


Evaluation of Phytoseiid and Iolinid Mites for Biological Control of the Tomato Russet Mite Aculops lycopersici (Acari: Eriophyidae)

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Pest, Beneficials, and Additional Food Source
2.2. Curative Releases with Nine Species of Phytoseiids on Individual Tomato Plants
2.3. Preventative Releases of Two Iolinids Species with One Application of Pollen/Week on Individual Tomato Plants
2.4. Demonstration Trial in Semi-Commercial Conditions
3. Results
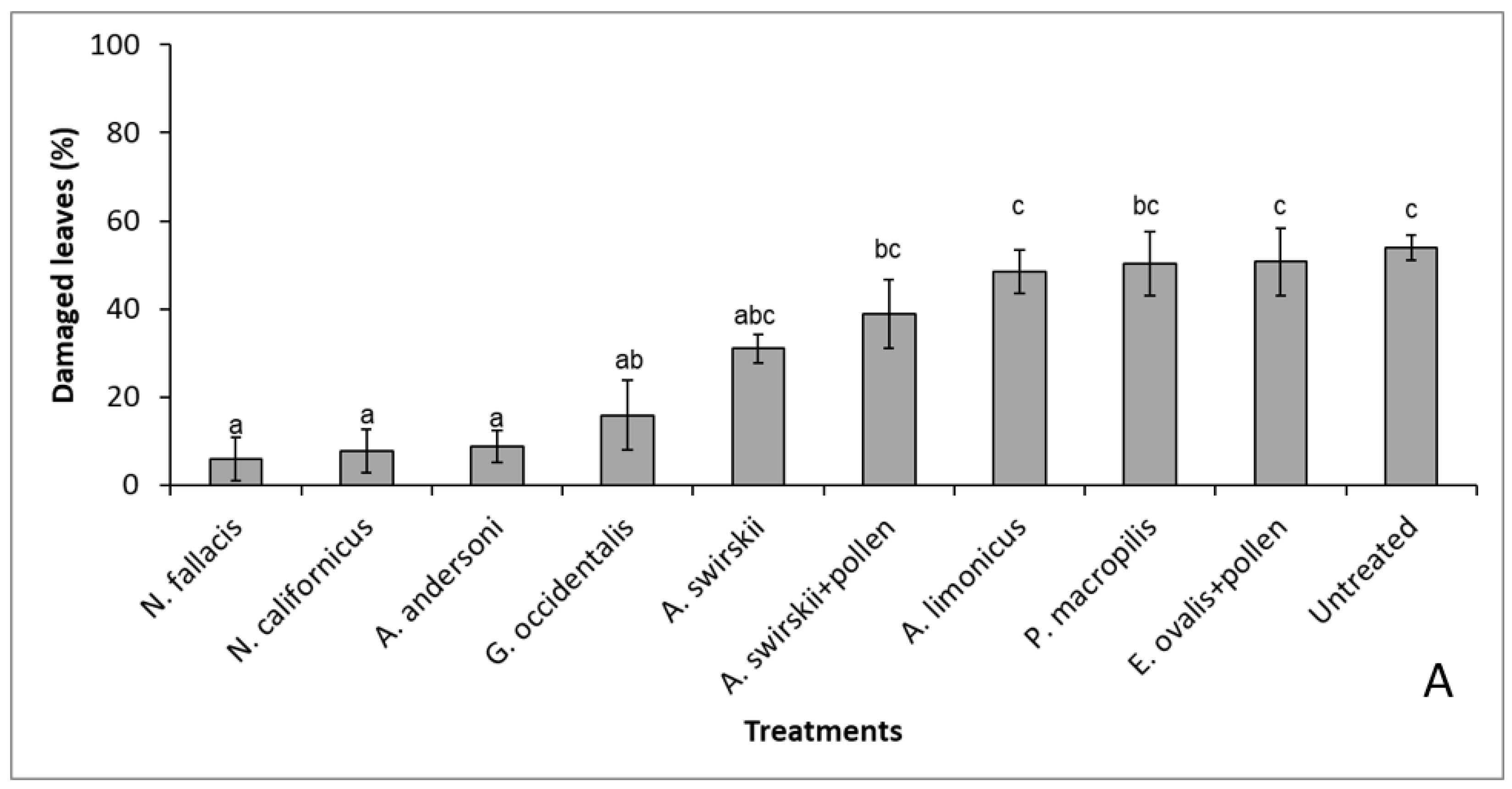
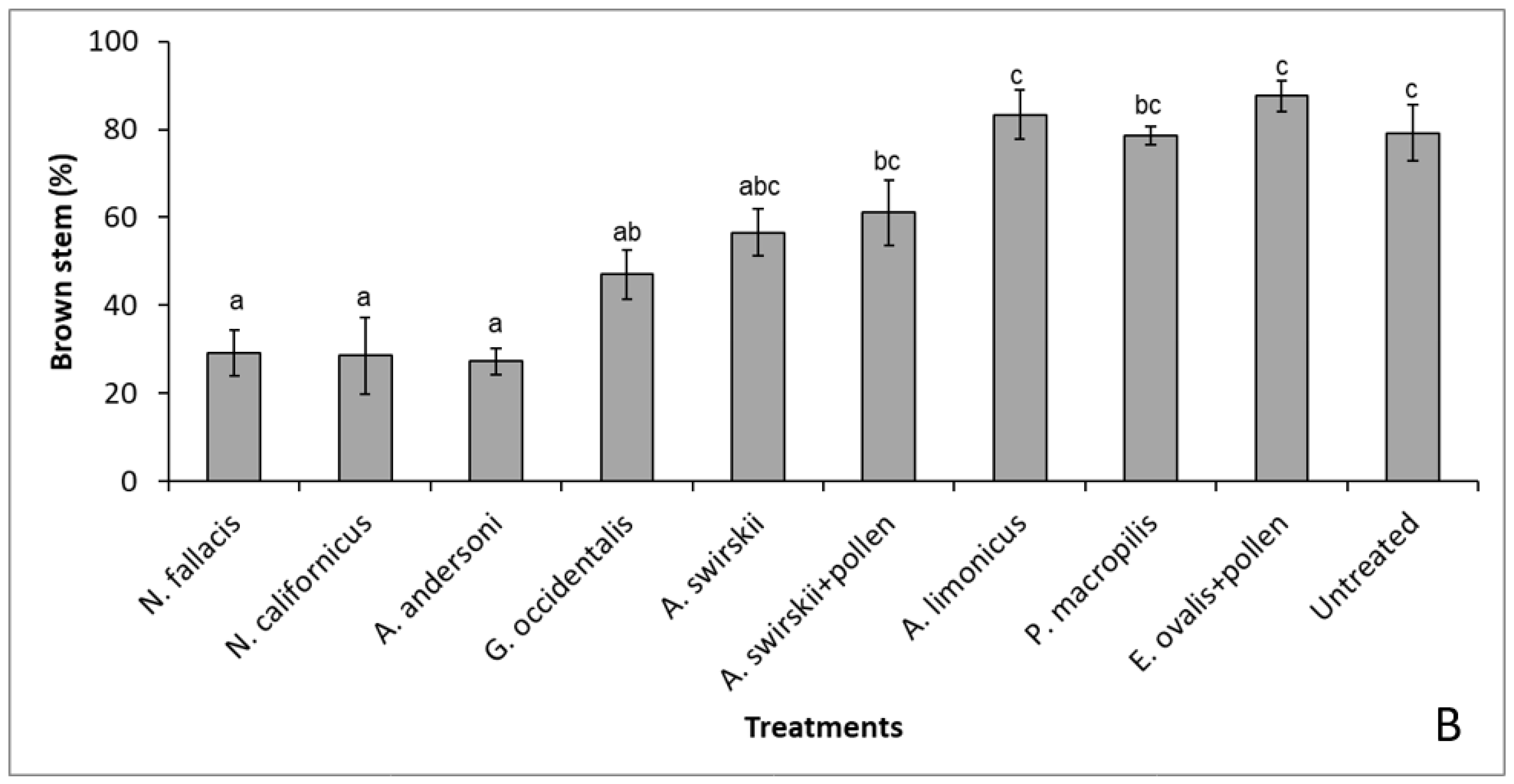
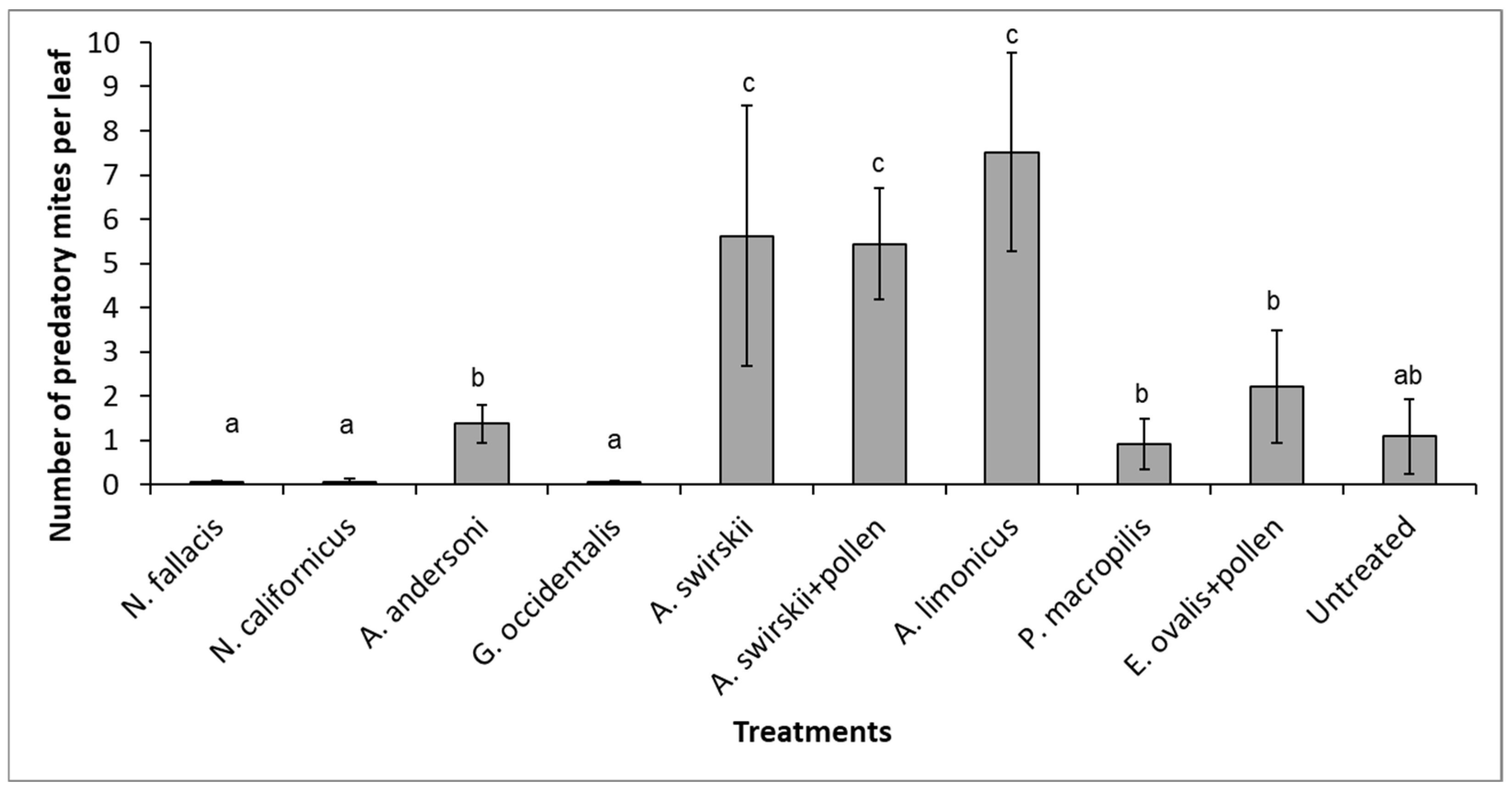
3.1. Curative Releases with Nine Species of Phytoseiids (Individual Plants)
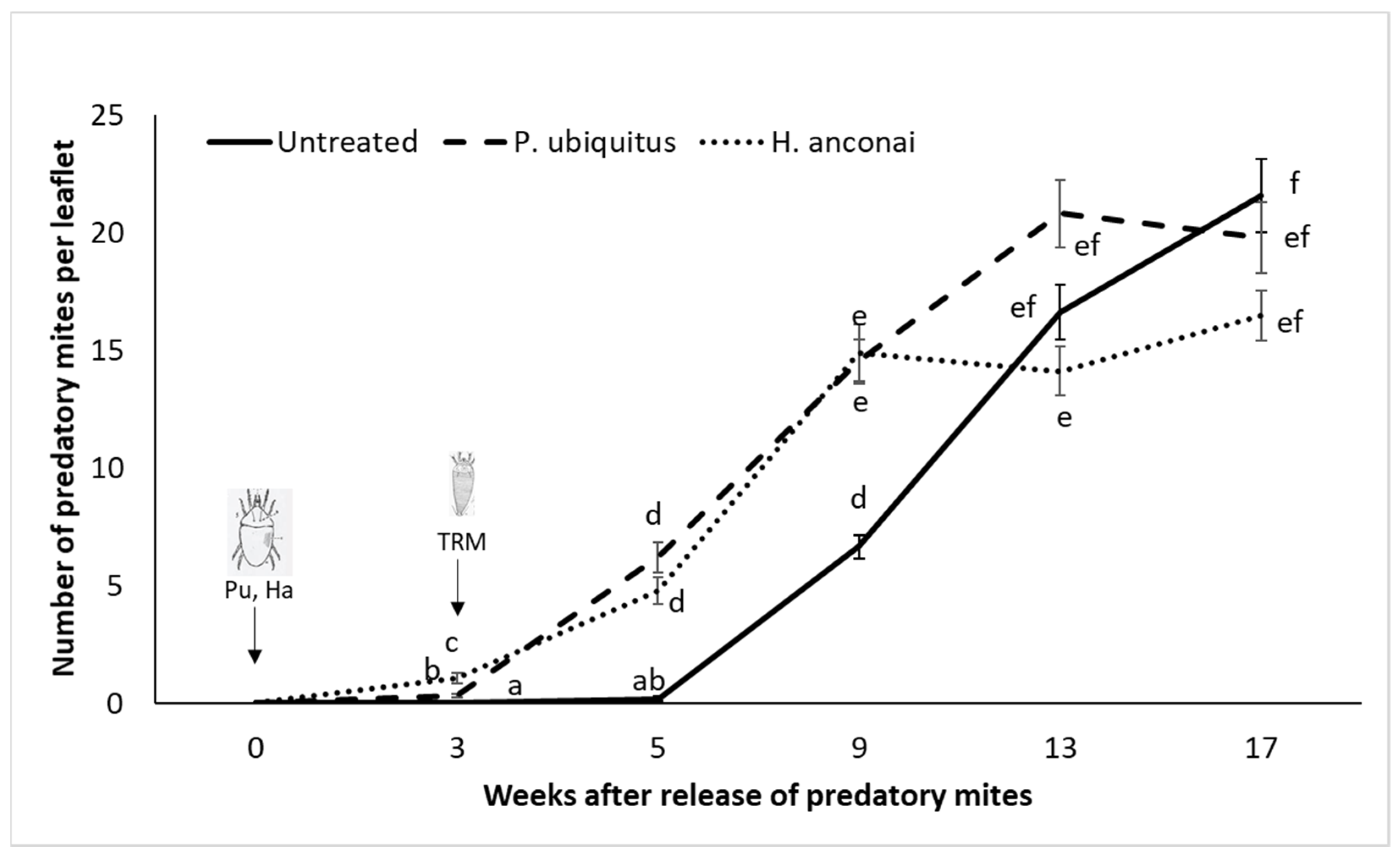
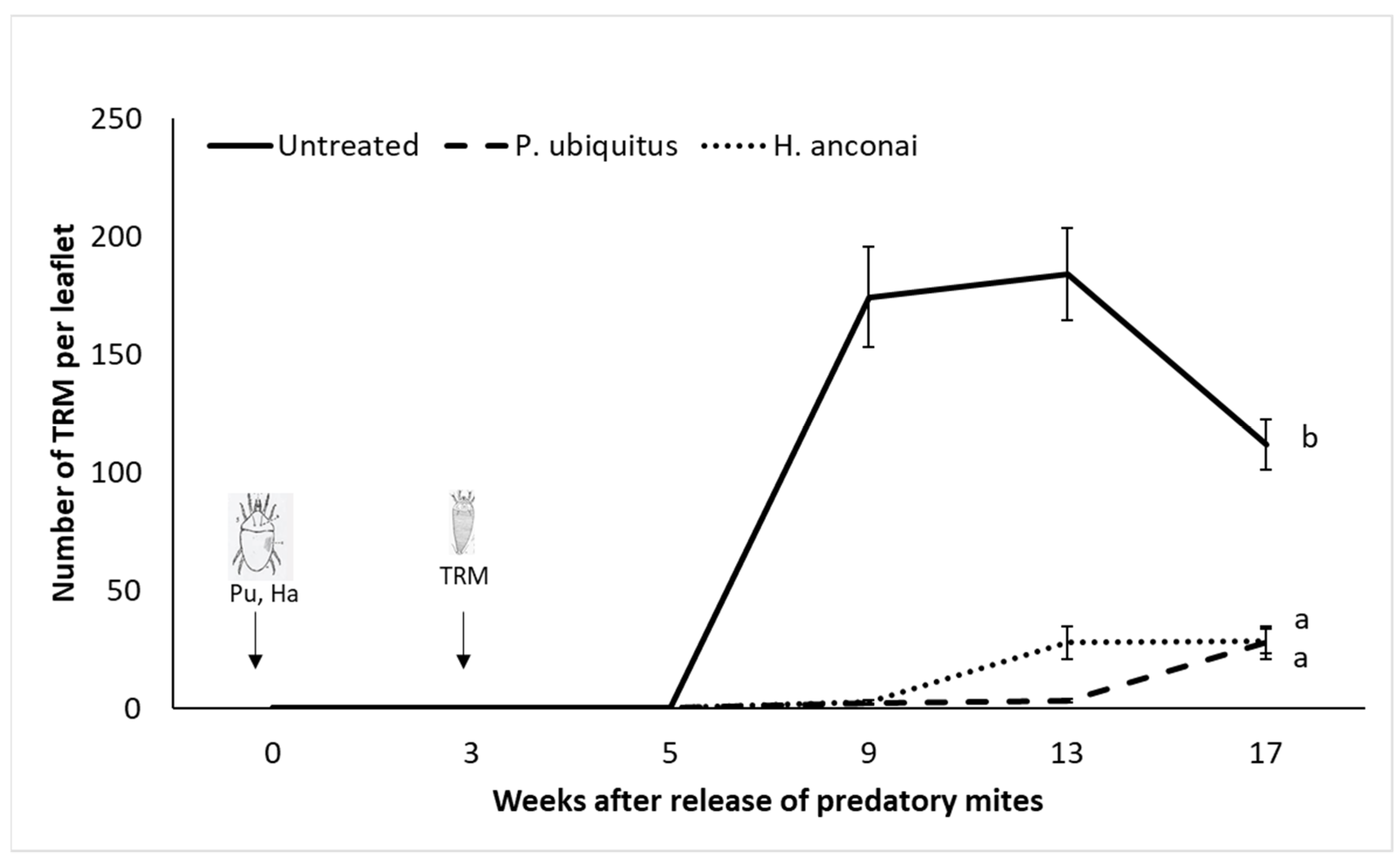
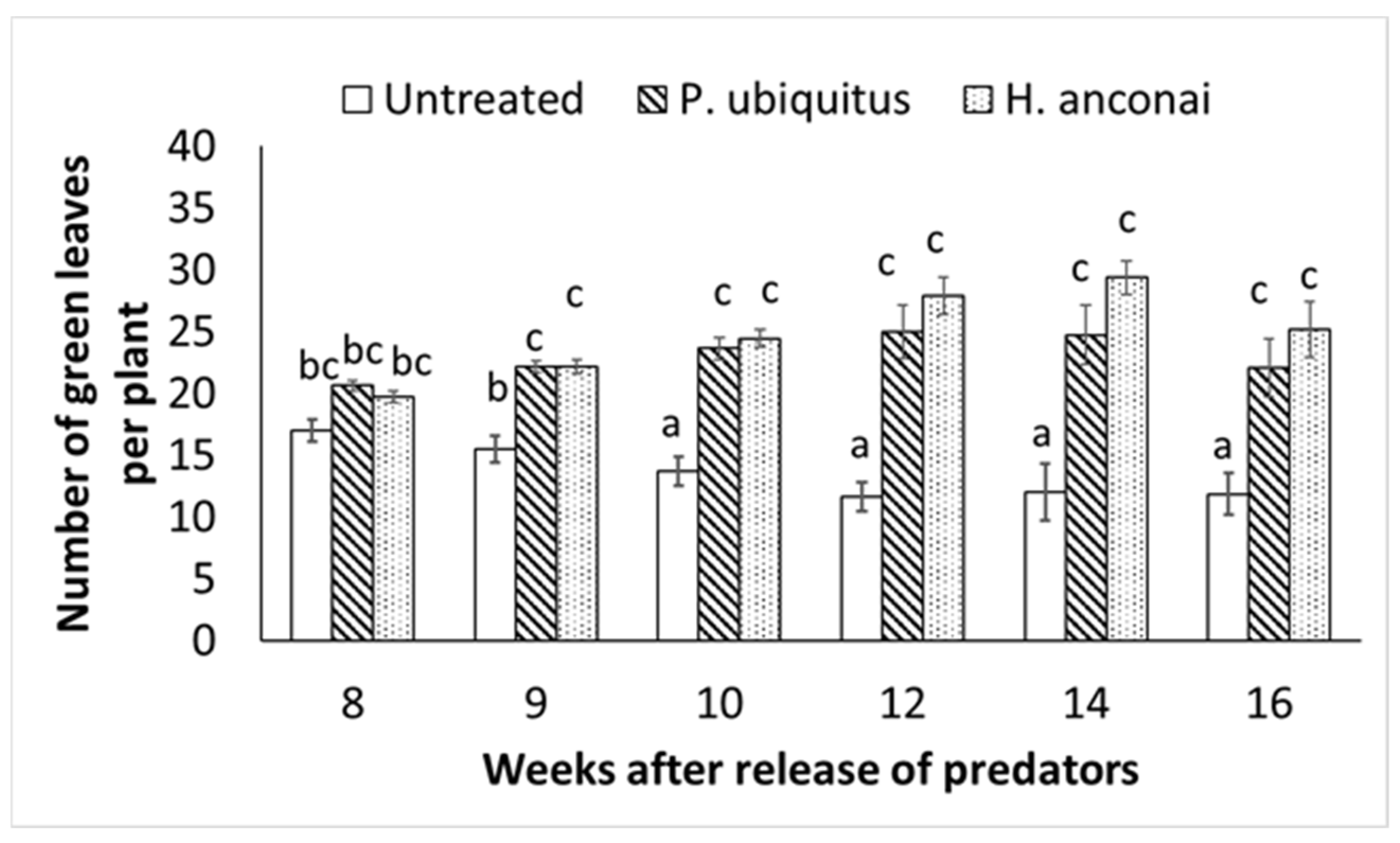
3.2. Preventative Releases of Homeopronematus Anconai and Pronematus Ubiquitus with One Application of Pollen/Week (Individual Plants)
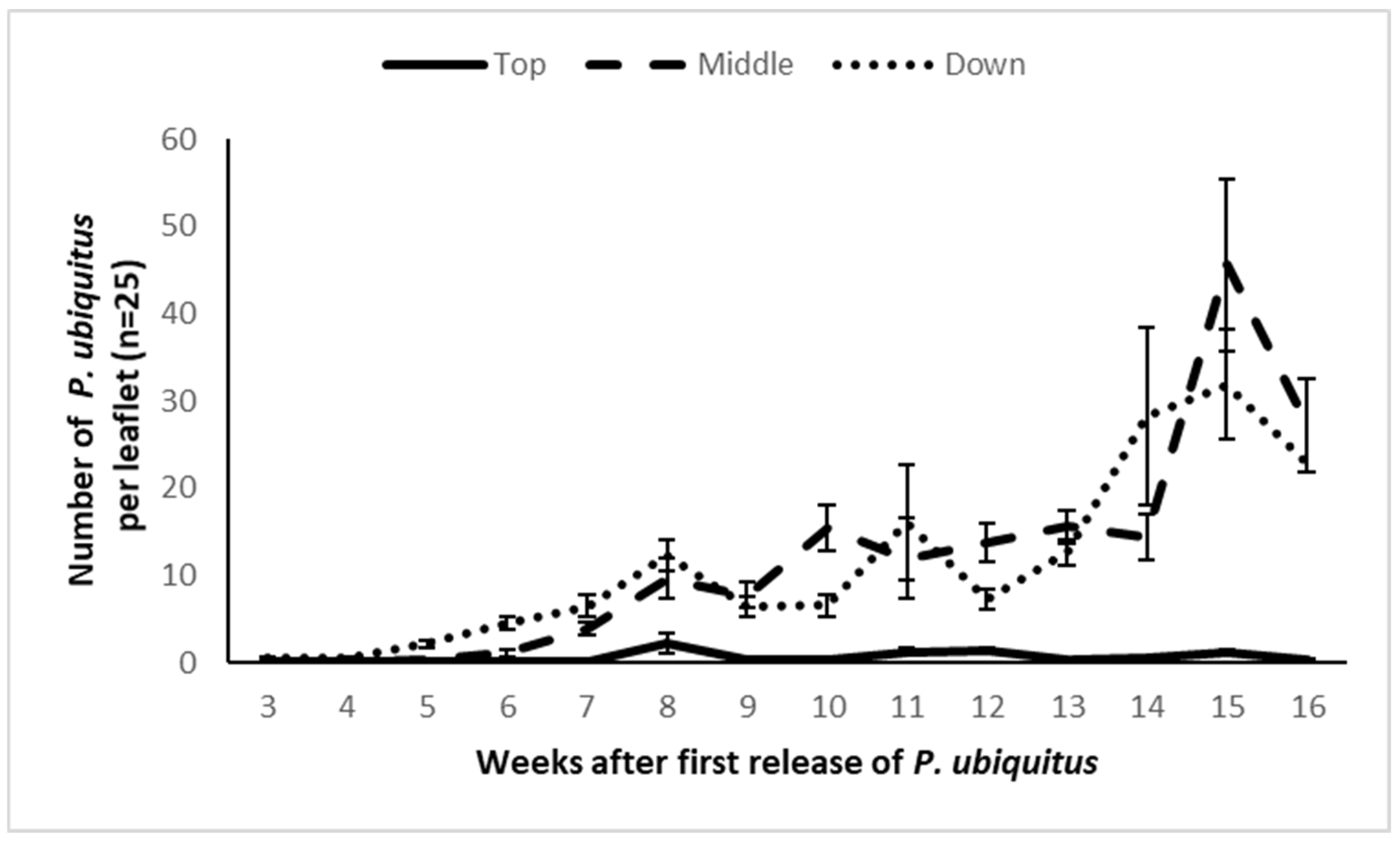
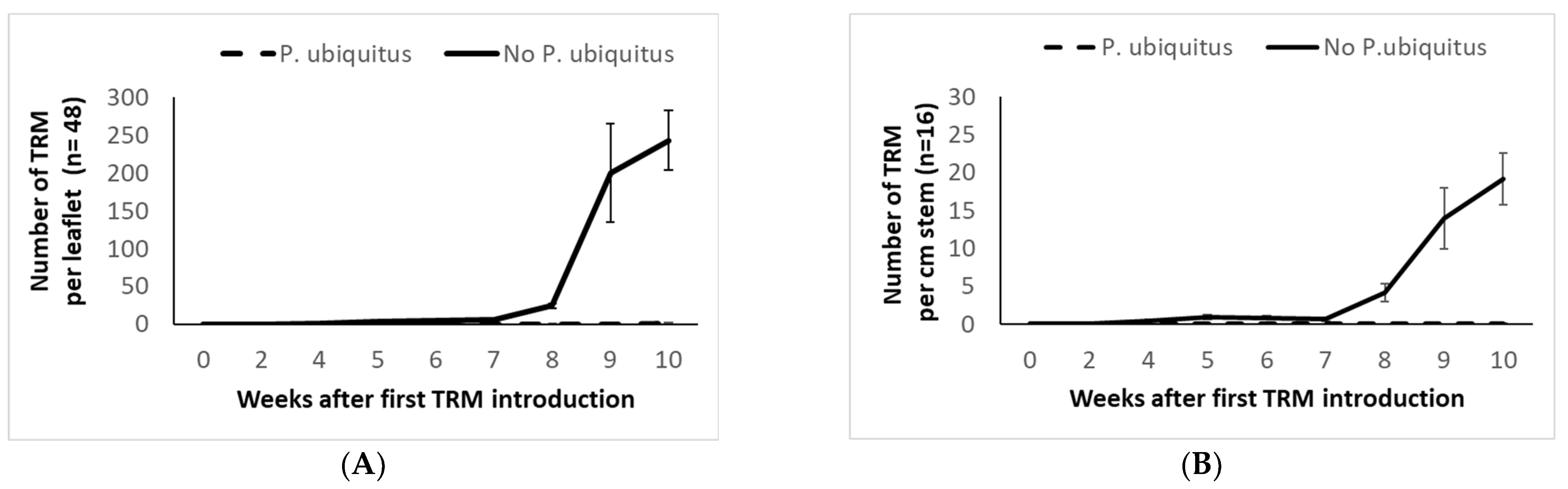
3.3. Demonstration Trial in Semi-Commercial Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vervaet, L.; De Vis, R.; De Clercq, P.; van Leeuwen, T. Is the emerging mite pest Aculops lycopersici controllable? Global and genome based insights in its biology and management. Pest Manag. Sci. 2021, 77, 2635–2644. [Google Scholar] [CrossRef] [PubMed]
- Perring, T.M.; Farrar, C.A. Historical perspective and current world status of the tomato russet mite (Acari: Eriophyidae). Misc. Publ. Entomol. Soc. Am. 1986, 63, 1–19. [Google Scholar]
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975; p. 615. [Google Scholar]
- Kay, I.R. Tomato russet mite: A serious pest of tomatoes. Qld. Agric. J. 1986, 112, 231–232. [Google Scholar]
- Keifer, H.H.; Baker, E.W.; Kono, T.; Delfinado, M.; Styler, W.E. An Illustrated Guide to Plant Abnormalities Caused by Eriophyid Mites in North America. In Agriculture Handbook; USDA: Washington, DC, USA, 1982; p. 178. [Google Scholar]
- Royalty, R.N.; Perring, T.M. Morphological analysis of damage to tomato leaflets by tomato russet mite (Acari: Eriophyidae). J. Econom. Entomol. 1988, 81, 816–820. [Google Scholar] [CrossRef]
- Duso, C.; Castagnoli, M.; Simoni, S.; Angeli, G. The impact of eriophyoids on crops: Recent issues on Aculus schlechtendali, Calepitrimerus vitis and Aculops lycopersici. Exp. Appl. Acarol. 2010, 51, 151–168. [Google Scholar] [CrossRef]
- Fischer, S.; Mourrut-Salesse, J. Tomato Russet Mite in Switzerland (Aculops lycopersici: Acari, Eriophyidae). Rev. Suisse Vitic. Arboric. Hortic. 2005, 37, 227–232. [Google Scholar]
- Steiner, M.Y.; Goodwin, S.; Wellham, T.M.; Barchia, I.M.; Spohr, L.J. Biological studies of the Australian predatory mite Typhlodromalus lailae (Schicha) (Acari: Phytoseiidae). Aust. J. Entomol. 2003, 42, 131–137. [Google Scholar] [CrossRef]
- Steiner, M.; Goodwin, S. Managing tomato russet mite, Aculops lycopersici (Massee) (Acari: Eriophyiidae) in greenhouse tomato crops. IOBC/WPRS Bull. 2005, 28, 245–248. [Google Scholar]
- Van Houten, Y.M.; Glas, J.J.; Hoogerbrugge, H.; Rothe, J.; Bolckmans, K.J.F.; Simoni, S.; Van Arkel, J.; Alba, J.M.; Kant, M.R.; Sabelis, M.W. Herbivory-associated degradation of tomato trichomes and its impact on biological control of Aculops lycopersici. Exp. Appl. Acarol. 2013, 60, 127–138. [Google Scholar] [CrossRef]
- Momen, F.M.; Abdel-Khalek, A. Effect of the tomato rust mite Aculops lycopersici (Acari: Eriophyidae) on the development and reproduction of three predatory phytoseiid mites. Int. J. Trop. Insect Sci. 2008, 28, 53–57. [Google Scholar] [CrossRef]
- Park, H.H.; Shipp, L.; Buitenhuis, R. Predation, development, and oviposition by the predatory mite Amblyseius swirskii (Acari: Phytoseiidae) on tomato russet mite (Acari: Eriophyidae). J. Econ. Entomol. 2010, 103, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Van Houten, Y.V.; Knapp, M.; Hoogerbrugge, H.; Bolckmans, K. The potential of Amblyseius swirskii as biocontrol agent for Aculops lycopersici on tomatoes. IOBC/WPRS Bull. 2013, 93, 51–57. [Google Scholar]
- Castagnoli, M.; Simoni, S.; Liguori, M. Evaluation of Neoseiulus californicus (McGregor) (Acari: Phytoseiidae) as a candidate for the control of Aculops lycopersici (Tryon) (Acari: Eriophyoidea): A preliminary study. Redia 2003, 86, 97–100. [Google Scholar]
- Brodeur, J.; Bouchard, A.; Turcott, G. Potential of four species of predatory mites as biological control agents of the tomato russet mite, Aculops lycopersici (Massee) (Eriophyidae). Can. Entomol. 1997, 129, 1–6. [Google Scholar] [CrossRef]
- Trottin-Caudal, Y.; Fournier, C.; Leyre, J.M. Biological Control of Aculops lycopersici (Massee) Using the Predatory Mites Neoseiulus californicus McGregor and Neoseiulus cucumeris (Oudemans) on Tomato Greenhouse Crops. STING 27. 2005. Available online: https://web.agrsci.dk/plb/iobc/sting/sting27 (accessed on 1 January 2021).
- Tixier, M.S.; Douin, M.; Rocio, O.; Gonzalez, L.; Pount, B.; Kreiter, S. Distribution and biological features of Typhlodromus (Anthoseius) recki (Acari: Phytoseiidae) on Tetranychus urticae, T. evansi (Acari: Tetranychidae) and Aculops lycopersici (Acari: Eriophyidae). Acarologia 2020, 60, 684–697. [Google Scholar] [CrossRef]
- Van Haren, R.J.; Steenhuis, M.M.; Sabelis, M.W.; De Ponti, O.D. Tomato stem trichomes and dispersal success of Phytoseiulus persimilis relative to its prey Tetranychus urticae. Exp. Appl. Acarol. 1987, 3, 115–121. [Google Scholar] [CrossRef]
- Kennedy, G.G. Tomato pests, parasitoids, and predators: Tritrophic interactions involving the genus Lycopersicon. Annu. Rev. Entomol. 2003, 48, 51–72. [Google Scholar] [CrossRef]
- Simmons, A.T.; Gurr, G.M. Trichomes of Lycopersicon species and their hybrids: Effects on pests and natural enemies. Agric. For. Entomol. 2005, 7, 265–276. [Google Scholar] [CrossRef]
- Sato, M.M.; de Moraes, G.J.; Haddad, M.L.; Wekesa, V.W. Effect of trichomes on the predation of Tetranychus urticae (Acari: Tetranychidae) by Phytoseiulus macropilis (Acari: Phytoseiidae) on tomato, and the interference of webbing. Exp. Appl. Acarol. 2011, 54, 21–32. [Google Scholar] [CrossRef]
- Buitenhuis, R.; Murphy, G.; Shipp, L.; Scott-Dupree, C. Amblyseius swirskii in greenhouse production systems: A floricultural perspective. Exp. Appl. Acarol. 2015, 65, 451–464. [Google Scholar] [CrossRef]
- Paspati, A.; Rambla Nebot, J.L.; López-Gresa, M.P.; Arbona, V.; Gómez-Cadenas, A.; Granell Richart, A.; González-Cabrera, J. Tomato trichomes are deadly hurdles limiting the establishment of Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae). Biol. Control 2021, 157, 1–9. [Google Scholar] [CrossRef]
- Roddick, J.G. The steroidal glycoalkaloid α-tomatine. Phytochemistry 1974, 13, 9–25. [Google Scholar] [CrossRef]
- Bingo Project (Breeding Invertebrates for Next Generation Biocontrol). Available online: https://www.bingo-itn.eu/en/bingo/show/RP3-Promoting-adaptability-of-Amblyseius-swirskii-predatory-mites-to-tomato-crop.htm (accessed on 20 July 2022).
- Legarrea, S.; Dong, L.; Glas, J.; van Houten, Y.; Kant, M.R. Distorted tomato trichomes facilitate biological control of the tomato russet mite using predatory mites. Integrated Control of Plant-Feeding Mites. IOBC/WPRS Bull. 2020, 149, 81–82. [Google Scholar]
- Legarrea, S.; Janssen, A.; Dong, L.; Glas, J.J.; van Houten, Y.M.; Scala, A.; Kant, M.R. Enhanced top-down control of herbivore population growth on plants with impaired defences. Funct. Ecol. 2022, 36, 2859–2872. [Google Scholar] [CrossRef]
- Sabelis, M.W. Phytoseiidae. In World Crop Pests 6, Eriophyoid Mites: Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier: Amsterdam, The Netherlands, 1996; pp. 427–456. [Google Scholar]
- Vervaet, L.; Parapurath, G.; De Vis, R.; van Leeuwen, T.; De Clercq, P. Potential of two omnivorous iolinid mites as predators of the tomato russet mite, Aculops lycopersici. Res. Sq. 2022, 95, 1–9. [Google Scholar] [CrossRef]
- Hessein, N.A.; Perring, T.M. Feeding habits of the Tydeidae with evidence of Homeopronematus anconai (Acari: Tydeidae) predation on Aculops lycopersici (Acari: Eriophyidae). Int. J. Acarol. 1986, 12, 215–221. [Google Scholar] [CrossRef]
- Rice, R.E. Bionomics of the Tomato Russet Mite, Aculops lycopersici. Master’s Thesis, University of California, Davis, CA, USA, 1961; p. 49. [Google Scholar]
- Carmona, M.M. Contribuicao para o conhecimento dos acaros das plantas cultivadas em Portugal V. Agron. Lusit 1970, 31, 137–183. [Google Scholar]
- Abou-Awad, B.A. The tomato russet mite, Aculops lycopersici (Massee) (Acari: Eriophyidae) in Egypt. Über die Rotgelbe Tomatenmilbe, Aculops lycopersici (Massee) (Acari, Eriophyidae) in Ägypten. Anz. Schädlingskd. Pfl. Umwelt. 1979, 52, 153–156. [Google Scholar] [CrossRef]
- Aysan, E.; Kumral, N.A. Tritrophic relationships among tomato cultivars, the rust mite, Aculops lycopersici (Massee) (Eriophyidae), and its predators. Acarologia 2018, 58, 5–17. [Google Scholar] [CrossRef]
- Kazmierski, A. A review of the genus Proctotydaeus Berlese (Actinedida: Tydeidae: Pronematinae). Acarologia 1998, 39, 33–47. [Google Scholar]
- Gerson, U.; Smiley, R.; Ochoa, R. Mites (Acari) for Pest Control 558; Blackwell Science Ltd.: Oxford, UK, 2003; p. 539. [Google Scholar]
- Flaherty, D.L.; Hoy, M.A. Biological control of Pacific mites and Willamette mites in San Joaquin Valley vineyards: Part III. Role of tydeid mites. Res. Pop. Ecol. 1971, 13, 80–96. [Google Scholar] [CrossRef]
- Calvert, D.J.; Huffaker, C.B. Predator [Metaseiulus occidentalis]-Prey [Pronematus spp.] interactions under sulfur and cattail pollen applications in a non commercial vineyard. Entomophaga 1974, 19, 361–369. [Google Scholar] [CrossRef]
- Hessein, N.A.; Perring, T.M. The importance of alternate foods for the mite Homeopronematus anconai (Acari: Tydeidae). Ann. Entomol. Soc. Am. 1988, 81, 488–492. [Google Scholar] [CrossRef]
- Da Silva, G.L.; da Cunha, U.S.; Ferla, N.J. Life cycle of Tydeus californicus (Acari: Tydeidae) on leaves of Inga marginata with and without pollen of Typha angustifolia under laboratory conditions. Int. J. Acarol. 2014, 40, 509–512. [Google Scholar] [CrossRef]
- Brickhill, C.D. Biological studies of two species of tydeid mites from California. Hilgardia 1958, 27, 601–620. [Google Scholar] [CrossRef]
- Rasmy, A.H. Relation between predaceous and phytophagous mites on Citrus. J. Appl. Entomol. 1971, 67, 6–9. [Google Scholar] [CrossRef]
- Borthakur, M. Mite predator of the scarlet mite Brevipalpus phoenicis (Geijskes). Note of the Tea Plant Protection Dept. at the Research Station, Tocklai (India). Two A Bud 1981, 18, 18–19. [Google Scholar]
- Laing, J.E.; Knop, N.F. Potential Use of Predaceous Mites Other than Phytoseiidae for Biological Control of Orchard Pests. Biological Control of Pests by Mites; University of California, Special Publication: Berkeley, CA, USA, 1983; pp. 28–35. [Google Scholar]
- Aguilar, H.; Childers, C.C.; Welbourn, W.C. Relative Abundance and Seasonal Occurrence of Mites in the Family Tydeidae on Citrus in Florida. In Proceedings of the 10th International Congress: Acarology; Halliday, R.B., Walter, D.E., Proctor, H.C., Norton, R.A., Colloff, M.J., Eds.; CSIRO Publishing: Collingwood, Australia, 2001; pp. 376–380. [Google Scholar]
- Liguori, M.; Simoni, S.; Castagnoli, M. Aspects of life history of Tydeus californicus (Banks) (Acari: Tydeidae). Redia 2002, 85, 143–153. [Google Scholar]
- Khanjani, M.; Balou, M.M. Some natural enemies of Eriophyid mites from Western Iran. IOBC/WPRS Bull. 2005, 28, 147–150. [Google Scholar]
- Lorenzon, M.; Pozzebon, A.; Duso, C.; Palevsky, E.; Weintraub, P.G.; Gerson, U.; Simoni, S. Notes on the phenology and the biology of Tydeus caudatus Dugès (Acari Tydeidae). IOBC/WPRS Bull. 2009, 50, 45–49. [Google Scholar]
- Çobanoğlu, S.; Kumral, N.A. The biodiversity and population fluctuation of plant parasitic and benificial mite species (Acari) in tomato fields of Ankara, Bursa and Yalova provinces. Turk. J. Entomol. 2014, 38, 197–214. [Google Scholar] [CrossRef]
- Schruft, G. Das Vorkommen von Milben aus der Familie Tydeidae (Acari) an Reben: VI. Beitrag über Untersuchungen zur Faunistik und Biologie der Milben (Acari) an Kulturreben (Vitis spec.). Z. Angew. Entomol. 1972, 71, 124–133. [Google Scholar] [CrossRef]
- Camporese, P.; Duso, C. Life history and life table parameters of the predatory mite Typhlodromus talbii. Entomol. Exp. Appl. 1995, 77, 149–157. [Google Scholar] [CrossRef]
- Duso, C.; Pozzebon, A.; Capuzzo, C.; Malagnini, V.; Otto, S.; Borgo, M. Grape downy mildew spread and mite seasonal abundance in vineyards: Effects on Tydeus caudatus and its predators. Biol. Control 2005, 32, 143–154. [Google Scholar] [CrossRef]
- Kawai, A.; Haque, M.M.; Tanaka, H.; Shiba, M. First record of Homeopronematus anconai (Baker) in Japan and predation on Aculops lycopersici (Massee). J. Acarol. Soc. Jpn. 2001, 10, 43–46. [Google Scholar] [CrossRef]
- Kawai, A.; Haque, M.M. Population dynamics of tomato russet mite, Aculops lycopersici (Massee) and its natural enemy, Homeopronematus anconai (Baker). Jarq-Jpn. Agr. Res. Q. 2004, 38, 161–166. [Google Scholar] [CrossRef]
- Pijnakker, J.; Arijs, Y.; de Souza, A.; Cellier, M.; Wäckers, F. The use of Typha angustifolia (cattail) pollen to establish the predatory mites Amblyseius swirskii, Iphiseius degenerans, Euseius ovalis and Euseius gallicus in glasshouse crops. Integrated Control of Plant-Feeding Mites pest. IOBC-WPRS Bull. 2016, 120, 47–54. [Google Scholar]
- Upton, M.S. Aqueous gum-chloral slide mounting media: An historical review. Bull. Entomol. Res. 1993, 83, 267–274. [Google Scholar] [CrossRef]
- Core Team, R. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Lenth, R. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]
- Van Houten, Y.M.; Hoogerbrugge, H.; Knapp, M. Potential of Pronematus ubiquitus to control tomato russet mite, Aculops lycopersici. IOBC/WPRS Bull. 2020, 149, 87–92. [Google Scholar]
- Pijnakker, J.; Hürriyet, A.; Overgaag, D.; Petit, C.; Vangansbeke, D.; Duarte, M.; Moerkens, R.; Wäckers, F. Crowd control: Well established predator populations can reduce damage by tomato russet mite Aculops lycopersici (Acari: Eriophyidae). IOBC/WPRS Bull. 2020, 149, 83–84. [Google Scholar]
- Duarte, M.V.; Vangansbeke, D.; Pijnakker, J.; Moerkens, R.; Benavente, A.; Arijs, Y.; Saucedo, A.L.F.; Wäckers, F. Evaluation of Natural and Factitious Food Sources for Pronematus ubiquitus on Tomato Plants. Insects 2021, 12, 1111. [Google Scholar] [CrossRef] [PubMed]
- Pijnakker, J.; Moerkens, R.; Vangansbeke, D.; Duarte, M.; Bellinkx, S.; Benavente, A.; Merckx, J.; Stevens, I.; Wäckers, F. Dual protection: A tydeoid mite effectively controls both a problem pest and a key pathogen in tomato. Pest Manag. Sci. 2021, 78, 355–361. [Google Scholar] [CrossRef] [PubMed]
- van de Velde, V.; Duarte, M.V.; Benavente, A.; Vangansbeke, D.; Wäckers, F.; De Clercq, P. Quest for the Allmitey: Potential of Pronematus ubiquitus (Acari: Iolinidae) as a biocontrol agent against Tetranychus urticae and Tetranychus evansi (Acari: Tetranychidae) on tomato (Solanum lycopersicum L.). bioRxiv 2021. [Google Scholar] [CrossRef]
- Knop, N.F.; Hoy, M.A. Biology of a tydeid mite, Homeopronematus anconai (n. comb.) (Acari: Tydeidae), important in San Joaquin valley vineyards. Hilgardia 1983, 51, 1–30. [Google Scholar] [CrossRef]
- van Rijn, P.C.; van Houten, Y.M.; Sabelis, M.W. Pollen improves thrips control with predatory mites. IOBC/WPRS Bull. 1999, 22, 209–212. [Google Scholar]
- Glas, J.J.; Alba, J.M.; Simoni, S.; Villarroel, C.A.; Stoops, M.; Schimmel, B.C.; Schuurink, R.C.; Sabelis, M.W.; Kant, M.R. Defense suppression benefits herbivores that have a monopoly on their feeding site but can backfire within natural communities. BMC Biol. 2014, 12, 98. [Google Scholar] [CrossRef]
- Pappas, M.L.; Broekgaarden, C.; Broufas, G.D.; Kant, M.R.; Messelink, G.J.; Steppuhn, A.; Wäckers, F.; Van Dam, N.M. Induced plant defences in biological control of arthropod pests: A double-edged sword. Pest Manag. Sci. 2017, 73, 1780–1788. [Google Scholar] [CrossRef]
- Pérez-Hedo, M.; Arias-Sanguino, Á.M.; Urbaneja, A. Induced tomato plant resistance against Tetranychus urticae triggered by the phytophagy of Nesidiocoris tenuis. Front. Plant Sci. 2018, 9, 1419. [Google Scholar] [CrossRef]
- Messelink, G.J.; Bennison, J.; Alomar, O.; Ingegno, B.L.; Tavella, L.; Shipp, L.; Palevsky, E.; Wäckers, F.L. Approaches to conserving natural enemy populations in greenhouse crops: Current methods and future prospects. BioControl 2014, 59, 377–393. [Google Scholar] [CrossRef]
- Pijnakker, J.; Vangansbeke, D.; Duarte, M.; Moerkens, R.; Wäckers, F.L. Predators and parasitoids-in-first: From inundative releases to preventative biological control in greenhouse crops. Front. Sustain. Food Syst. 2020, 4, 595630. [Google Scholar] [CrossRef]
- Benson, C.M.; Labbé, R.M. Exploring the role of supplemental foods for improved greenhouse biological control. Ann. Entomol. Soc. Am. 2021, 114, 302–321. [Google Scholar] [CrossRef]
- Royalty, R.N.; Perring, T.M. Comparative toxicity of acaricides to Aculops lycopersici and Homeopronematus anconai (Acari: Eriophyidae, Tydeidae). J. Econ. Entomol. 1987, 80, 348–351. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pijnakker, J.; Hürriyet, A.; Petit, C.; Vangansbeke, D.; Duarte, M.V.A.; Arijs, Y.; Moerkens, R.; Sutter, L.; Maret, D.; Wäckers, F. Evaluation of Phytoseiid and Iolinid Mites for Biological Control of the Tomato Russet Mite Aculops lycopersici (Acari: Eriophyidae). Insects 2022, 13, 1146. https://doi.org/10.3390/insects13121146
Pijnakker J, Hürriyet A, Petit C, Vangansbeke D, Duarte MVA, Arijs Y, Moerkens R, Sutter L, Maret D, Wäckers F. Evaluation of Phytoseiid and Iolinid Mites for Biological Control of the Tomato Russet Mite Aculops lycopersici (Acari: Eriophyidae). Insects. 2022; 13(12):1146. https://doi.org/10.3390/insects13121146
Chicago/Turabian StylePijnakker, Juliette, Asli Hürriyet, Clément Petit, Dominiek Vangansbeke, Marcus V. A. Duarte, Yves Arijs, Rob Moerkens, Louis Sutter, Dylan Maret, and Felix Wäckers. 2022. "Evaluation of Phytoseiid and Iolinid Mites for Biological Control of the Tomato Russet Mite Aculops lycopersici (Acari: Eriophyidae)" Insects 13, no. 12: 1146. https://doi.org/10.3390/insects13121146
APA StylePijnakker, J., Hürriyet, A., Petit, C., Vangansbeke, D., Duarte, M. V. A., Arijs, Y., Moerkens, R., Sutter, L., Maret, D., & Wäckers, F. (2022). Evaluation of Phytoseiid and Iolinid Mites for Biological Control of the Tomato Russet Mite Aculops lycopersici (Acari: Eriophyidae). Insects, 13(12), 1146. https://doi.org/10.3390/insects13121146