef]
- Holmes, E.C.; Worobey, M.; Rambaut, A. Phylogenetic evidence for recombination in dengue virus. Mol. Biol. Evol. 1999, 16, 405–409. [Google Scholar] [CrossRef]
- Lam, H.M.; Ratmann, O.; Boni, M.F. Improved algorithmic complexity for the 3SEQ recombination detection algorithm. Mol. Biol. Evol. 2018, 35, 247–251. [Google Scholar] [CrossRef]
- Martin, J.H.; Mound, L.A. An annotated check list of the World’s whiteflies (Insecta: Hemiptera: Aleyrodidae). Zootaxa 2007, 1492, 1–84. [Google Scholar] [CrossRef]
- Manzari, S.; Quicke, D.L.J. A cladistic analysis of whiteflies, Subfamily Aleyrodinae (Hemiptera: Sternorrhyncha: Aleyrodidae). J. Nat. Hist. 2006, 40, 2423–2554. [Google Scholar] [CrossRef]
- Evans, G. The Whiteflies (Hemiptera: Aleyrodidae) of the World and Their Host Plants and Natural Enemies. 2007. Available online: http://keys.lucidcentral.org/keys/v3/whitefly/PDF_PwP%20ETC/world-whitefly-catalog-Evans.pdf (accessed on 29 June 2022).
- Janz, N.; Nylin, S. The Oscillation Hypothesis of Host-Plant Range and Speciation; Tilmon, K.J., Ed.; University of California Press: London, UK, 2008; pp. 203–215. [Google Scholar]
- Angiosperm Phylogeny Group. An update of the Angiosperm phylogeny group classification for the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 2009, 161, 105–121. [Google Scholar] [CrossRef]
- Miyatake, Y. A list of the whiteflies of Japan with their host plant and distribution data (Homoptera: Aleyrodidae). Rostria 1980, 32, 291–330. (In Japanese) [Google Scholar]
- Narita, S.; Nomura, M.; Kato, Y.; Fukatsu, T. Genetic structure of sibling butterfly species affected by Wolbachia infection sweep: Evolutionary and biogeographical implications. Mol. Ecol. 2006, 15, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Tong, X.; Li, Y.Y.; Sun, Q.; Gao, Y.; Zhang, S.H.; Wang, R.; Chen, X.Y. Presence of cryptic species in host insects forms a hierarchical Wolbachia infection pattern. Entomol. Genet. 2022, 42, 571–578. [Google Scholar] [CrossRef]
- Jaenike, J.; Dyer, K.A.; Cornish, C.; Minhas, M.S. Asymmetrical reinforcement and Wolbachia infection in Drosophila. PLoS Biol. 2006, 4, 325. [Google Scholar] [CrossRef] [PubMed]
- Bruzzese, D.J.; Schuler, H.; Wolfe, T.M.; Glover, M.M.; v Mastroni, J.V.; Doellman, M.M.; Tait, C.; Yee, W.L.; Rull, J.; Aluja, M.; et al. Testing the potential contribution of Wolbachia to speciation when Cytoplasmic incompatibility becomes associated with host-related reproductive isolation. Mol. Ecol. 2022, 31, 2935–2950. [Google Scholar] [CrossRef]
- Bailly-Bechet, M.; Martins-Simões, P.; Szöllosi, G.J.; Mialdea, G.; Sagot, M.F.; Charlat, S. How long does Wolbachia remain on board? Mol. Biol. Evol. 2017, 34, 1183–1193. [Google Scholar] [CrossRef]
- Nikoh, N.; Hosokawa, T.; Moriyama, M.; Oshima, K.; Hattori, M.; Fukatsu, T. Evolutionary origin of insect–Wolbachia nutritional mutualism. Proc. Natl. Acad. Sci. USA 2014, 111, 10257–10262. [Google Scholar] [CrossRef]
- Moran, N.A. Accelerated evolution and Muller’s rachet in endosymbiotic bacteria. Proc. Natl. Acad. Sci. USA 1996, 93, 2873–2878. [Google Scholar] [CrossRef]
- Jiggins, F.M.; von der Schulenburg, J.H.; Hurst, G.D.; Majerus, M.E. Recombination confounds interpretations of Wolbachia evolution. Proc. Biol. Sci. 2001, 268, 1423–1427. [Google Scholar] [CrossRef]
- Awadalla, P. The evolutionary genomics of pathogen recombination. Nat. Rev. Genet. 2003, 4, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, H.; Zhou, R. Genome-wide evidence for positive selection and recombination in Actinobacillus pleuropneumoniae. BMC Evol. Biol. 2011, 11, 203. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.H.; Zhu, D.H.; Liu, Z.; Zhao, L.; Su, C.Y. High levels of multiple infections, recombination, and horizontal transmission of Wolbachia in the Andricus mukaigawae (Hymenoptera; Cynipidae) communities. PLoS ONE 2013, 8, e78970. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.L.; Pepper-Tunick, E.; Svedberg, J.; Byrne, A.; Ruelas Castillo, J.; Vollmers, C.; Beinart, R.A.; Corbett-Detig, R. Horizontal transmission and recombination maintain forever young bacterial symbiont genomes. PLoS Genet. 2020, 16, e1008935. [Google Scholar] [CrossRef]
- Zhou, X.F.; Li, Z.X. Establishment of the Cytoplasmic incompatibility-inducing Wolbachia strain wMel in an important agricultural pest insect. Sci. Rep. 2016, 6, 39200. [Google Scholar] [CrossRef] [PubMed]
- Vavre, F.; Fleury, F.; Lepetit, D.; Fouillet, P.; Boulétreau, M. Phylogenetic evidence for horizontal transmission of Wolbachia in host-parasitoid associations. Mol. Biol. Evol. 1999, 16, 1711–1723. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Li, S.J.; Xue, X.; Yin, X.J.; Ren, S.X.; Jiggins, F.M.; Greeff, J.M.; Qiu, B.L. The intracellular bacterium Wolbachia uses parasitoid wasps as phoretic vectors for efficient horizontal transmission. PLoS Pathog. 2015, 10, e1004672. [Google Scholar] [CrossRef]
- Johannesen, J. Tracing the history and ecological context of Wolbachia double infection in a specialist host (Urophora cardui)—Parasitoid (Eurytoma serratulae) system. Ecol. Evol. 2017, 7, 986–996. [Google Scholar] [CrossRef]
- DeBach, P. Biological Control of Insect Pests and Weeds; Reihold: New York, NY, USA, 1964; p. 676. [Google Scholar]
- Ozawa, A.; Uchiyama, T. Parasitism of the tea spiny whitefly, Aleurocanthus camelliae Kanmiya & Kasai, by Encarsia smithi (Silvestri) in tea fields in Shizuoka Prefecture in 2012, two years after the first identification of the pest. Ann. Rept. Kansai Pl. Prot. 2013, 55, 89–91. [Google Scholar]
- Uesugi, R.; Yara, K.; Sato, Y. Changes in population density of Aleurocanthus camelliae (Hemiptera: Aleyrodidae) and parasitism rate of Encarsia smithi (Hymenoptera: Aphelinidae) during the early invasion stages. Appl. Entomol. Zool. 2016, 51, 581–588. [Google Scholar] [CrossRef]
- Kuwana, I. Notes on a newly imported parasite of the Spiny whitefly attacking Citrus in Japan. In Proceedings of the fifth Pacific Science Congress Organized by the Pacific Science Association and the National Research Council of Canada, Victoria and Vancouver, BC, Canada, 1–14 June 1933; University of Toronto: Toronto, ON, Canada, 1934; pp. 3521–3525. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Whitefly Species | Haplotype | Locality (Prefecture) | Host Plant | Year | Label Isolate * |
|---|---|---|---|---|---|
| A. camelliae | B1 | Shizuoka | C. sinensis | 2010 | A1V10 |
| B1 | Shizuoka | C. sinensis | 2011 | A1V11 | |
| B1 | Shizuoka | C. sinensis | 2017 | A1V17 | |
| B1 | Shizuoka | C. sinensis | 2018 | A1V18 | |
| B1 | Shizuoka | C. sinensis | 2019 | A1V19 | |
| B1 | Shizuoka | C. sinensis | 2020 | A1V20 | |
| B1 | Shizuoka | C. japonica | 2020 | A1W20 | |
| B1 | Shizuoka | E. japonica | 2021 | A1X21 | |
| B1 | Shizuoka | C. sasanqua | 2020 | A1Y20 | |
| B1 | Shiga | C. sinensis | 2009 | B1V09 | |
| B1 | Shiga | C. sinensis | 2020 | B1V20 | |
| B1 | Mie | C. sinensis | 2011 | C1V11 | |
| B1 | Osaka | C. sasanqua | 2020 | D1Y20 | |
| B1 | Kyoto | C. sinensis | 2009 | E1V09 | |
| B1 | Kyoto | C. sinensis | 2020 | E1V20 | |
| B1 | Tokyo | E.japonica | 2022 | F1W22 | |
| A. spiniferus | A1 | Shizuoka | Ci.sinensis | 2020 | A2Z20 a |
| A1 | Shizuoka | Ci.sinensis | 2021 | A2Z21 | |
| A1 | Shizuoka | Ci.sinensis | 2022 | A2Z22 | |
| ? | Tokyo | E. japonica | 2020 | F2X20 | |
| P. euryae | Shizuoka | E. japonica | 2021 | A3X21 | |
| Shizuoka | C. sinensis | 2020 | A3V20 | ||
| Kyoto | C. sinensis | 2020 | E3V20 |
| Gene | Isolates | Type a | Close Relative | % Similarity | Source | Infection |
|---|---|---|---|---|---|---|
| COI-1 | A1V20-1 | B1 | Encarsia sp. | 90.94 | KJ444561.1 | (+) |
| A1W20-1 | B1 | Encarsia inquirenda | 92.74 | MH928989.1 | (+) | |
| A1W20-2 | B1 | Encarsia perniciosi | 90.28 | JQ083717.1 | (+) | |
| A1W20-3 | B1 | A. spiniferus | 99.38 | KJ437166.1 | (+) | |
| D1Y20 | B1 | A. spiniferus | 99.53 | KJ437166.1 | (+) | |
| B1V20 | B1 | Encarsia obtusiclava | 90.17 | MG813798.1 | (−) | |
| F2X20-1 | - | Aleurocanthus sp. | 81.75 | KY835557.1 | (−) | |
| F2X20-2 | - | Aleurocanthus sp. | 81.95 | KY836994.1 | (−) | |
| A1V20-2 | B1 | Eretmocerus orchamoplati | 88.78 | JF750712.1 | (+) | |
| A2Z20-1 b | - | E. orchamoplati | 84.62 | JF750714.1 | (+) | |
| A2Z20-2 b | - | Aleurocanthus arecae | 83.18 | MZ301225.1 | (−) | |
| COI-2 | F2X20-4 | - | E. smithi type 2 | 99.46 | AB786724.1 | (−) |
| F2X20-3 | - | E. smithi type 1 | 97.82 | AB786726.1 | (+) | |
| F2X20-5 | - | T. acaciae | 80.72 | MT901108.1 | (−) | |
| A2Z21-1 | - | E. smithi type 1 | 99.32 | AB786726.1 | (−) | |
| A2Z21-2 | - | E. smithi type 1 | 99.32 | AB786726.1 | (−) | |
| A1V20-3 | B1 | E. smithi type 1 | 99.57 | AB786726.1 | (+) | |
| A1V20-4 | B1 | E. smithi type 1 | 98.29 | AB786726.1 | (+) | |
| COB | F2X20-3 | - | Encarsia formosa | 86.44 | MG813797.1 | (+) |
| F2X20-4 | - | E. formosa | 86.49 | MG813797.1 | (−) | |
| A1V20-5 | B1 | E. formosa | 86.39 | MG813797.1 | (+) | |
| A1V20-6 | B1 | Eretmocerus sp. | 84.89 | KX714964.1 | (+) | |
| A1V20-7 | B1 | E. formosa | 85.91 | MG813797.1 | (+) | |
| A1X21 | B1 | Eretmocerus sp. | 85.16 | KX714964.1 | (+) | |
| A2Z21-3 | - | E. formosa | 86.57 | MG813797.1 | (−) | |
| A2Z21-4 | - | E. formosa | 86.26 | MG813797.1 | (−) |
| Species | Host | No. Samples Assessed | mtCOI Gene of Host Amplification | Positive Infection (Nested PCR) | Infection Rate c (%) |
|---|---|---|---|---|---|
| A. camelliae | C. sinensis | 738 | 728 | 703 | 96.5 |
| C. sasanqua | 30 | 30 | 2 | 6.7 | |
| C. japonica | 15 | 15 | 6 | 40 | |
| E. japonica | 1 | 1 | 1 | 100 a | |
| A. spiniferus | C.sinensis | 104 | 103 | 2 | 1.9 |
| Aleurocanthus cf. A. spiniferus | E. japonica | 40 | 40 | 1 | 2.5 |
| E. smithi | A. spiniferus | 16 | 16 | 0 | 0 |
| Eretmocerus | A. camelliae | 7 | 7 | 7 | 100 |
| A. spiniferusb | 1 | 1 | 1 | 100 | |
| Total | 952 | 941 | 722 |
| Gene | Sample Pool | N | S | h | Molecular Diversity Indices | Neutrality Tests | |||
|---|---|---|---|---|---|---|---|---|---|
| Hd | π | k | Tajima’s D (P) | Fu and Li’s F (P) | |||||
| wsp | A. camelliae populations | 30 | 5 | 3 | 0.1 | 0.00099 | 0.33 | −2.00763 (<0.05) * | −3.34142 (<0.02) ** |
| Associated populations * | 8 | 122 | 8 | 1.0 | 0.13692 | 46.14 | −0.60085 (>0.10) ns | −0.61175 (>0.10) ns | |
| 16S rRNA | A. camelliae populations | 51 | 85 | 21 | 0.8 | 0.02292 | 7.71 | −2.31567 (<0.01) ** | −3.93027 (<0.02) ** |
| Associated populations * | 9 | 36 | 2 | 0.2 | 0.02026 | 7.33 | −1.99788 (<001) ** | −2.48500 (<0.02) ** | |
| Isolates | Sequence Dissimilarity | Phenotypic Screening | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | pk1a | pk1b | pk2b1 | pk2b2 | cifA | cifB | ||
| 1 | A1V20-1 | 0.000 | 0.004 | 0.004 | (−) | (−) | (−) | (−) | (−) | (−) | |
| 2 | A1V20-2 | 0.000 | 0.004 | 0.004 | (−) | (−) | (−) | (−) | (−) | (−) | |
| 3 | F2X20 | 0.006 | 0.006 | 0.004 | (−) | (−) | (−) | (−) | (−) | (−) | |
| 4 | A1X21 | 0.006 | 0.006 | 0.006 | (−) | (−) | (−) | (−) | (−) | (−) | |
| Gene | No. Events a | Putative Recombination b | Major Parent c (% Similarity) | Minor Parent d (% Similarity) | Analysis | GENECONV | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R | G | B | M | C | S | P | L | 3S | Start | End | |||||
| wsp | 1 | E3V20 | E1V20 (82.6) | Unknown | (−) | + | (−) | + | + | (−) | + | (−) | (−) | 94 | 254 |
| 16S rRNA | 2 | A1V20-10 | A1V20-13 (96.3) | Unknown | (−) | + | (−) | + | + | + | + | (−) | (−) | 284 | 372 |
| 3 | A1V20-27 | A1V20-13 (96.7) | Unknown | + | + | (−) | (−) | + | (−) | + | (−) | (−) | 237 | 376 | |
| 4 | F2X20 | A1V20-13 (95.2) | Unknown | + | + | (−) | + | + | (−) | + | (−) | (−) | 288 | 380 | |
| MLST | 5 | A1V20-3 | A1V19-1 (94) | Unknown | + | + | + | + | + | (−) | + | (−) | (−) | 409 | 755 |
| 6 | A1V20-2 | A1V19-1 (94.2) | Unknown | + | + | + | + | + | (−) | + | (−) | (−) | 396 | 764 | |
| 7 | A1V20-4 | A1V19-2 (93.1) | Drosophila simulans (89.4) | + | + | + | + | + | + | + | (−) | (−) | 1 | 362 | |
| 8 | A1V20-1 | A1V19-2 (96.5) | D. simulans (97) | + | + | + | + | + | + | + | (−) | (−) | 131 | 361 | |
| 9 | A1V19-2 | A1Y20 (94.7) | A1V20-4 (99.8) | + | + | + | + | + | + | + | (−) | (−) | 1250 | ∞~ | |
| 10 | A1V19-1 | A1Y20 (95.2) | Brugia malayi (92.8) | + | + | (−) | + | + | (−) | + | (−) | (−) | ∞~ | 628 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andrianto, E.; Kasai, A. Wolbachia in Black Spiny Whiteflies and Their New Parasitoid Wasp in Japan: Evidence of the Distinct Infection Status on Aleurocanthus camelliae Cryptic Species Complex. Insects 2022, 13, 788. https://doi.org/10.3390/insects13090788
Andrianto E, Kasai A. Wolbachia in Black Spiny Whiteflies and Their New Parasitoid Wasp in Japan: Evidence of the Distinct Infection Status on Aleurocanthus camelliae Cryptic Species Complex. Insects. 2022; 13(9):788. https://doi.org/10.3390/insects13090788
Chicago/Turabian StyleAndrianto, Eko, and Atsushi Kasai. 2022. "Wolbachia in Black Spiny Whiteflies and Their New Parasitoid Wasp in Japan: Evidence of the Distinct Infection Status on Aleurocanthus camelliae Cryptic Species Complex" Insects 13, no. 9: 788. https://doi.org/10.3390/insects13090788
APA StyleAndrianto, E., & Kasai, A. (2022). Wolbachia in Black Spiny Whiteflies and Their New Parasitoid Wasp in Japan: Evidence of the Distinct Infection Status on Aleurocanthus camelliae Cryptic Species Complex. Insects, 13(9), 788. https://doi.org/10.3390/insects13090788