Trypanosoma cruzi Parasite Load Modulates the Circadian Activity Pattern of Triatoma infestans

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Circadian Locomotor Activity
2.2. Detection and Quantification Trypanosoma cruzi DNA
2.3. Statistical Analyses
3. Results
3.1. Triatomine Circadian Locomotor Activity
3.2. Detection and Quantification of Trypanosoma cruzi DNA
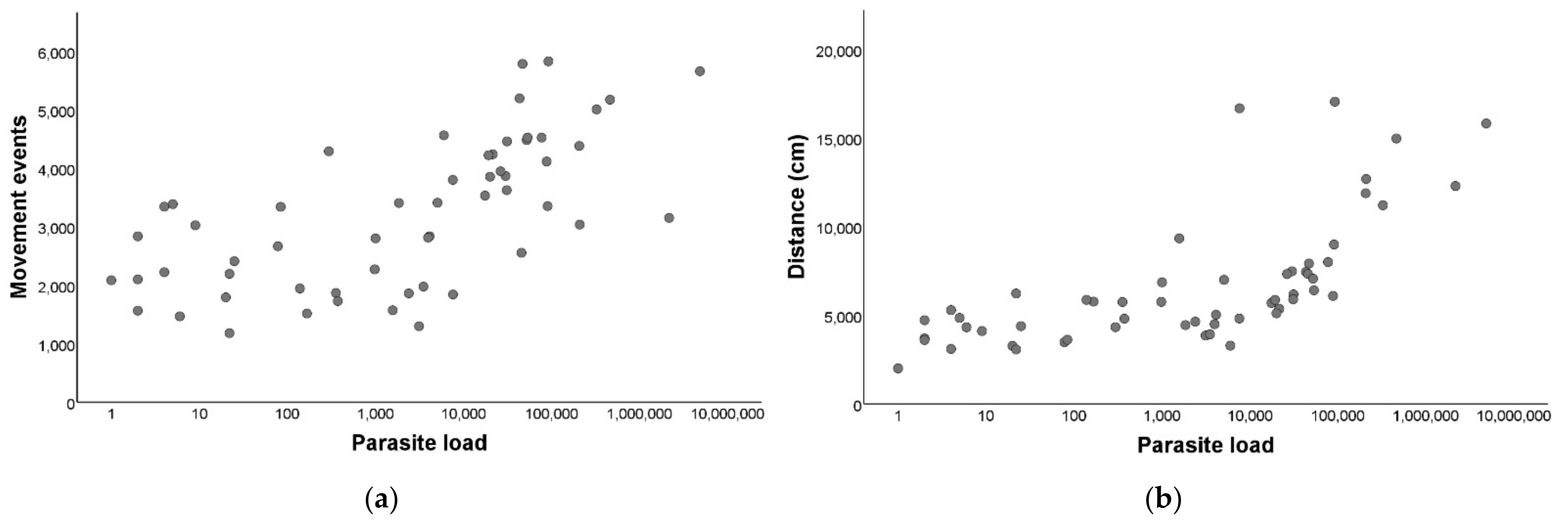
3.3. Triatomine Circadian Locomotor Activity and Parasite Load
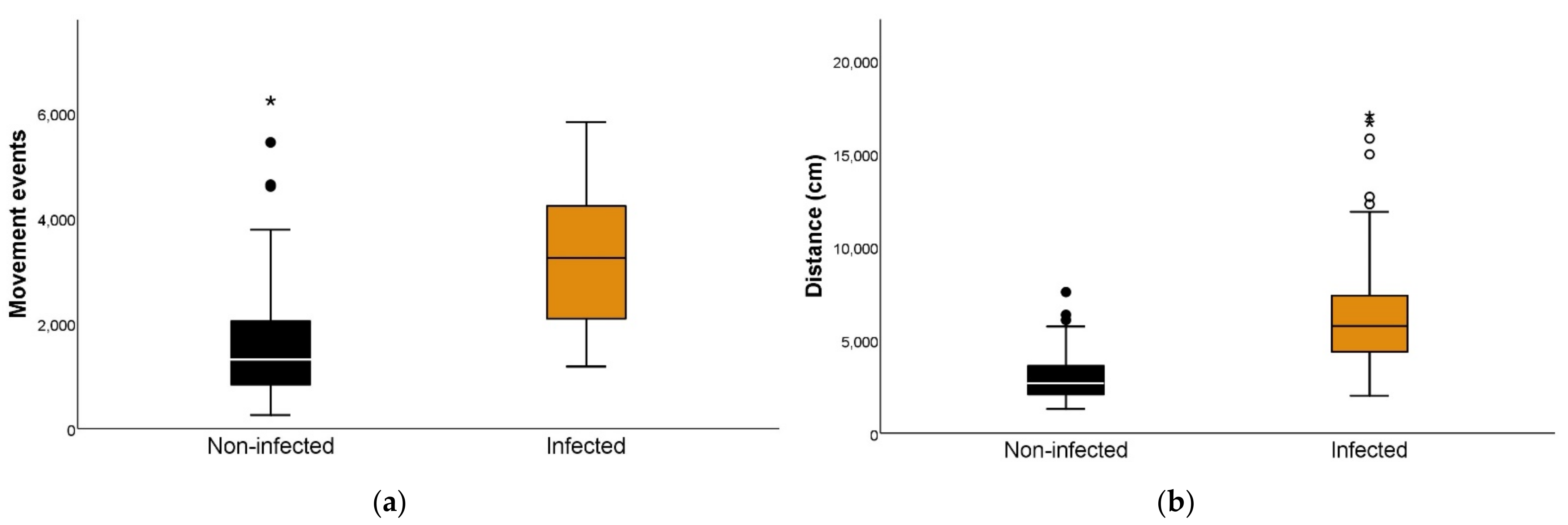
3.4. Parasite Load Effect on the Movement Events of Triatoma infestans
3.5. Parasite Load Effect on the Distance Traveled by Triatoma infestans
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Chagas Disease (Also Known as American Trypanosomiasis). Available online: https://www.who.int/es/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 8 May 2021).
- Lent, H.; Wygodzinsky, P.W. Revision of the Triatominae (Hemiptera, Reduviidae), and their significance as vectors of Chagas’ disease. Bull. Am. Mus. Nat. Hist. 1979, 163, 130–138. [Google Scholar]
- Noireau, F.; Bastrenta, B.; Catala, S.; Dujardin, J.P.; Panzera, F.; Torres, M.; Perez, R.; Galvao, C.; Jurberg, J. Sylvatic population of Triatoma infestans from the Bolivian Chaco: From field collection to characterization. Mem. Inst. Oswaldo Cruz 2000, 95 (Suppl. 1), 119–122. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Noireau, F.; Diosque, P.; Jansen, A.M. Trypanosoma cruzi: Adaptation to its vectors and its hosts. Vet. Res. 2009, 40. [Google Scholar] [CrossRef] [PubMed]
- Bacigalupo, A.; Segura, J.A.; García, A.; Hidalgo, J.; Galuppo, S.; Cattan, P.E. Primer hallazgo de vectores de la enfermedad de Chagas asociados a matorrales silvestres en la Región Metropolitana, Chile. Rev. Med. Chil. 2006, 134, 1230–1236. [Google Scholar] [CrossRef]
- Bacigalupo, A.; Torres-Pérez, F.; Segovia, V.; García, A.; Correa, J.P.; Moreno, L.; Arroyo, P.; Cattan, P.E. Sylvatic foci of the Chagas disease vector Triatoma infestans in Chile: Description of a new focus and challenges for control programs. Mem. Inst. Oswaldo Cruz 2010, 105, 633–641. [Google Scholar] [CrossRef]
- Noireau, F.; Cortez, M.G.; Monteiro, F.A.; Jansen, A.M.; Torrico, F. Can wild Triatoma infestans foci in Bolivia jeopardize Chagas disease control efforts? Trends Parasitol. 2005, 21, 7–10. [Google Scholar] [CrossRef]
- Ceballos, L.A.; Piccinali, R.V.; Berkunsky, I.; Kitron, U.; Gürtler, R.E. First finding of melanic sylvatic Triatoma infestans (Hemiptera: Reduviidae) colonies in the Argentine Chaco. J. Med. Entomol. 2009, 46, 1195–1202. [Google Scholar] [CrossRef]
- Rolón, M.; Vega, M.C.; Román, F.; Gómez, A.; Rojas de Arias, A. First report of colonies of sylvatic Triatoma infestans (Hemiptera: Reduviidae) in the Paraguayan Chaco, using a trained dog. PLoS Negl. Trop. Dis. 2011, 5, e1026. [Google Scholar] [CrossRef]
- Poulin, R.; Thomas, F. Phenotypic variability induced by parasites: Extent and evolutionary implications. Parasitol. Today 1999, 15, 28–32. [Google Scholar] [CrossRef]
- Botto-Mahan, C.; Cattan, P.E.; Medel, R. Chagas disease parasite induces behavioural changes in the kissing bug Mepraia spinolai. Acta Trop. 2006, 98, 219–223. [Google Scholar] [CrossRef]
- Marliére, N.P.; Latorre-Estivalis, J.M.; Lorenzo, M.G.; Carrasco, D.; Alves-Silva, J.; Rodrigues, J.e.O.; Ferreira, L.e.L.; Lara, L.e.M.; Lowenberger, C.; Guarneri, A.A. Trypanosomes modify the behavior of their insect hosts: Effects on locomotion and on the expression of a related gene. PLoS Negl. Trop. Dis. 2015, 9, e0003973. [Google Scholar] [CrossRef]
- Ramírez-González, M.G.; Flores-Villegas, A.L.; Salazar-Schettino, P.M.; Gutiérrez-Cabrera, A.E.; Rojas-Ortega, E.; Córdoba-Aguilar, A. Zombie bugs? Manipulation of kissing bug behavior by the parasite Trypanosoma cruzi. Acta Trop. 2019, 200, 105177. [Google Scholar] [CrossRef]
- Pérez, G.; Muñoz-San Martín, C.; Chacón, F.; Bacigalupo, A.; Cattan, P.E.; Solís, R. Modification of the daily activity pattern of the diurnal triatomine Mepraia spinolai (Hemiptera: Reduviidae) induced by Trypanosoma cruzi (Trypanosomatida: Trypanosomatidae) infection. J. Med. Entomol. 2021, 58, 2474–2478. [Google Scholar] [CrossRef]
- Pereyra, N.; Lobbia, P.A.; Mougabure-Cueto, G. Effects of the infection with Trypanosoma cruzi on the feeding and excretion/defecation patterns of Triatoma infestans. Bull. Entomol. Res. 2020, 110, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Goddard, J. Infectious Diseases and Arthropods, 3rd ed.; Humana Press: Cham, Switzerland, 2018; pp. 33–34. [Google Scholar] [CrossRef]
- Barrozo, R.B.; Reisenman, C.E.; Guerenstein, P.; Lazzari, C.R.; Lorenzo, M.G. An inside look at the sensory biology of triatomines. J. Insect Physiol. 2017, 97, 3–19. [Google Scholar] [CrossRef]
- Lorenzo, M.G.; Lazzari, C.R. Activity pattern in relation to refuge exploitation and feeding in Triatoma infestans (Hemiptera: Reduviidae). Acta Trop. 1998, 70, 163–170. [Google Scholar] [CrossRef]
- Núñez, J.A. Food source orientation and activity in Rhodnius prolixus Stål (Hemiptera: Reduviidae). Bull. Entomol. Res. 1982, 72, 253–262. [Google Scholar] [CrossRef]
- McEwen, P.; Lehane, M. Factors influencing flight initiation in the triatomine bug Triatoma sordida (Hemiptera: Reduviidae). Int. J. Trop. Insect Sci. 1993, 14, 461–464. [Google Scholar] [CrossRef]
- Barrozo, R.B.; Minoli, S.A.; Lazzari, C.R. Circadian rhythm of behavioural responsiveness to carbon dioxide in the blood-sucking bug Triatoma infestans (Heteroptera: Reduviidae). J. Insect Physiol. 2004, 50, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Bodin, A.; Barrozo, R.B.; Couton, L.; Lazzari, C.R. Temporal modulation and adaptive control of the behavioural response to odours in Rhodnius prolixus. J. Insect Physiol. 2008, 54, 1343–1348. [Google Scholar] [CrossRef]
- Pavan, M.G.; Corrêa-Antônio, J.; Peixoto, A.A.; Monteiro, F.A.; Rivas, G.B.S. Rhodnius prolixus and R. robustus (Hemiptera: Reduviidae) nymphs show different locomotor patterns on an automated recording system. Parasites Vectors 2016, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.; Garay, M.; Gorla, D.; Murua, F.; Favot, R. Caracterización de los estadios ninfales del género Triatoma Laporte 1833 I. Triatoma infestans Klug, 1834 (Hemiptera, Reduviidae). Rev. Soc. Entomol. Argent. 1981, 40, 91–102. [Google Scholar]
- Ott Joslin, J. Blood collection techniques in exotic small mammals. J. Exot. Pet. Med. 2009, 18, 117–139. [Google Scholar] [CrossRef]
- Erickson, R.L.; Terzi, M.C.; Jaber, S.M.; Hankenson, F.C.; McKinstry-Wu, A.; Kelz, M.B.; Marx, J.O. Intraperitoneal continuous-Rate infusion for the maintenance of anesthesia in laboratory mice (Mus musculus). J. Am. Assoc. Lab. Anim. Sci. 2016, 55, 548–557. [Google Scholar]
- Reisenman, C.E.; Lazzari, C. Spectral sensitivity of the photonegative reaction of the blood-sucking bug Triatoma infestans (Heteroptera: Reduviidae). J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2006, 192, 39–44. [Google Scholar] [CrossRef]
- Piron, M.; Fisa, R.; Casamitjana, N.; López-Chejade, P.; Puig, L.; Vergés, M.; Gascón, J.; Gómez i Prat, J.; Portús, M.; Sauleda, S. Development of a real-time PCR assay for Trypanosoma cruzi detection in blood samples. Acta Trop. 2007, 103, 195–200. [Google Scholar] [CrossRef]
- Duffy, T.; Bisio, M.; Altcheh, J.; Burgos, J.M.; Diez, M.; Levin, M.J.; Favaloro, R.R.; Freilij, H.; Schijman, A.G. Accurate real-time PCR strategy for monitoring bloodstream parasitic loads in Chagas disease patients. PLoS Negl. Trop. Dis. 2009, 3, e419. [Google Scholar] [CrossRef]
- Moreira, O.C.; Ramírez, J.D.; Velázquez, E.; Melo, M.F.; Lima-Ferreira, C.; Guhl, F.; Sosa-Estani, S.; Marin-Neto, J.A.; Morillo, C.A.; Britto, C. Towards the establishment of a consensus real-time qPCR to monitor Trypanosoma cruzi parasitemia in patients with chronic Chagas disease cardiomyopathy: A substudy from the BENEFIT trial. Acta Trop. 2013, 125, 23–31. [Google Scholar] [CrossRef]
- Muñoz-San Martín, C.; Apt, W.; Zulantay, I. Real-time PCR strategy for the identification of Trypanosoma cruzi discrete typing units directly in chronically infected human blood. Infect. Genet. Evol. 2017, 49, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Kooy, R.F.; Ashall, F.; Van der Ploeg, M.; Overdulve, J.P. On the DNA content of Trypanosoma cruzi. Mol. Biochem. Parasitol. 1989, 36, 73–76. [Google Scholar] [CrossRef]
- Pycke, J.R. Some tests for uniformity of circular distributions powerful against multimodal alternatives. Can. J. Stat. 2010, 38, 80–96. [Google Scholar] [CrossRef]
- Bravo, N.; Muñoz, C.; Nazal, N.; Saavedra, M.; Martínez, G.; Araya, E.; Apt, W.; Zulantay, I. Real-Time PCR in faecal samples of Triatoma infestans obtained by xenodiagnosis: Proposal for an exogenous internal control. Parasites Vectors 2012, 5, 59. [Google Scholar] [CrossRef]
- Saavedra, M.; Zulantay, I.; Apt, W.; Castillo, J.; Araya, E.; Martínez, G.; Rodríguez, J. Quantification by real-time PCR of Trypanosoma cruzi DNA in samples of Triatoma infestans used in xenodiagnosis of chronic Chagas disease patients. Parasites Vectors 2016, 9, 382. [Google Scholar] [CrossRef]
- Dias, F.D.A.; Guerra, B.; Vieira, L.R.; Perdomo, H.D.; Gandara, A.C.P.; Amaral, R.J.V.D.; Vollú, R.E.; Gomes, S.A.O.; Lara, F.A.; Sorgine, M.H.F.; et al. Monitoring of the parasite load in the digestive tract of Rhodnius prolixus by combined qPCR analysis and imaging techniques provides new insights into the trypanosome life cycle. PLoS Negl. Trop. Dis. 2015, 9, e0004186. [Google Scholar] [CrossRef]
- Mateus, J.; Guerrero, P.; Lasso, P.; Cuervo, C.; González, J.M.; Puerta, C.J.; Cuéllar, A. An animal model of acute and chronic chagas disease with the reticulotropic Y strain of Trypanosoma cruzi that depicts the multifunctionality and dysfunctionality of T cells. Front. Immunol. 2019, 10, 918. [Google Scholar] [CrossRef] [PubMed]
- Campos, R.; Botto-Mahan, C.; Ortiz, S.; Acuña, M.; Cattan, P.E.; Solari, A. Trypanosoma cruzi detection in blood by xenodiagnosis and polymerase chain reaction in the wild rodent Octodon degus. Am. J. Trop. Med. Hyg. 2007, 76, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Schenone, H. Xenodiagnosis. Mem. Inst. Oswaldo Cruz 1999, 94, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Yefi-Quinteros, E.; Muñoz-San Martín, C.; Bacigalupo, A.; Correa, J.P.; Cattan, P.E. Trypanosoma cruzi load in synanthropic rodents from rural areas in Chile. Parasites Vectors 2018, 11, 171. [Google Scholar] [CrossRef]
- Enriquez, G.F.; Garbossa, G.; Macchiaverna, N.P.; Argibay, H.D.; Bua, J.; Gürtler, R.E.; Cardinal, M.V. Is the infectiousness of dogs naturally infected with Trypanosoma cruzi associated with poly-parasitism? Vet. Parasitol. 2016, 223, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Macchiaverna, N.P.; Enriquez, G.F.; Bua, J.; Fernández, M.P.; Sartor, P.A.; Gürtler, R.E.; Cardinal, M.V. Human infectiousness and parasite load in chronic patients seropositive for Trypanosoma cruzi in a rural area of the Argentine Chaco. Infect. Genet. Evol. 2020, 78, 104062. [Google Scholar] [CrossRef]
- Añez, N.; East, J.S. Studies on Trypanosoma rangeli Tejera, 1920 II. Its effect on feeding behaviour of triatomine bugs. Acta Trop. 1984, 41, 93–95. [Google Scholar] [CrossRef]
- Canals, M.; Solis, R.; Valderas, J.; Ehrenfeld, M.; Cattan, P.E. Preliminary studies on temperature selection and activity cycles of Triatoma infestans and T. spinolai (Heteroptera:Reduviidae), Chilean vectors of Chagas’ disease. J. Med. Entomol. 1997, 34, 11–17. [Google Scholar] [CrossRef]
- Moore, J. Altered behavior in cockroaches (Periplaneta americana) infected with an archiacanthocephalan, Moniliformis moniliformis. J. Parasitol. 1983, 69, 1174–1176. [Google Scholar] [CrossRef]
- Ponton, F.; Lefevre, T.; Lebarbenchon, C.; Thomas, F.; Loxdale, H.D.; Marché, L.; Renault, L.; Perrot-Minnot, M.J.; Biron, D.G. Do distantly related parasites rely on the same proximate factors to alter the behaviour of their hosts? Proc. Biol. Sci. 2006, 273, 2869–2877. [Google Scholar] [CrossRef] [PubMed]
- Helluy, S.; Thomas, F. Parasitic manipulation and neuroinflammation: Evidence from the system Microphallus papillorobustus (Trematoda)—Gammarus (Crustacea). Parasites Vectors 2010, 3, 38. [Google Scholar] [CrossRef] [PubMed]
- Blenau, W.; Thamm, M. Distribution of serotonin (5-HT) and its receptors in the insect brain with focus on the mushroom bodies: Lessons from Drosophila melanogaster and Apis mellifera. Arthropod Struct. Dev. 2011, 40, 381–394. [Google Scholar] [CrossRef]
- Müller, U. The nitric oxide system in insects. Prog. Neurobiol. 1997, 51, 363–381. [Google Scholar] [CrossRef]
- Evans, O.; Caragata, E.P.; Mcmeniman, C.J.; Woolfit, M.; Green, D.C.; Williams, C.R.; Franklin, C.E.; O’Neill, S.L.; Mcgraw, E.A. Increased locomotor activity and metabolism of Aedes aegypti infected with a life-shortening strain of Wolbachia pipientis. J. Exp. Biol. 2009, 212, 1436–1441. [Google Scholar] [CrossRef]
- Lima-Camara, T.N.; Bruno, R.V.; Luz, P.M.; Castro, M.G.; Lourenço-De-Oliveira, R.; Sorgine, M.H.F.; Peixoto, A.A. Dengue infection increases the locomotor activity of Aedes aegypti females. PLoS ONE 2011, 6, e17690. [Google Scholar] [CrossRef]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef]
- Tallon, A.K.; Lorenzo, M.G.; Moreira, L.A.; Martinez Villegas, L.E.; Hill, S.R.; Ignell, R. Dengue infection modulates locomotion and host seeking in Aedes aegypti. PLoS Negl. Trop. Dis. 2020, 14, e0008531. [Google Scholar] [CrossRef]
- Lee, J.H.; Rowley, W.A.; Platt, K.B. Vector competence of Culex tarsalis (Diptera: Culicidae) from Iowa for a sympatric strain of western equine encephalomyelitis (WEE-7738) virus. J. Med. Entomol. 2000, 37, 194–197. [Google Scholar] [CrossRef]
- Han, Y.; Van Oers, M.M.; Van Houte, S.; Ros, V.I.D. Virus-induced behavioural changes in insects. In Host Manipulations by Parasites and Viruses; Springer: Cham, Switzerland, 2015; pp. 149–174. [Google Scholar]
- Lazzari, C.R.; Fischbein, D.; Insausti, T.C. Differential control of light-dark adaptation in the ocelli and compound eyes of Triatoma infestans. J. Insect Physiol. 2011, 57, 1545–1552. [Google Scholar] [CrossRef]
- Lazzari, C.R. Circadian organization of locomotion activity in the haematophagous bug Triatoma infestans. J. Insect Physiol. 1992, 38, 895–903. [Google Scholar] [CrossRef]
- Lazzari, C.R. The behaviour of kissing Bugs. In Triatominae-The Biology of Chagas Disease Vectors; Springer: Cham, Switzerland, 2021; pp. 215–238. [Google Scholar] [CrossRef]
- Peterson, J.K.; Graham, A.L.; Dobson, A.P.; Chávez, O.T. Rhodnius prolixus life history outcomes differ when infected with different Trypanosoma cruzi I strains. Am. J. Trop. Med. Hyg. 2015, 93, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Coura, J.R. Transmission of chagasic infection by oral route in the natural history of Chagas disease. Rev. Soc. Bras. Med. Trop. 2006, 39 (Suppl. 3), 113–117. [Google Scholar] [PubMed]
- Silva-Dos-Santos, D.; Barreto-de-Albuquerque, J.; Guerra, B.; Moreira, O.C.; Berbert, L.R.; Ramos, M.T.; Mascarenhas, B.A.S.; Britto, C.; Morrot, A.; Serra Villa-Verde, D.M.; et al. Unraveling Chagas disease transmission through the oral route: Gateways to Trypanosoma cruzi infection and target tissues. PLoS Negl. Trop. Dis. 2017, 11, e0005507. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parasite-Equivalents/ Mg of Triatoma infestans DNA | N of Triatoma infestans |
|---|---|
| <1 | 1 |
| 1–<10 | 8 |
| 10–<100 | 6 |
| 100–<1000 | 6 |
| 1000–<10,000 | 12 |
| 10,000–<100,000 | 17 |
| 100,000–>100,000 | 6 |
| Infection Status | N | Units | Median | Mean | SE | Min | Max | |
|---|---|---|---|---|---|---|---|---|
| Movement events in photophase | Non-infected | 52 | Events | 589.5 | 635.88 | 39.16 | 168 | 1585 |
| Infected | 56 | Events | 1596 | 1853.69 | 99.99 | 846 | 3753 | |
| Movement events in scotophase | Non-infected | 52 | Events | 664.5 | 1059.1 | 161.13 | 22 | 4808 |
| Infected | 56 | Events | 1328 | 1314.89 | 110.69 | 109 | 3190 | |
| Movement events total | Non-infected | 52 | Events | 1293.5 | 1694.98 | 182.94 | 258 | 6232 |
| Infected | 56 | Events | 3188 | 3168.57 | 167.02 | 1180 | 5829 | |
| Distance traveled in photophase | Non-infected | 52 | cm | 1340.4 | 1406.37 | 54.43 | 743.8 | 2191.1 |
| Infected | 56 | cm | 2895.1 | 3514.61 | 242.08 | 1063.4 | 8735.2 | |
| Distance traveled in scotophase | Non-infected | 52 | cm | 1501.55 | 1673.48 | 166.44 | 321.6 | 5603.2 |
| Infected | 56 | cm | 2221.35 | 2826.7 | 293.33 | 513.9 | 13,533.4 | |
| Distance traveled total | Non-infected | 52 | cm | 2672.15 | 3079.85 | 190.32 | 1299.3 | 7573 |
| Infected | 56 | cm | 5724.15 | 6341.31 | 442.87 | 2008.9 | 17,039.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chacón, F.; Muñoz-San Martín, C.; Bacigalupo, A.; Álvarez-Duhart, B.; Solís, R.; Cattan, P.E. Trypanosoma cruzi Parasite Load Modulates the Circadian Activity Pattern of Triatoma infestans. Insects 2022, 13, 76. https://doi.org/10.3390/insects13010076
Chacón F, Muñoz-San Martín C, Bacigalupo A, Álvarez-Duhart B, Solís R, Cattan PE. Trypanosoma cruzi Parasite Load Modulates the Circadian Activity Pattern of Triatoma infestans. Insects. 2022; 13(1):76. https://doi.org/10.3390/insects13010076
Chicago/Turabian StyleChacón, Francisco, Catalina Muñoz-San Martín, Antonella Bacigalupo, Bárbara Álvarez-Duhart, Rigoberto Solís, and Pedro E. Cattan. 2022. "Trypanosoma cruzi Parasite Load Modulates the Circadian Activity Pattern of Triatoma infestans" Insects 13, no. 1: 76. https://doi.org/10.3390/insects13010076
APA StyleChacón, F., Muñoz-San Martín, C., Bacigalupo, A., Álvarez-Duhart, B., Solís, R., & Cattan, P. E. (2022). Trypanosoma cruzi Parasite Load Modulates the Circadian Activity Pattern of Triatoma infestans. Insects, 13(1), 76. https://doi.org/10.3390/insects13010076