

Behavior and Bioadhesives: How Bolas Spiders, Mastophora hutchinsoni, Catch Moths


Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Field Measurements
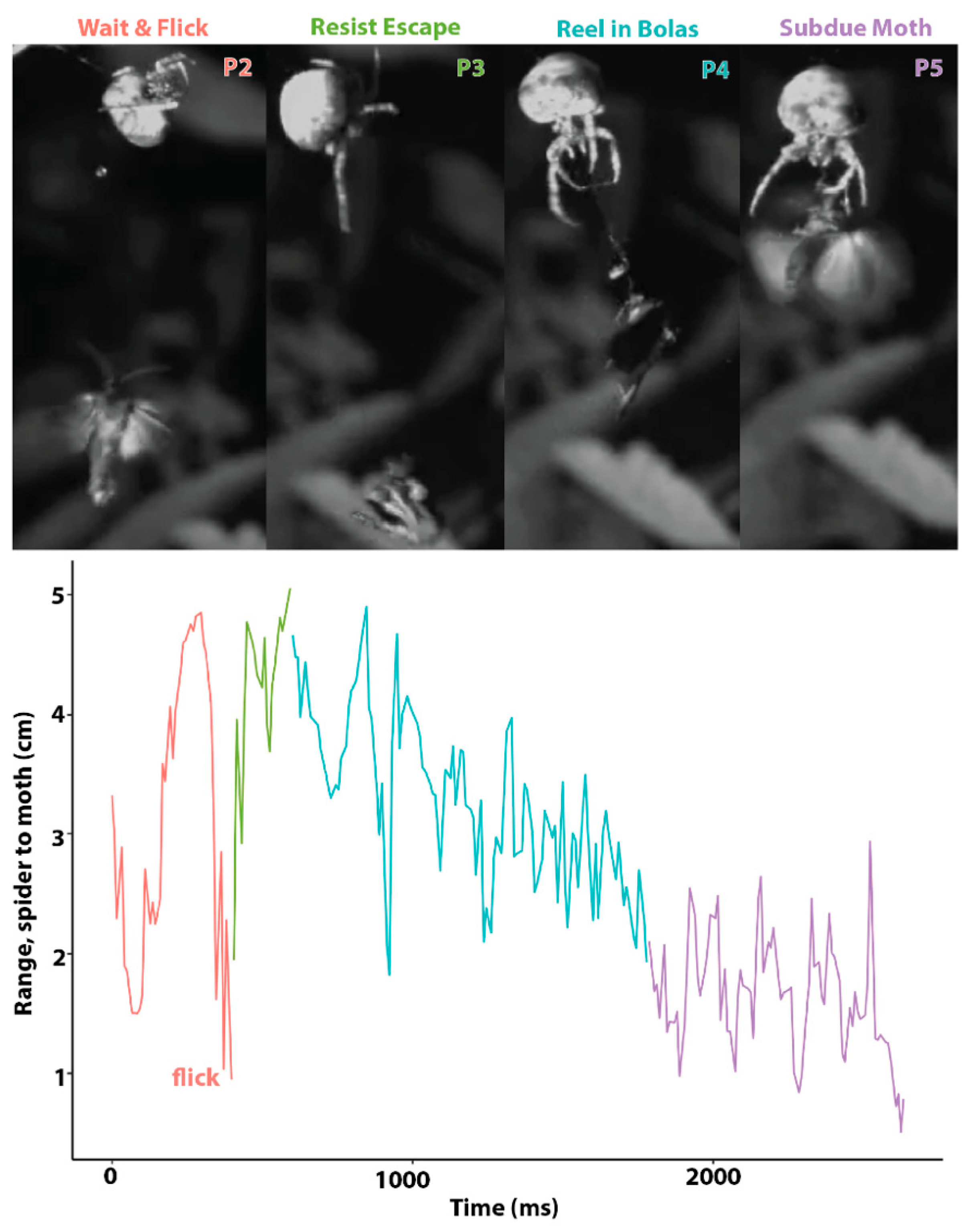
2.2. Kinematics of Prey Capture
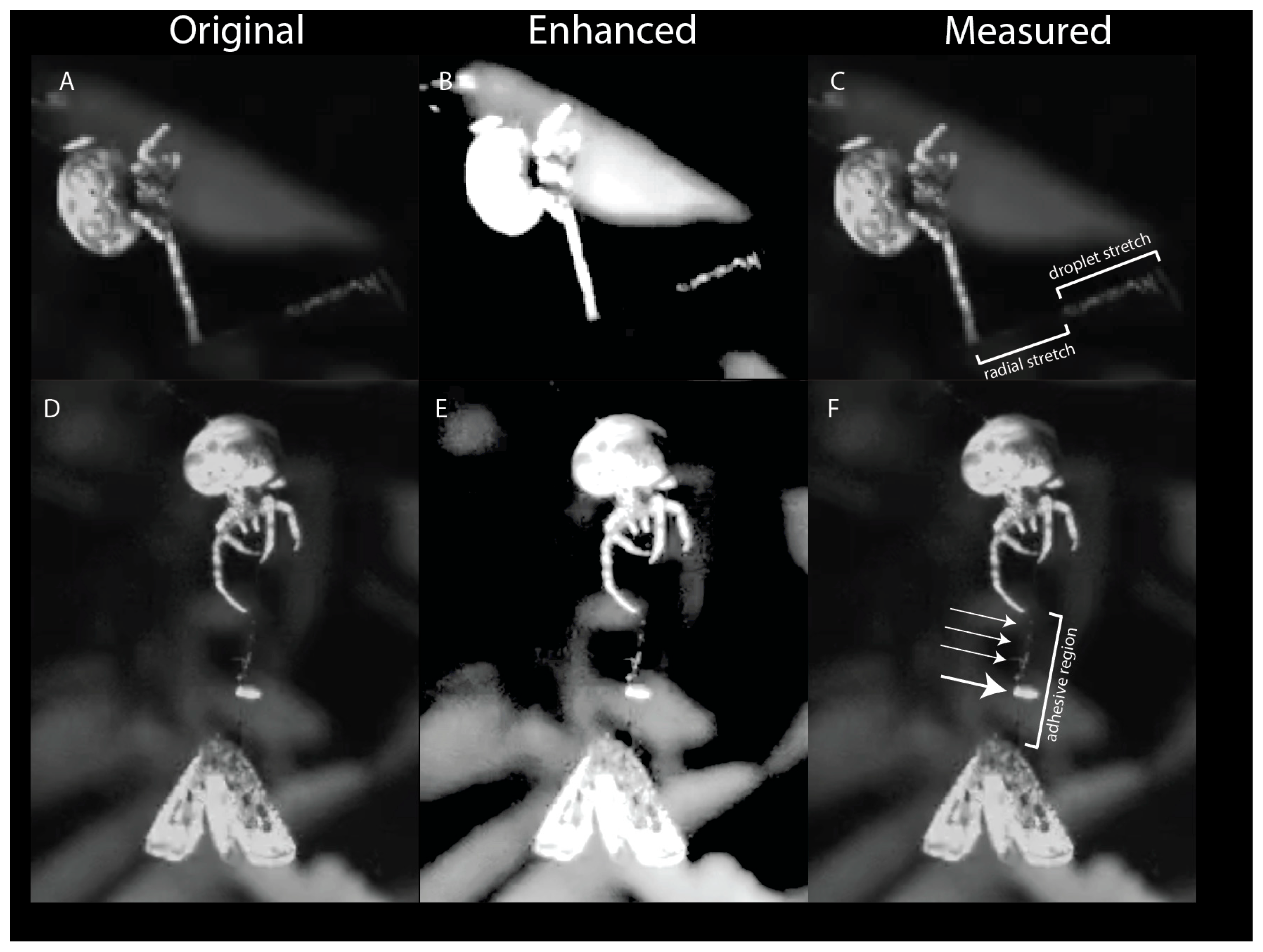
2.3. Material Properties of the Droplet
3. Results
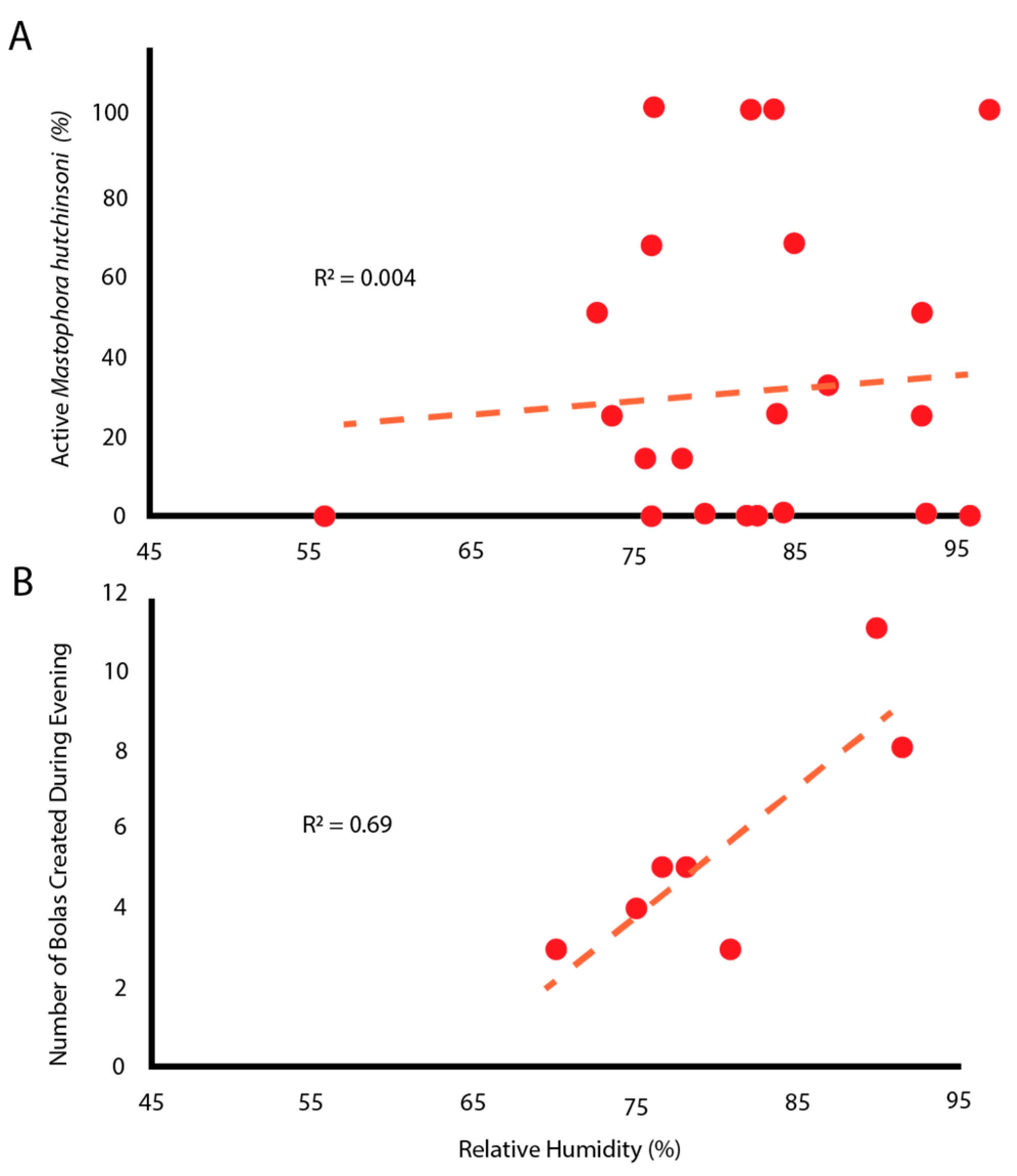
3.1. Observations on Bolas Building Behavior
3.2. Kinematics of Prey Capture
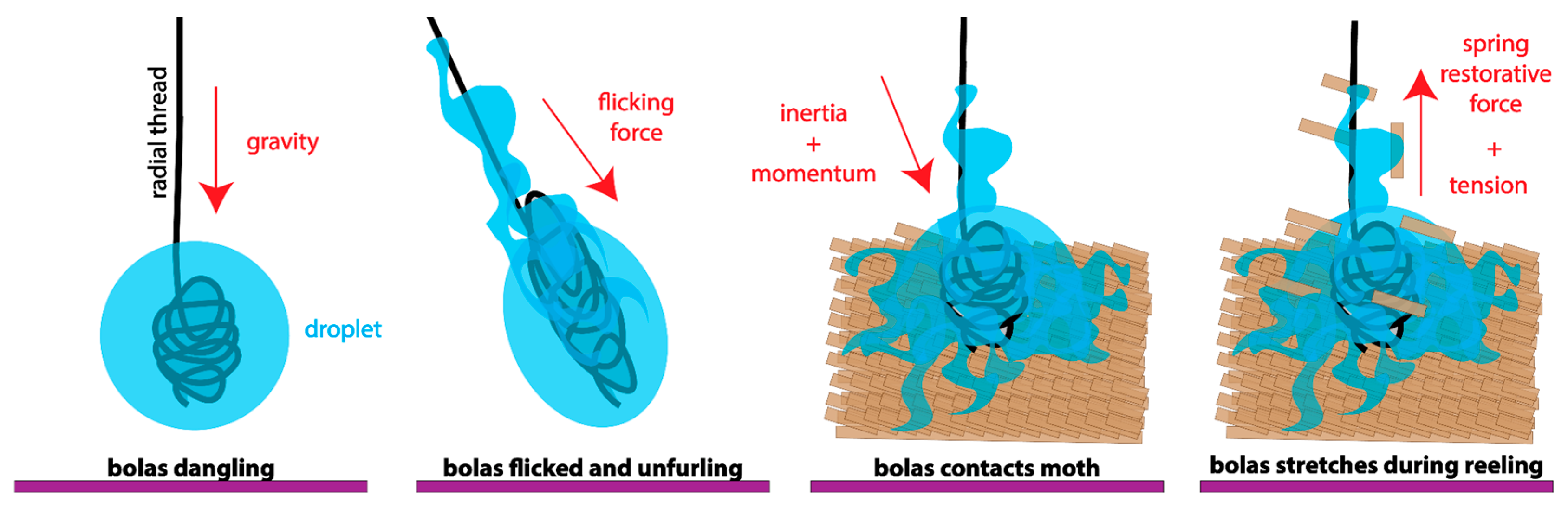
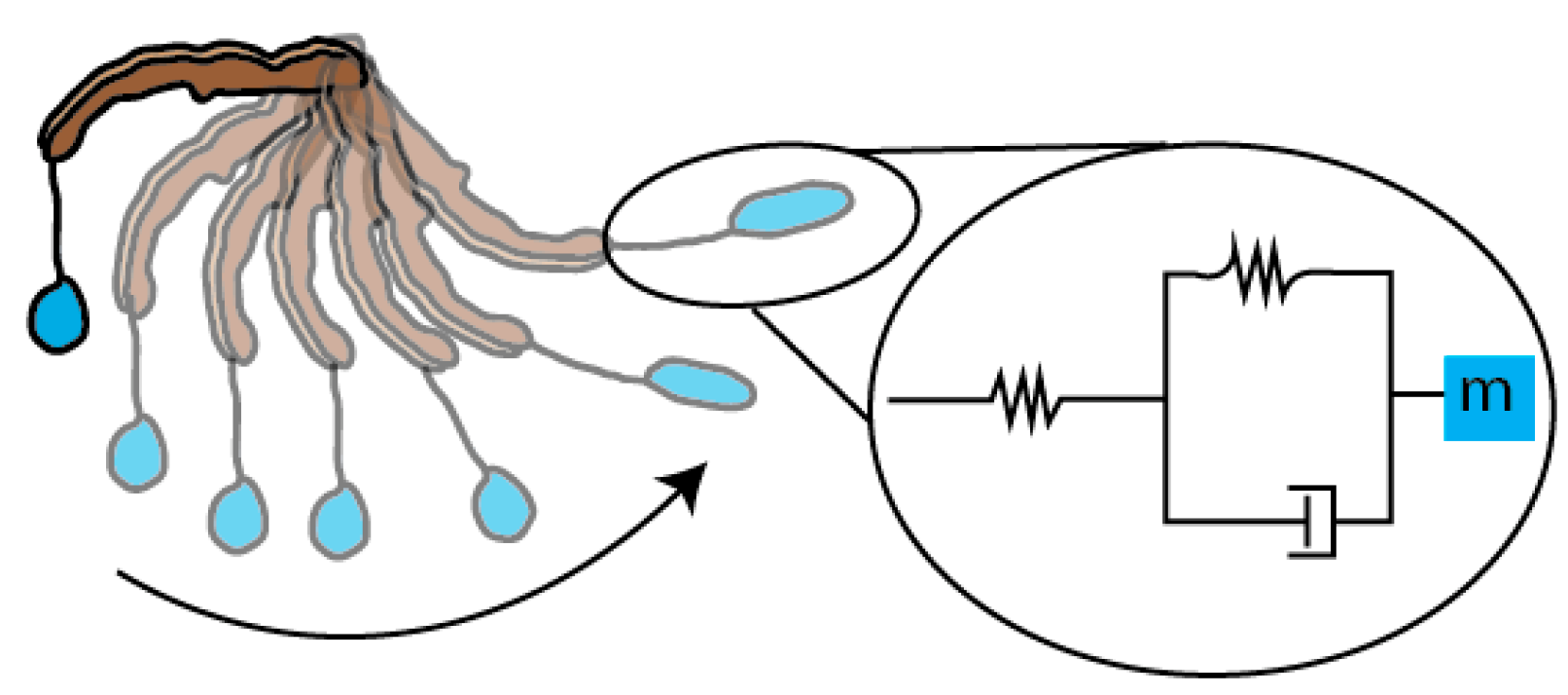
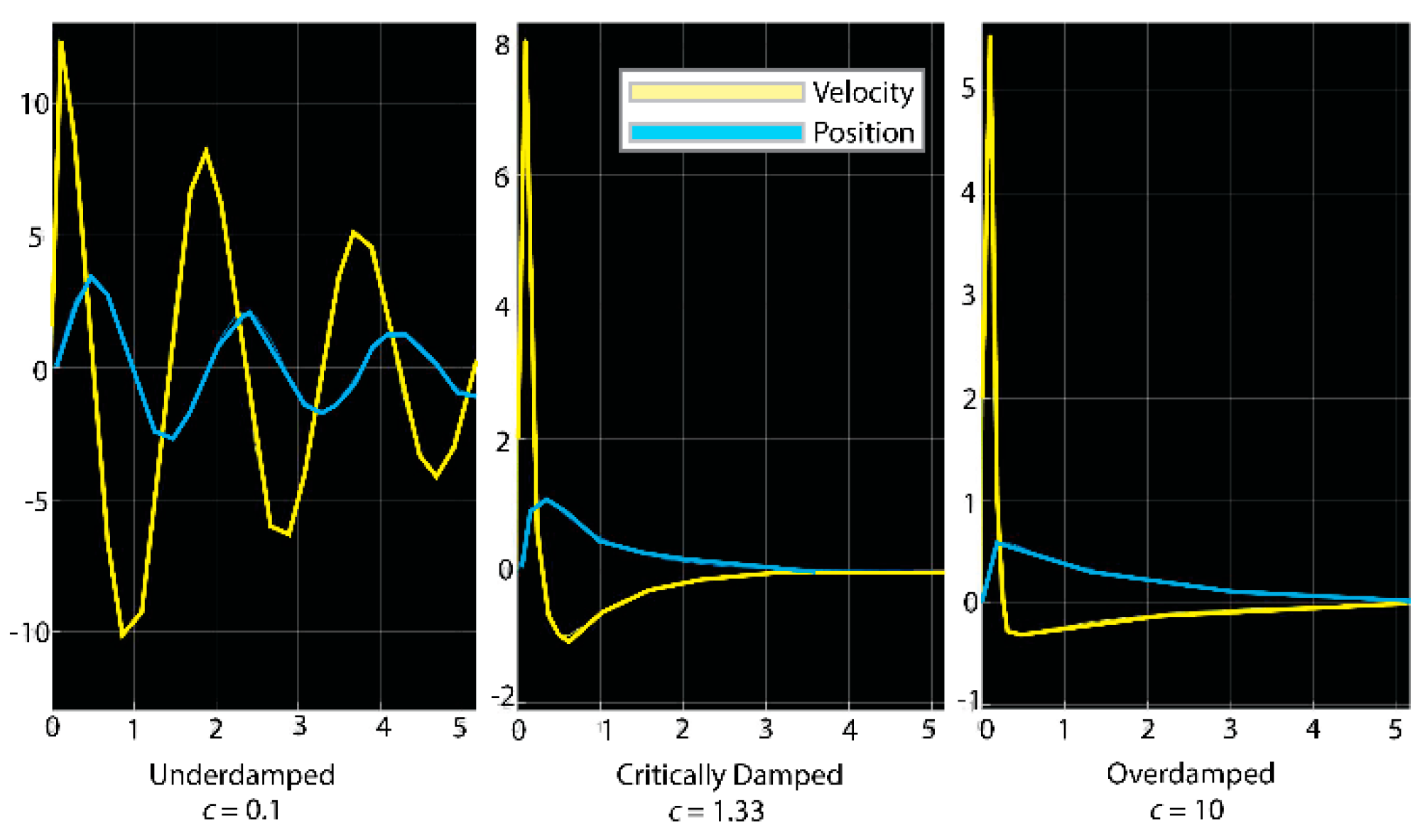
3.3. Model of Bolas as Viscoelastic Spring during Prey Capture
4. Discussion
4.1. Inquisitive Prey
4.2. Environmentally Constrained Predators
4.3. Predator–Prey Interactions via a Viscoelastic Bolas
4.4. Adaptations for the Capture of Moths
4.5. Caveats and Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foelix, R.F. Biology of Spiders; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Blackledge, T.A.; Scharff, N.; Coddington, J.A.; Szüts, T.; Wenzel, J.W.; Hayashi, C.Y.; Agnarsson, I. Reconstructing web evolution and spider diversification in the molecular era. Proc. Natl. Acad. Sci. USA 2009, 106, 5229–5234. [Google Scholar] [CrossRef] [PubMed]
- Stowe, M.K. Prey specialization in the Araneidae. In Spiders Webs Behavior and Evolution; Stanford University Press: Stanford, CA, USA, 1986; pp. 101–131. [Google Scholar]
- Diaz, C.; Maksuta, D.; Amarpuri, G.; Tanikawa, A.; Miyashita, T.; Dhinojwala, A.; Blackledge, T.A. The moth specialist spider Cyrtarachne akirai uses prey scales to increase adhesion. J. R. Soc. Interfaces 2020, 17, 20190792. [Google Scholar] [CrossRef] [PubMed]
- Andersen, S.O. Biochemistry of Insect Cuticle. Annu. Rev. Entomol. 1979, 24, 29–59. [Google Scholar] [CrossRef]
- Miyashita, T.; Sakamaki, Y.; Shinkai, A. Evidence against moth attraction by Cyrtarachne, a genus related to bolas spiders. Acta Arachnol. 2001, 50, 1–4. [Google Scholar] [CrossRef]
- Tanikawa, A. Two new species of the genus Cyrtarachne (Araneae: Araneidae) from Japan hitherto identified as C. inaequalis. Acta Arachnol. 2013, 62, 95–101. [Google Scholar] [CrossRef][Green Version]
- Cartan, C.K.; Miyashita, T. Extraordinary web and silk properties of Cyrtarachne (Araneae, Araneidae): A possible link between orb-webs and bolas. Biol. J. Linn. Soc. 2000, 71, 219–235. [Google Scholar] [CrossRef]
- Baba, Y.G.; Kusahara, M.; Maezono, Y.; Miyashita, T. Adjustment of web-building initiation to high humidity: A constraint by humidity-dependent thread stickiness in the spider Cyrtarachne. Naturwissenschaften 2014, 101, 587–593. [Google Scholar] [CrossRef]
- Diaz, C.; Tanikawa, A.; Miyashita, T.; Dhinojwala, A.; Blackledge, T.A. Silk structure rather than tensile mechanics explains web performance in the moth-specialized spider, Cyrtarachne. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2018, 329, 120–129. [Google Scholar] [CrossRef]
- Diaz, C.; Tanikawa, A.; Miyashita, T.; Amarpuri, G.; Jain, D.; Dhinojwala, A.; Blackledge, T.A. Supersaturation with water explains the unusual adhesion of aggregate glue in the webs of the moth-specialist spider, Cyrtarachne akirai. R. Soc. Open Sci. 2018, 5, 181296. [Google Scholar] [CrossRef]
- Diaz, C., Jr.; Baker, R.H.; Long, J.H., Jr.; Hayashi, C.Y. Connecting materials, performance and evolution: A case study of the glue of moth-catching spiders (Cyrtarachninae). J. Exp. Biol. 2022, 225 (Suppl. 1), jeb243271. [Google Scholar] [CrossRef]
- Miyashita, T.; Kasada, M.; Tanikawa, A. Experimental evidence that high humidity is an essential cue for web building in Pasilobus spiders. Behaviour 2017, 154, 709–718. [Google Scholar] [CrossRef]
- Eberhard, W.G. Aggressive chemical mimicry by a bolas spider. Science 1977, 198, 1173–1175. [Google Scholar] [CrossRef] [PubMed]
- Eberhard, W.G. The natural history and behavior of the bolas spider Mastophora dizzydeani sp. n. (Araneidae). Psyche 1980, 87, 143–169. [Google Scholar] [CrossRef]
- Yeargan, K.V. Ecology of a bolas spider, Mastophora hutchinsoni: Phenology, hunting tactics, and evidence for aggressive chemical mimicry. Oecologia 1988, 74, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Yeargan, K.V. Biology of bolas spiders. Annu. Rev. Entomol. 1994, 39, 81–99. [Google Scholar] [CrossRef]
- Gertsch, W.J. The North American bolas spiders of the genera Mastophora and Agatostichus. Bull. Am. Mus. 1955, 106, 4. [Google Scholar]
- Local weather forecast, news and Conditions. Weather Underground. (n.d.). Available online: https://www.wunderground.com/ (accessed on 19 September 2021).
- Charmant, J. Kinovea (0.8.15). Joan Charmant and Contributions. 2011. Available online: https://www.kinveo.org (accessed on 27 September 2021).
- Stauffer, S.L.; Coguill, S.L.; Lewis, R.V. Comparison of physical properties of three silks from Nephila clavipes and Araneus gemmoides. J. Arachnol. 1994, 22, 5–11. [Google Scholar]
- Harihara, P.; Childs, D.W. Solving Problems in Dynamics and Vibrations Using MATLAB; Department of Mechanical Engineering Texas A & M University: College Station, TX, USA, 2014; pp. 11–21. [Google Scholar]
- Mass-Spring-Damper in Simulink and Simscape—MATLAB & Simulink. 2021. Available online: https://www.mathworks.com/help/physmod/simscape/ug/mass-spring-damper-in-simulink-and-simscape.html (accessed on 30 September 2021).
- Blackledge, T.A.; Cardullo, R.A.; Hayashi, C.Y. Polarized light microscopy, variability in spider silk diameters, and the mechanical characterization of spider silk. Invertebr. Biol. 2005, 124, 165–173. [Google Scholar] [CrossRef]
- Agnarsson, I.; Blackledge, T.A. Can a spider web be too sticky? Tensile mechanics constrains the evolution of capture spiral stickiness in orb-weaving spiders. J. Zool. 2009, 278, 134–140. [Google Scholar] [CrossRef]
- Sahni, V.; Blackledge, T.A.; Dhinojwala, A. Changes in the adhesive properties of spider aggregate glue during the evolution of cobwebs. Sci. Rep. 2011, 1, 1. [Google Scholar] [CrossRef]
- Sensenig, A.T.; Lorentz, K.A.; Kelly, S.P.; Blackledge, T.A. Spider orb webs rely on radial threads to absorb prey kinetic energy. J. R. Soc. Interfaces 2012, 9, 1880–1891. [Google Scholar] [CrossRef] [PubMed]
- Cardé, R.T.; Mafra-Neto, A. Mechanisms of flight of male moths to pheromone. In Insect Pheromone Research; Springer: Boston, MA, USA, 1997; pp. 275–290. [Google Scholar]
- Cardé, R.T. Navigation along windborne plumes of pheromone and resource-linked odors. Annu. Rev. Entomol. 2021, 66, 317–336. [Google Scholar] [CrossRef] [PubMed]
- Wolfin, M.S.; Raguso, R.A.; Davidowitz, G.; Goyret, J. Context-dependency of in-flight responses by Manduca sexta moths to ambient differences in relative humidity. J. Exp. Biol. 2018, 221, jeb.177774. [Google Scholar]
- De Gennes, P.G.; Brochard-Wyart, F.; Quéré, D. Capillarity and Wetting Phenomena: Drops, Bubbles, Pearls, Waves; Springer: New York, NY, USA, 2004; Volume 315. [Google Scholar]
- Vuckovac, M.; Backholm, M.; Timonen, J.V.I.; Ras, R.H.A. Viscosity-enhanced droplet motion in sealed superhydrophobic capillaries. Sci. Adv. 2020, 6, eaba5197. [Google Scholar] [CrossRef] [PubMed]
- Tanikawa, A. Japanese Spiders of the Genus Ordgarius (Araneae: Araneidae). Acta Arachnol. 1997, 46, 101–110. [Google Scholar] [CrossRef][Green Version]
- Shinkai, A.; Shinkai, E. The natural history, bolas construction, and hunting behavior of the bolas spider, Ordgarius sexspinosus (Thorell) (Araneae: Araneidae). Acta Arachnol. 2002, 51, 149–154. [Google Scholar] [CrossRef]
- Haynes, K.F.; Yeargan, K.V.; Gemeno, C. Detection of prey by a spider that aggressively mimics pheromone blends. J. Insect Behav. 2001, 14, 535–544. [Google Scholar] [CrossRef]
- Hutchinson, C.E. A bolas-throwing spider. Sci. Am. 1903, 89, 172. [Google Scholar] [CrossRef][Green Version]
- Leroy, J.; Jocquac, R.; Leroy, A. On the behaviour of the African bolas-spider Cladomelea akermani Hewitt (Araneae, Araneidae, Cyrtarachninae), with description of the male. Ann. Natal Mus. 1998, 39, 1–9. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average ± Standard Deviation (N = 5) | |
| Maximum moth speed (ms−1) | 3.75 ± 3.09 |
| Maximum spider speed (ms−1) | 1.44 ± 0.98 |
| Moth Kinematics | |
| Impact velocity (ms−1) | 0.22 ± 0.17 |
| Impact kinetic energy (μJ) | 2.23 ± 2.65 |
| Maximum kinetic energy (μJ) | 710.15 ± 1165.4 |
| Capture Kinematics | |
| Reeling rate (ms−1) | 0.017 ± 0.007 |
| Falling speed (ms−1) | 0.266 ± 0.111 |
| Duration of reeling (s) | 2.25 ± 0.34 |
| Distance from spider when dropping (cm) | 1.21 ± 0.64 |
| Silk Kinematics | |
| Droplet strain (ε) | 5.95 ± 1.59 |
| Radial silk strain (ε) | 0.32 ± 0.15 |
| Droplet spring constant (μNm−1) | 10.61 ± 4.6 (N = 4) |
| Video | Maximum Acceleration amax (ms−2) | Spring Constant k (μNm−1) |
|---|---|---|
| 1 | 0.103 | 0.6 |
| 2 | 0.387 | 1.67 |
| 3 | 0.449 | 1.72 |
| 4 | 0.257 | 1.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diaz, C., Jr.; Long, J.H., Jr. Behavior and Bioadhesives: How Bolas Spiders, Mastophora hutchinsoni, Catch Moths. Insects 2022, 13, 1166. https://doi.org/10.3390/insects13121166
Diaz C Jr., Long JH Jr. Behavior and Bioadhesives: How Bolas Spiders, Mastophora hutchinsoni, Catch Moths. Insects. 2022; 13(12):1166. https://doi.org/10.3390/insects13121166
Chicago/Turabian StyleDiaz, Candido, Jr., and John H. Long, Jr. 2022. "Behavior and Bioadhesives: How Bolas Spiders, Mastophora hutchinsoni, Catch Moths" Insects 13, no. 12: 1166. https://doi.org/10.3390/insects13121166
APA StyleDiaz, C., Jr., & Long, J. H., Jr. (2022). Behavior and Bioadhesives: How Bolas Spiders, Mastophora hutchinsoni, Catch Moths. Insects, 13(12), 1166. https://doi.org/10.3390/insects13121166