
Evaluation of the Likelihood of Establishing False Codling Moth (Thaumatotibia leucotreta) in Australia via the International Cut Flower Market


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
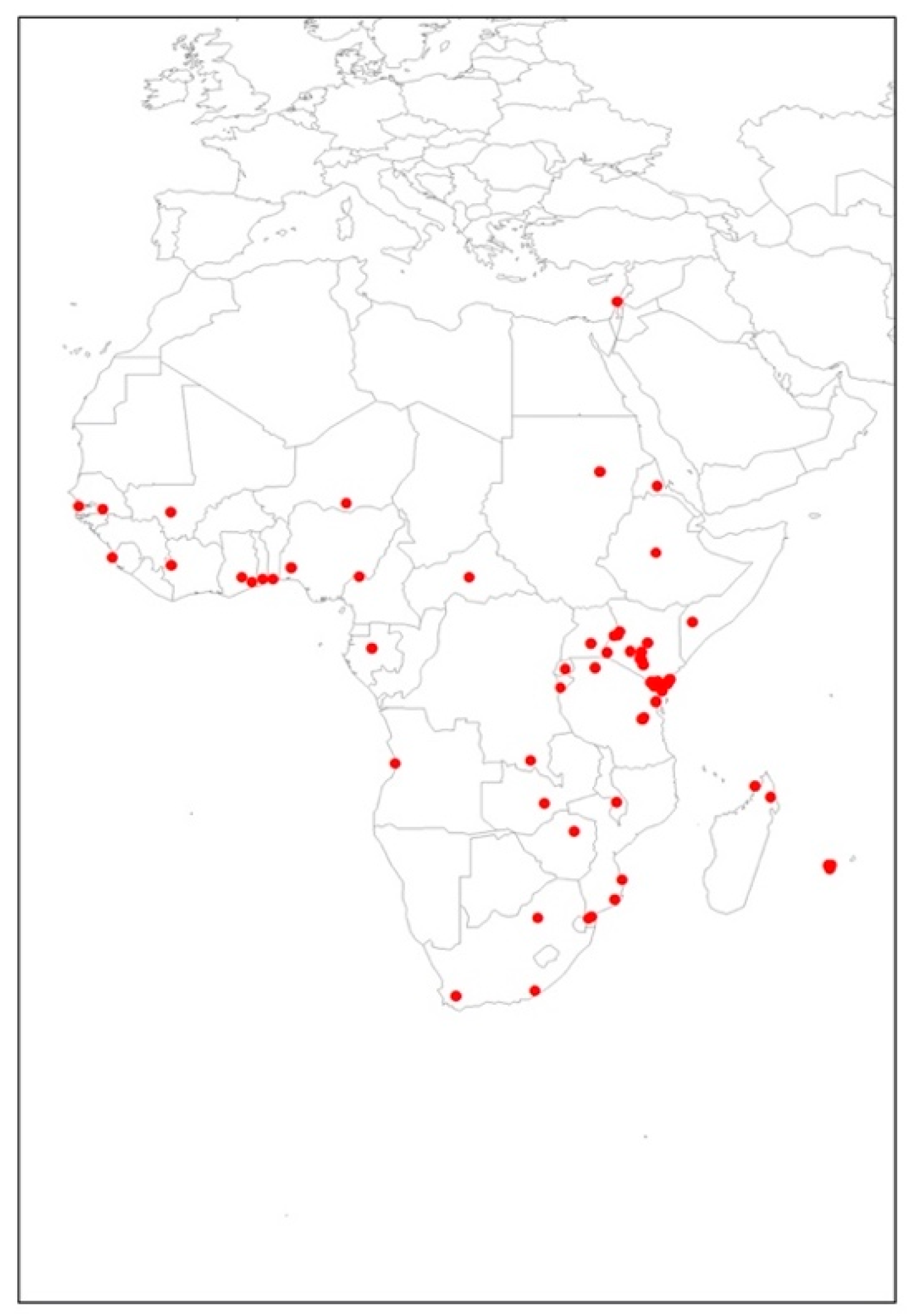
2.1. Thaumatotibia leucotreta Occurrence Data
2.2. Environmental Data
2.3. Maxent Modeling
2.4. Model Performance and Preferred Model Selection
2.5. Prediction of Habitat Suitability and Identification of Key Variables
3. Results
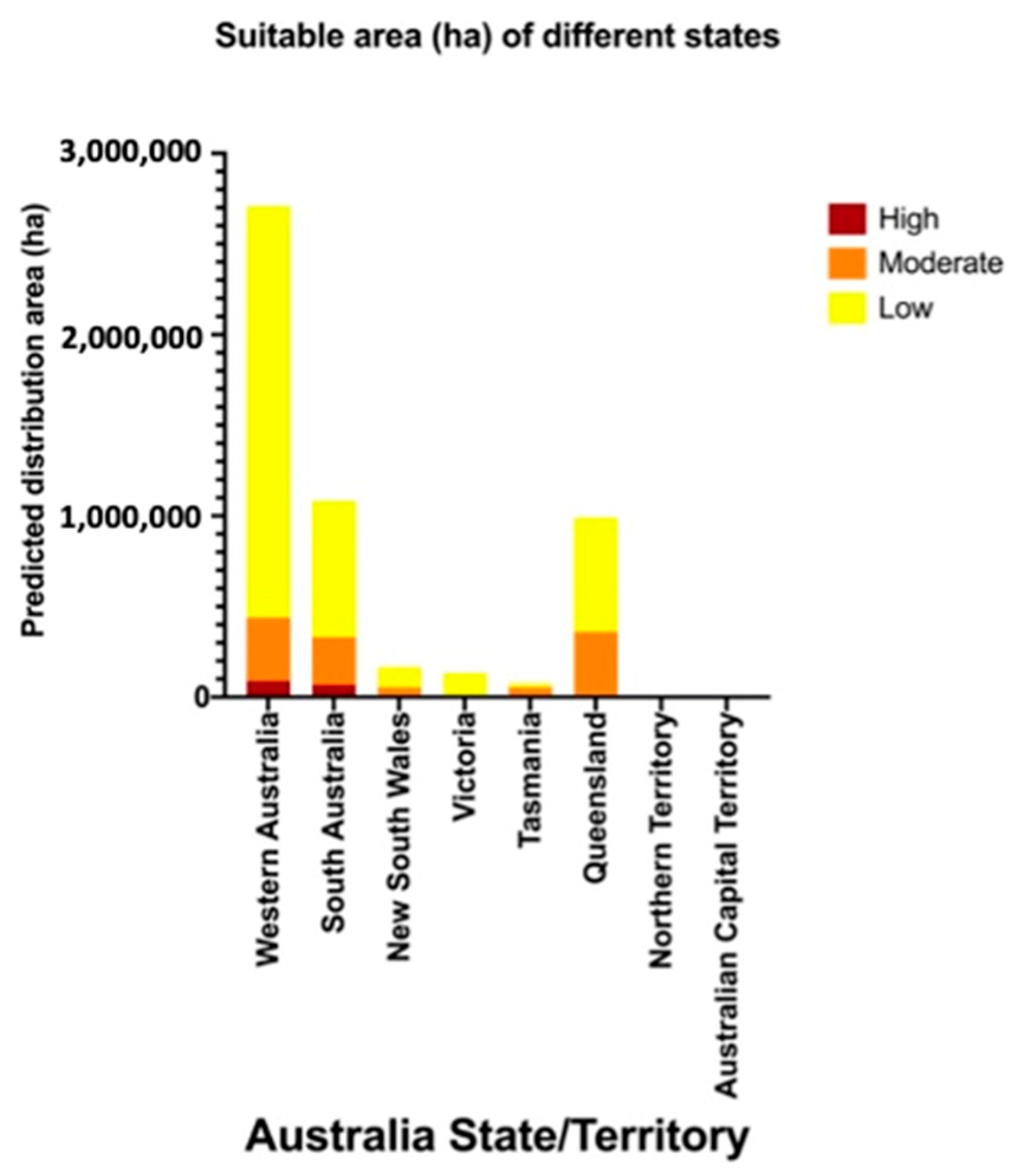
3.1. Predicted Habitat Suitability of Areas for Potential Establishment by Thaumatotibia leucotreta
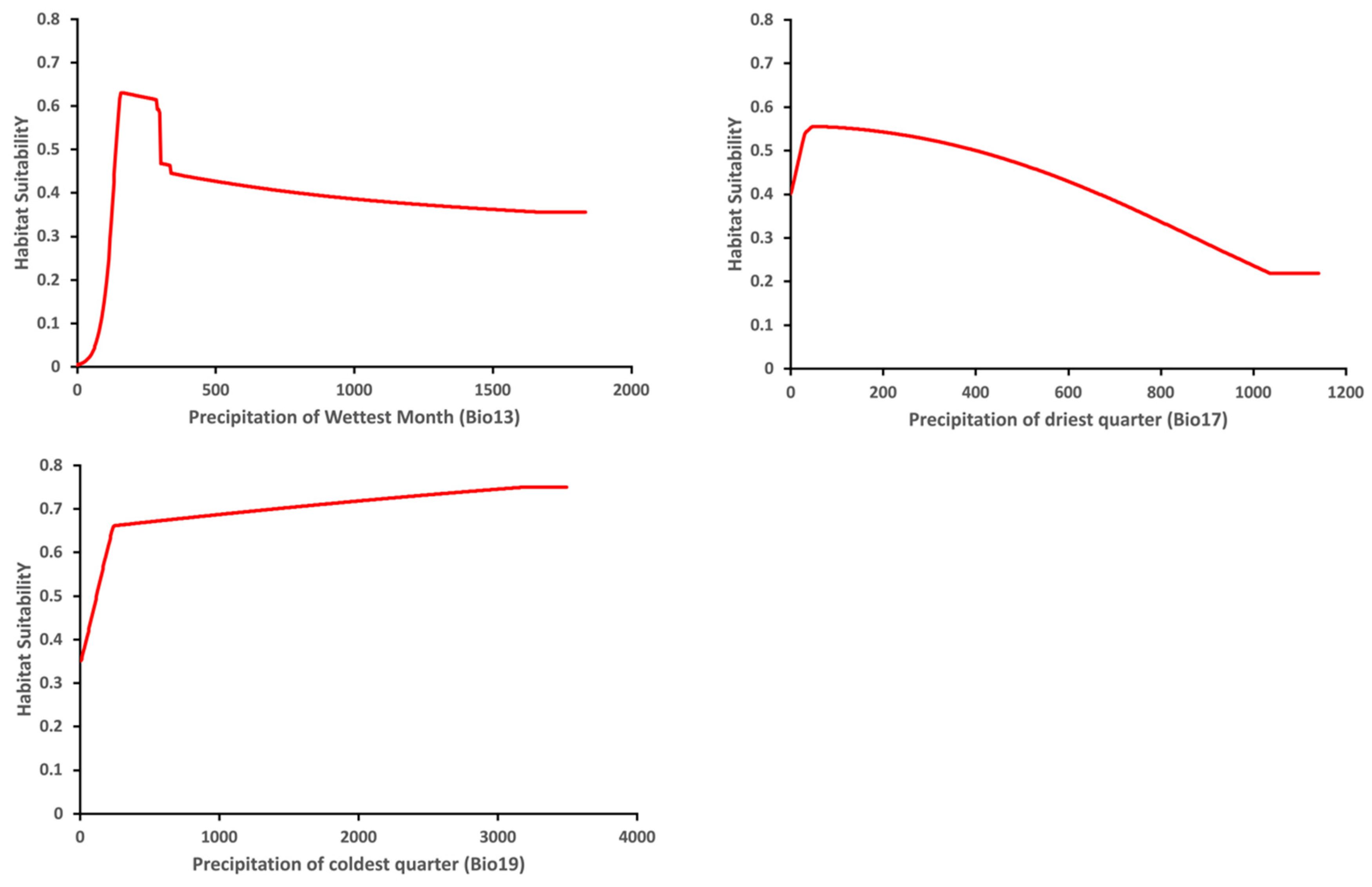
3.2. Key Climatic Variables Influencing the Predicted Distribution of Thaumatotibia leucotreta
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| bio1 | bio2 | bio3 | bio4 | bio5 | bio6 | bio7 | bio8 | bio9 | bio10 | bio11 | bio12 | bio13 | bio14 | bio15 | bio16 | bio17 | bio18 | bio19 | ND > 10 °C | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| bio1 | 1 | −0.29 | −0.2 | 0.06 | 0.89 | 0.78 | −0.27 | 0.94 | 0.92 | 0.97 | 0.94 | 0.04 | 0.15 | −0.23 | 0.34 | 0.16 | −0.14 | −0.28 | −0.03 | 0.99 |
| bio2 | −0.29 | 1 | 0.29 | 0.05 | −0.02 | −0.73 | 0.91 | −0.36 | −0.37 | −0.35 | −0.36 | −0.66 | −0.49 | −0.51 | 0.38 | −0.63 | −0.6 | −0.23 | −0.58 | −0.31 |
| bio3 | −0.2 | 0.29 | 1 | −0.87 | −0.14 | −0.06 | −0.05 | −0.38 | −0.03 | −0.35 | 0.02 | 0.05 | −0.03 | 0 | −0.06 | −0.14 | 0.03 | 0.05 | 0.11 | −0.22 |
| bio4 | 0.06 | 0.05 | −0.87 | 1 | 0.11 | −0.22 | 0.38 | 0.24 | −0.18 | 0.21 | −0.22 | −0.3 | −0.19 | −0.16 | 0.24 | −0.14 | −0.27 | −0.05 | −0.4 | 0.08 |
| bio5 | 0.89 | −0.02 | −0.14 | 0.11 | 1 | 0.55 | 0.03 | 0.81 | 0.82 | 0.89 | 0.82 | −0.18 | 0.05 | −0.45 | 0.49 | 0 | −0.36 | −0.36 | −0.18 | 0.87 |
| bio6 | 0.78 | −0.73 | −0.06 | −0.22 | 0.55 | 1 | −0.76 | 0.72 | 0.84 | 0.75 | 0.86 | 0.43 | 0.35 | 0.16 | −0.06 | 0.41 | 0.31 | −0.12 | 0.4 | 0.78 |
| bio7 | −0.27 | 0.91 | −0.05 | 0.38 | 0.03 | −0.76 | 1 | −0.28 | −0.41 | −0.26 | −0.43 | −0.73 | −0.54 | −0.54 | 0.4 | −0.63 | −0.65 | −0.24 | −0.63 | −0.28 |
| bio8 | 0.94 | −0.36 | −0.38 | 0.24 | 0.81 | 0.72 | −0.28 | 1 | 0.79 | 0.96 | 0.83 | 0.07 | 0.19 | −0.13 | 0.31 | 0.23 | −0.06 | −0.12 | −0.08 | 0.94 |
| bio9 | 0.92 | −0.37 | −0.03 | −0.18 | 0.82 | 0.84 | −0.41 | 0.79 | 1 | 0.88 | 0.97 | 0.1 | 0.19 | −0.18 | 0.23 | 0.17 | −0.03 | −0.37 | 0.19 | 0.9 |
| bio10 | 0.97 | −0.35 | −0.35 | 0.21 | 0.89 | 0.75 | −0.26 | 0.96 | 0.88 | 1 | 0.88 | 0.03 | 0.16 | −0.23 | 0.34 | 0.18 | −0.14 | −0.28 | −0.01 | 0.96 |
| bio11 | 0.94 | −0.36 | 0.02 | −0.22 | 0.82 | 0.86 | −0.43 | 0.83 | 0.97 | 0.88 | 1 | 0.16 | 0.24 | −0.17 | 0.24 | 0.23 | −0.03 | −0.28 | 0.13 | 0.93 |
| bio12 | 0.04 | −0.66 | 0.05 | −0.3 | −0.18 | 0.43 | −0.73 | 0.07 | 0.1 | 0.03 | 0.16 | 1 | 0.67 | 0.63 | −0.48 | 0.85 | 0.66 | 0.55 | 0.55 | 0.11 |
| bio13 | 0.15 | −0.49 | −0.03 | −0.19 | 0.05 | 0.35 | −0.54 | 0.19 | 0.19 | 0.16 | 0.24 | 0.67 | 1 | 0.13 | 0.13 | 0.89 | 0.18 | 0.58 | 0.15 | 0.16 |
| bio14 | −0.23 | −0.51 | 0 | −0.16 | −0.45 | 0.16 | −0.54 | −0.13 | −0.18 | −0.23 | −0.17 | 0.63 | 0.13 | 1 | −0.84 | 0.36 | 0.96 | 0.31 | 0.63 | −0.17 |
| bio15 | 0.34 | 0.38 | −0.06 | 0.24 | 0.49 | −0.06 | 0.4 | 0.31 | 0.23 | 0.34 | 0.24 | −0.48 | 0.13 | −0.84 | 1 | −0.15 | −0.85 | −0.09 | −0.71 | 0.27 |
| bio16 | 0.16 | −0.63 | −0.14 | −0.14 | 0 | 0.41 | −0.63 | 0.23 | 0.17 | 0.18 | 0.23 | 0.85 | 0.89 | 0.36 | −0.15 | 1 | 0.41 | 0.6 | 0.33 | 0.2 |
| bio17 | −0.14 | −0.6 | 0.03 | −0.27 | −0.36 | 0.31 | −0.65 | −0.06 | −0.03 | −0.14 | −0.03 | 0.66 | 0.18 | 0.96 | −0.85 | 0.41 | 1 | 0.21 | 0.75 | −0.09 |
| bio18 | −0.28 | −0.23 | 0.05 | −0.05 | −0.36 | −0.12 | −0.24 | −0.12 | −0.37 | −0.28 | −0.28 | 0.55 | 0.58 | 0.31 | −0.09 | 0.6 | 0.21 | 1 | −0.07 | −0.24 |
| bio19 | −0.03 | −0.58 | 0.11 | −0.4 | −0.18 | 0.4 | −0.63 | −0.08 | 0.19 | −0.01 | 0.13 | 0.55 | 0.15 | 0.63 | −0.71 | 0.33 | 0.75 | −0.07 | 1 | −0.01 |
| DDA10 1 | 0.99 | −0.31 | −0.22 | 0.08 | 0.87 | 0.78 | −0.28 | 0.94 | 0.9 | 0.96 | 0.93 | 0.11 | 0.16 | −0.17 | 0.27 | 0.2 | −0.09 | −0.24 | −0.01 | 1 |
References
- Australian Bureau of Agricultural and Resource Economics. Agricultural Overview: June Quarter 2021. Available online: https://www.agriculture.gov.au/abares/research-topics/agricultural-outlook/agriculture-overview (accessed on 18 February 2021).
- Australian Bureau of Agricultural and Resource Economics. Snapshot of Australian Agriculture 2021. Available online: https://www.agriculture.gov.au/abares/products/insights/snapshot-of-australian-agriculture-2021 (accessed on 10 September 2021).
- Horticulture Innovation Australia. The Australian Horticulture Statistics Handbook 2019/20. Available online: https://www.horticulture.com.au/growers/help-your-business-grow/research-reports-publications-fact-sheets-and-more/grower-resources/ha18002-assets/australian-horticulture-statistics-handbook (accessed on 15 July 2020).
- International Trade Centre. International Trade Centre Trade Map. Available online: https://www.trademap.org/Index.aspx (accessed on 3 January 2021).
- Department of Agriculture Water and the Environment. Final Risk Analysis for Cut Flower and Foliage Imports-Part 1. Available online: https://www.agriculture.gov.au/sites/default/files/sitecollectiondocuments/biosecurity/risk-analysis/plant-reviews/final-report-cut-flowers.pdf (accessed on 1 April 2020).
- European and Mediterranean Plant Protection Organization. Pest Risk Analysis for Thaumatotibia leucotreta. Available online: http://www.eppo.int/QUARANTINE/Pest_Risk_Analysis/PRA_intro.htm (accessed on 18 August 2021).
- Centre for Agriculture and Bioscience International. Thaumatotibia leucotreta. Invasive Species Compendium. Available online: https://www.cabi.org/isc/datasheet/6904 (accessed on 2 December 2020).
- Ministry of Agriculture Nature and Food Quality (Netherlands). Working Together to Stop the False Codling Moth. Available online: https://www.agroberichtenbuitenland.nl/landeninformatie/kenia/achtergrond/events/fcm-2019 (accessed on 10 November 2021).
- Loomans, A.; Noort, T.V.; Schenk, M.; Delbianco, A.; Vos, S. Pest survey card on Thaumatotibia leucotreta. EFSA Support. Publ. 2020, 17, 1916E. [Google Scholar]
- Bloem, S.; Carpenter, J.; Hofmeyr, H. Area-wide control tactics for the false codling moth Thaumatotibia leucotreta in South Africa: A potential invasive species. In Area-Wide Control of Insect Pests; Springer: Dordrecht, The Netherlands, 2007; pp. 351–359. [Google Scholar]
- Wysoki, M. New records of Lepidopterous pests of Macadamia in Israel. Phytoparasitica 1986, 14, 2. [Google Scholar] [CrossRef]
- Everatt, M. Pest Specific Plant Health Response Plan: Outbreaks for Thaumatotibia leucotreta. Available online: https://planthealthportal.defra.gov.uk/assets/uploads/Thaumatotibia-leucotreta-contingency-plan-v7.pdf (accessed on 2 August 2021).
- Mutyambai, D.M.; Mbeche, N.A.I.; Onamu, E.; Kasina, M.J.; Nderitu, J.H.; Mweke, A.N. False codling moth, Thaumatotibia leucotreta (Meyrick) a new threat to horticulture industry: Stakeholders’ perspectives on the status, impact and management in Kenya. J. Plant Dis. Prot. 2020, 127, 799–804. [Google Scholar] [CrossRef]
- Moore, S.D. Biological control of a phytosanitary pest (Thaumatotibia leucotreta): A case study. Int. J. Env. Res. Public Health 2021, 18, 1198. [Google Scholar] [CrossRef] [PubMed]
- International Plant Protection Convention. Guidelines for a Phytosanitary Import Regulatory System. Available online: https://www.ippc.int/static/media/files/publication/en/2017/08/ISPM_20_2017_En_2017-06-26.pdf (accessed on 1 September 2022).
- De-Jager, Z.M. Biology and Ecology of the False Codling Moth, Thaumatotibia leucotreta (Meyr.); Stellenbosch University: Stellenbosch, South Africa, 2013. [Google Scholar]
- Venette, R.C.; Davis, E.E.; Da-Costa, M.; Heisler, H.; Larson, M. Mini Risk Assessment: False Codling Moth, Thaumatotibia (=Cryptophlebia) leucotreta (Meyrick) [Lepidoptera: Tortricidae]; University of Minnesota: Minneapolis, MN, USA, 2003. [Google Scholar]
- Adom, M.; Fening, K.O.; Billah, M.K.; Wilson, D.D.; Hevi, W.; Clottey, V.A.; Ansah-Amprofi, F.; Bruce, A.Y. Pest status, bio-ecology and management of the false codling moth, Thaumatotibia leucotreta (Meyr.) (Lepidoptera: Tortricidae) and its implication for international trade. Bull. Entomol. Res. 2020, 111, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Timm, A.E. Morphological and Molecular Studies of Tortricid Moths of Economic Importance to the South African Fruit Industry; University of Stellenbosch: Stellenbosch, South Africa, 2005. [Google Scholar]
- Grains Research and Development Corporation. Green Bridge Factsheet. Available online: https://grdc.com.au/resources-and-publications/all-publications/factsheets/2020/grdc-fs-greenbridge (accessed on 7 July 2022).
- Stotter, R.L.; Samways, M.J.; Hattingh, V. Preparing the way for sterile insect release: Determination of false codling moth distribution across a landscape mosaic. Crop Protect. 2014, 60, 1–4. [Google Scholar] [CrossRef]
- Moraes, T.; Da-Silva, A.F.; Leite, N.A.; Karam, D.; Mendes, S.M. Survival and development of fall armyworm (Lepidoptera: Noctuidae) in weeds during the off-season. Fla. Entomol. 2020, 103, 288–292. [Google Scholar] [CrossRef]
- Plant Health Australia Ltd. Generic Contingency Plan; Exotic Chewing Insects Affecting the Grains Industry: Canberra, ACT, Australia, 2015; Available online: https://www.planthealthaustralia.com.au/wp-content/uploads/2016/03/Chewing-insect-pests-of-grain-CP.pdf (accessed on 12 June 2021).
- Australian Interim Inspector-General of Biosecurity. Effectiveness of Biosecurity Controls for Imported Fresh Cut Flowers. Available online: https://www.igb.gov.au/sites/default/files/documents/effectiveness-bio-controls-imported-fresh-cut-flowers.pdf (accessed on 5 July 2021).
- European Commission-EUROPHYT. European Union Notification System for Plant Health Interceptions Annual Report. Available online: https://food.ec.europa.eu/plants/plant-health-and-biosecurity/europhyt/interceptions-annual-reports_en (accessed on 1 June 2022).
- Reed, W. The false codling moth, Cryptophlebia leucotreta Meyr. (Lepidoptera: Olethreutidae) as a pest of cotton in Uganda. Cotton Grow. Rev. 1974, 51, 213–225. [Google Scholar]
- Terblanche, J.S.; De-Jager, Z.; Boardman, L.; Addison, P. Physiological traits suggest limited diapause response in false codling moth Thaumatotibia leucotreta (Lepidoptera: Tortricidae). J. Appl. Entomol. 2014, 138, 683–691. [Google Scholar] [CrossRef]
- Boardman, L.; Grout, T.G.; Terblanche, J.S. False codling moth Thaumatotibia leucotreta (Lepidoptera: Tortricidae) larvae are chill-susceptible. Insect Sci. 2012, 19, 315–328. [Google Scholar] [CrossRef]
- Moore, S.D.; Peyper, M.; Kirkman, W.; Marsberg, T.; Albertyn, S.; Stephen, P.R.; Thackeray, S.R.; Grout, T.G.; Sharp, G.; Sutton, G.; et al. Efficacy of various low temperature and exposure time combinations for Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae) larvae. J. Econ. Entomol. 2022, 115, 1115–1128. [Google Scholar] [CrossRef] [PubMed]
- Huisamen, E.J.; Karsten, M.; Terblanche, J.S. Consequences of Thermal Variation during Development and Transport on Flight and Low-Temperature Performance in False Codling Moth (Thaumatotibia leucotreta): Fine-Tuning Protocols for Improved Field Performance in a Sterile Insect Programme. Insects 2022, 13, 315. [Google Scholar] [CrossRef] [PubMed]
- Bettina, G.; Neil, W. Postharvest Care of Cut Flowers. Available online: https://archive.dpi.nsw.gov.au/content/agriculture/horticulture/care (accessed on 5 August 2021).
- Stotter, R.L.; Terblanche, J.S. Low-temperature tolerance of false codling moth Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae) in South Africa. J. Therm. Biol. 2009, 34, 320–325. [Google Scholar] [CrossRef]
- Stibick, J. New Pest Response Guidelines False Codling Moth Thaumatotibia leucotreta. 2007. Available online: https://docslib.org/doc/1278329/new-pest-response-guidelines-false-codling-moth-thaumatotibia-leucotreta (accessed on 11 November 2020).
- Elith, J.; Leathwick, J.R. Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Franklin, J. Mapping Species Distributions: Spatial Inference and Prediction; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Jaynes, E.T. Information theory and statistical mechanics. Phys. Rev. 1957, 106, 620. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Elith, J.H.; Graham, C.P.; Anderson, R.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend-Peterson, A. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Porter, J.H.; Parry, M.L.; Carter, T.R. The potential effects of climatic change on agricultural insect pests. Agric. For. Meteorol. 1991, 57, 221–240. [Google Scholar] [CrossRef]
- Levi-Zada, A.; Fefer, D.; Madar, R.; Steiner, S.; Kaspi, R. Evaluation of pheromone of false codling moth Thaumatotibia leucotreta in Israel by sequential SPME/GCMS analysis and field trials. J. Pest Sci. 2020, 93, 519–529. [Google Scholar] [CrossRef]
- Greene, T.; Jacobs, P. 2020 Tied for Warmest Year on Record, NASA Analysis Shows. Available online: https://www.nasa.gov/press-release/2020-tied-for-warmest-year-on-record-nasa-analysis-shows (accessed on 21 August 2021).
- Muscarella, R.; Galante, P.J.; Soley-Guardia, M.; Boria, R.A.; Kass, J.M.; Uriarte, M.; Anderson, R.P. ENMeval: An R package for conducting spatially independent evaluations and estimating optimal model complexity for Maxent ecological niche models. Methods Ecol. Evol. 2014, 5, 1198–1205. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modelling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Dudík, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Legendre, P. Spatial autocorrelation: Trouble or new paradigm? Ecology 1993, 74, 1659–1673. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef]
- Kiritani, K. Predicting impacts of global warming on population dynamics and distribution of arthropods in Japan. Popul. Ecol. 2006, 48, 5–12. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Daiber, C.C. A study of the biology of the false codling moth [Cryptophlebia leucotreta (Meyr.)]: The adult and generations during the year. Phytophylactica 1980, 12, 187–194. [Google Scholar]
- Kumar, S.; Neven, L.G.; Zhu, H.; Zhang, R. Assessing the global risk of establishment of Cydia pomonella (Lepidoptera: Tortricidae) using CLIMEX and MaxEnt niche models. J. Econ. Entomol. 2015, 108, 1708–1719. [Google Scholar] [CrossRef]
- Sambaraju, K.R.; Carroll, A.L.; Zhu, J.; Stahl, K.; Moore, R.D.; Aukema, B.H. Climate change could alter the distribution of mountain pine beetle outbreaks in western Canada. Ecography 2012, 35, 211–223. [Google Scholar] [CrossRef]
- Kumar, S.; Neven, L.G.; Yee, W.L. Evaluating correlative and mechanistic niche models for assessing the risk of pest establishment. Ecosphere 2014, 5, 1–23. [Google Scholar] [CrossRef]
- Geographic Information System Company (ESRI). ArcGIS; Environmental Systems Research Institute: Redlands, CA, USA, 2018. [Google Scholar]
- Nugent, J. Calculating Growing Degree Days. Available online: https://www.canr.msu.edu/uploads/files/Research_Center/NW_Mich_Hort/General/CalculatingGrowingDegreeDays.pdf. (accessed on 11 November 2021).
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; García Marquéz, J.R.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Phillips, S.J. A brief tutorial on Maxent. AT T Res. 2005, 190, 231–259. [Google Scholar]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Gogol-Prokurat, M. Predicting habitat suitability for rare plants at local spatial scales using a species distribution model. Ecol. Appl. 2011, 21, 33–47. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Hu, S. Akaike Information Criterion; North Carolina State University: Raleigh, NC, USA, 2012. [Google Scholar]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distribution. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Liu, C.; Newell, G.; White, M. On the selection of thresholds for predicting species occurrence with presence-only data. Ecol. Evol. 2016, 6, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed]
- Jenks, G.F. The data model concept in statistical mapping. Int. Yearb. Cartogr. 1967, 7, 186–190. [Google Scholar]
- Australian Bureau of Agricultural and Resource Economics. Catchment Scale Land Use of Australia-Update December 2020. Available online: https://www.agriculture.gov.au/abares/aclump/catchment-scale-land-use-of-australia-update-december-2020 (accessed on 18 July 2021).
- Meyer, J.S.; Ingersoll, C.G.; McDonald, L.L.; Boyce, M.S. Estimating uncertainty in population growth rates: Jackknife vs. bootstrap techniques. Ecology 1986, 67, 1156–1166. [Google Scholar] [CrossRef]
- Bulgari, R.; Petrini, A.; Cocetta, G.; Nicoletto, C.; Ertani, A.; Sambo, P.; Ferrante, A.; Nicola, S. The Impact of COVID-19 on horticulture: Critical issues and opportunities derived from an unexpected occurrence. Horticulturae 2021, 7, 124. [Google Scholar] [CrossRef]
- Inglese, P.; Sabatino, L. The fresh produce supply chain and its changes after the COVID-19. In COVID-19 and Communities; Springer: Cham, Swizherland, 2022; pp. 193–197. [Google Scholar]
- Pizano, M. Global Trends in the Cut Flower Trade. Available online: https://aiph.org/floraculture/news/global-trends-in-the-cut-flower-trade/ (accessed on 7 July 2022).
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Bureau of Meteorology. Australia. Map of Climate Zones of Australia. Available online: http://www.bom.gov.au/climate/how/newproducts/images/zones.shtml. (accessed on 27 July 2021).
- Stotter, R.L. Spatial and Temporal Distribution of False Codling Moth across Landscapes in the Citrusdal Area (Western Cape Province, South Africa); University of Stellenbosch: Stellenbosch, South Africa, 2009. [Google Scholar]
- Dawson, I. Plant Hardiness Zones for Australia. Available online: https://www.anbg.gov.au/gardens/research/hort.research/zones.html (accessed on 1 August 2021).
- Daiber, C.C. A study of the biology of the false codling moth [Cryptophlebia leucotreta (Meyr.)]: The larva. Phytophylactica 1979, 11, 123–132. [Google Scholar]
- Daiber, C.C. A study of the biology of the false codling moth [Cryptophlebia leucotreta (Meyr.)]: The cocoon. Phytophylactica 1979, 11, 151–157. [Google Scholar]
- Bureau of Meteorology. Australia. Monthly Rainfall. Available online: http://www.bom.gov.au/tmp/cdio/IDCJAC0001_009500.pdf (accessed on 12 September 2021).
- Australian Bureau of Statistics. Value of Agricultural Commodities Produces Australia 2019-20 Financial Year. Available online: https://www.abs.gov.au/statistics/industry/agriculture/value-agricultural-commodities-produced-australia/latest-release (accessed on 13 September 2021).
- Australian Bureau of Agricultural and Resource Economics. About My Region-Bunbury Wester Australia. Available online: https://www.agriculture.gov.au/abares/research-topics/aboutmyregion/wa-bunbury#forestry-sector (accessed on 15 September 2021).
- Department of Primary Industries and Regional Development. Horticulture. Available online: http://www.waopenforbusiness.wa.gov.au/Sectors/Horticulture (accessed on 15 September 2021).
- Department of Primary Industries and Regional Development. Ord River Development and Irrigated Agriculture. Available online: https://www.agric.wa.gov.au/assessment-agricultural-expansion/ord-river-development-and-irrigated-agriculture-western-australia (accessed on 16 September 2021).
- Wang, C.J.; Wang, R.; Yu, C.M.; Dang, X.P.; Sun, W.G.; Li, Q.F.; Wang, X.T.; Wan, J.Z. Risk assessment of insect pest expansion in alpine ecosystems under climate change. Pest Manag. Sci. 2021, 77, 3165–3178. [Google Scholar] [CrossRef]
- Lehmann, P.; Ammunét, T.; Barton, M.; Battisti, A.; Eigenbrode, S.D.; Jepsen, J.U.; Kalinkat, G.; Neuvonen, S.; Niemelä, P.; Terblanche, J.S.; et al. Complex responses of global insect pests to climate warming. Front. Ecol. Environ. 2020, 18, 141–150. [Google Scholar] [CrossRef]
- Willers, P.; Holmden, E.; Bedford, E.C.G. An integrated pest control programme for citrus in the Eastern Cape. Farming S. Afr. 1981, 47, 1–7. [Google Scholar]
- Moore, S.D. The Development and Evaluation of Cryptophlebia leucotreta Granulovirus (CrleGV) as a Biological Control Agent for the Management of False Codling Moth, Cryptophlebia leucotreta, on Citrus; Rhodes University: Grahamstown, South Africa, 2002. [Google Scholar]
- Carpenter, J.E.; Bloem, S.; Hofmeyr, J.H. Acceptability and suitability of eggs of false codling moth (Lepidoptera: Tortricidae) from irradiated parents to parasitism by Trichogrammatoidea cryptophlebiae (Hymenoptera: Trichogrammatidae). Biol. Control 2004, 30, 351–359. [Google Scholar] [CrossRef]
- Zimba, K.; Hill, M.P.; Moore, S.D.; Heshula, U. Agathis bishopi (Hymenoptera: Braconidae) as a potential tool for detecting oranges infested with Thaumatotibia leucotreta (Lepidoptera: Tortricidae). J. Insect Behav. 2015, 28, 618–633. [Google Scholar] [CrossRef]
- Malan, A.P.; Von-Diest, J.I.; Moore, S.D.; Addison, P. Control options for false codling moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae), in South Africa, with emphasis on the potential use of entomopathogenic nematodes and fungi. Afr. Entomol. 2018, 26, 14–29. [Google Scholar] [CrossRef]
- Newton, P.J.; Bedford, E.C.G.; Van-den-Berg, M.A.; De-Villiers, E.A. False codling moth Cryptophlebia leucotreta (Meyrick). In Citrus Pests in the Republic of South Africa. Dynamic Ad, Nelspruit, South Africa; ARC: Sheffield, UK, 1998; pp. 194–200. [Google Scholar]
- Bedford, E.C.G.; Van-den-Berg, M.A.; De-Villiers, E.A. Citrus Pests in the Republic of South Africa, 2nd ed.; Institute for Tropical and Subtropical Crops: Nelspruit, South Africa, 1998. [Google Scholar]
- Nyiira, Z.M. A note on the natural enemies of lepidopterous larvae in cotton bolls in Uganda. Ann. Entomol. Soc. Am. 1970, 63, 1461–1462. [Google Scholar] [CrossRef]
- Catung, H.D.; Born, H.A. Population studies of the false codling moth, Cryptophlebia leucotreta Meyr., on citrus in the Transvaal. Phytophylactica 1974, 6, 31–38. [Google Scholar]
- Moore, S.; Hattingh, V. A review of current pre-harvest control options for false codling moth in citrus in southern Africa. SA Fruit J. 2012, 11, 82–85. [Google Scholar]
- Fournier, D.; De-Biseau, J.C.; Aron, S. Genetics, behaviour and chemical recognition of the invading ant Pheidole megacephala. Mol. Ecol. 2009, 18, 186–199. [Google Scholar] [CrossRef]
- Jaworski, C.C.; Chailleux, A.; Bearez, P.; Desneux, N. Apparent competition between major pests reduces pest population densities on tomato crop, but not yield loss. J. Pest Sci. 2015, 88, 793–803. [Google Scholar] [CrossRef]
- European Food Safety Authority; Baker, R.; Gilioli, G.; Behring, C.; Candiani, D.; Gogin, A.; Kaluski, T.; Kinkar, M.; Mosbach-Schulz, O.; Neri, F.M.; et al. Thaumatotibia leucotreta Pest Report to Support Ranking of EU Candidate Prioirity Pests. 2019. Available online: https://docslib.org/doc/330200/thaumatotibia-leucotreta-pest-report-to-support-ranking-of-eu-candidate-priority-pests (accessed on 5 July 2021).








| Variable Code | Variable Title | Unit | Modeling Used in This Study |
|---|---|---|---|
| Bio1 | Annual mean temperature | °C | |
| Bio2 | Mean diurnal range | °C | |
| Bio3 | Isothermality | % | |
| Bio4 | Temperature seasonality | °C | |
| Bio5 | Maximum temperature of warmest month | °C | |
| Bio6 | Minimum temperature of coldest month | °C | |
| Bio7 | Temperature Annual Range | °C | |
| Bio8 | Mean temperature of wettest quarter | °C | |
| Bio9 | Mean temperature of driest quarter | °C | |
| Bio10 | Mean temperature of warmest quarter | °C | |
| Bio11 | Mean temperature of coldest quarter | °C | |
| Bio12 | Annual Precipitation | mm | |
| Bio13 | Precipitation of Wettest Month | mm | |
| Bio14 | Precipitation of Driest month | mm | |
| Bio15 | Precipitation seasonality (coefficient of variation) | % | |
| Bio16 | Precipitation of Wettest quarter | mm | |
| Bio17 | Precipitation of Driest quarter | mm | |
| Bio18 | Precipitation of Warmest Quarter | mm | |
| Bio19 | Precipitation of Coldest quarter | mm | |
| DDA10 | Degree days above 10 °C | dd |
| Suitability | High | Moderate | Low | Unsuitable | |
|---|---|---|---|---|---|
| State/Territory | |||||
| Western Australia | 89,670 | 348,652 | 2,271,156 | 41,408,183 | |
| South Australia | 69,766 | 262,205 | 750,311 | 13,348,525 | |
| New South Wales | 530 | 53,004 | 112,464 | 25,191,545 | |
| Victoria | 81 | 9564 | 123,151 | 10,449,260 | |
| Tasmania | 52 | 52,179 | 23,382 | 152,890 | |
| Queensland | 45 | 360,547 | 628,732 | 4,414,677 | |
| Northern Territory | 3 | 3 | 149 | 37,673 | |
| Australian Capital Territory | 0 | 0 | 0 | 553 | |
| Grand Total | 160,147 | 1,086,154 | 3,909,346 | 95,003,307 | |
| Regions of State/Territory | Area (ha) | Cropping | Perennial Horticulture | Seasonal Horticulture | Production from Irrigated Agriculture and Plantations | Irrigated Cropping | Irrigated Perennial Horticulture | Irrigated Seasonal Horticulture | Intensive Horticulture |
|---|---|---|---|---|---|---|---|---|---|
| Albany (WA) | 63,376 | ||||||||
| Manjimup (WA) | 22,162 | ||||||||
| Augusta-Margaret River-Busselton (WA) | 4132 | ||||||||
| Fleurieu-Kangaroo Island (SA) | 47,386 | ||||||||
| Limestone Coast (SA) | 22,328 | ||||||||
| The Eyre Peninsula and South West (SA) | 52 | ||||||||
| Shoalhaven (NSW) | 209 | ||||||||
| South Coast (NSW) | 321 | ||||||||
| Glenelg-Southern Grampians (Vic.) | 81 | ||||||||
| West Coast (Tas.) | 52 | ||||||||
| Far North (Qld) | 36 | ||||||||
| Gladstone-Biloela (Qld) | 9 | ||||||||
| Daly-Tiwi-West Arnhem (NT) | 3 |
| Variable Code | Variable Title | Unit | Percent Contribution (%) | Permutation Importance (%) |
|---|---|---|---|---|
| Bio7 | Temperature annual range | °C | 31.2 | 67.3 |
| Bio4 | Temperature seasonality | °C | 20.9 | 4.8 |
| DDA10 | Degree days above 10 °C | dd | 17.9 | 8.6 |
| Bio13 | Precipitation of wettest month | mm | 13.3 | 9.7 |
| Bio6 | Minimum temperature of coldest month | °C | 11.8 | 0.6 |
| Bio17 | Precipitation of driest quarter | mm | 4.0 | 8.0 |
| Bio19 | Precipitation of coldest quarter | mm | 0.8 | 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Emery, R.N.; Coupland, G.T.; Ren, Y.; McKirdy, S.J. Evaluation of the Likelihood of Establishing False Codling Moth (Thaumatotibia leucotreta) in Australia via the International Cut Flower Market. Insects 2022, 13, 883. https://doi.org/10.3390/insects13100883
Li X, Emery RN, Coupland GT, Ren Y, McKirdy SJ. Evaluation of the Likelihood of Establishing False Codling Moth (Thaumatotibia leucotreta) in Australia via the International Cut Flower Market. Insects. 2022; 13(10):883. https://doi.org/10.3390/insects13100883
Chicago/Turabian StyleLi, Xingyu, Robert N. Emery, Grey T. Coupland, Yonglin Ren, and Simon J. McKirdy. 2022. "Evaluation of the Likelihood of Establishing False Codling Moth (Thaumatotibia leucotreta) in Australia via the International Cut Flower Market" Insects 13, no. 10: 883. https://doi.org/10.3390/insects13100883
APA StyleLi, X., Emery, R. N., Coupland, G. T., Ren, Y., & McKirdy, S. J. (2022). Evaluation of the Likelihood of Establishing False Codling Moth (Thaumatotibia leucotreta) in Australia via the International Cut Flower Market. Insects, 13(10), 883. https://doi.org/10.3390/insects13100883