
Ultrastructure of Antennal Sensilla in Adults of Dioryctria rubella Hampson (Lepidoptera: Pyralidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Scanning Electron Microscopy (SEM)
2.3. Terminology
2.4. Measurements and Statistical Analysis
3. Results
3.1. General Morphology of Antennal Sensilla in D. rubella Adults
3.2. Features of Sensilla Trichodea
3.3. Features of Sensilla Coeloconica
3.4. Features of Sensilla Auricillica
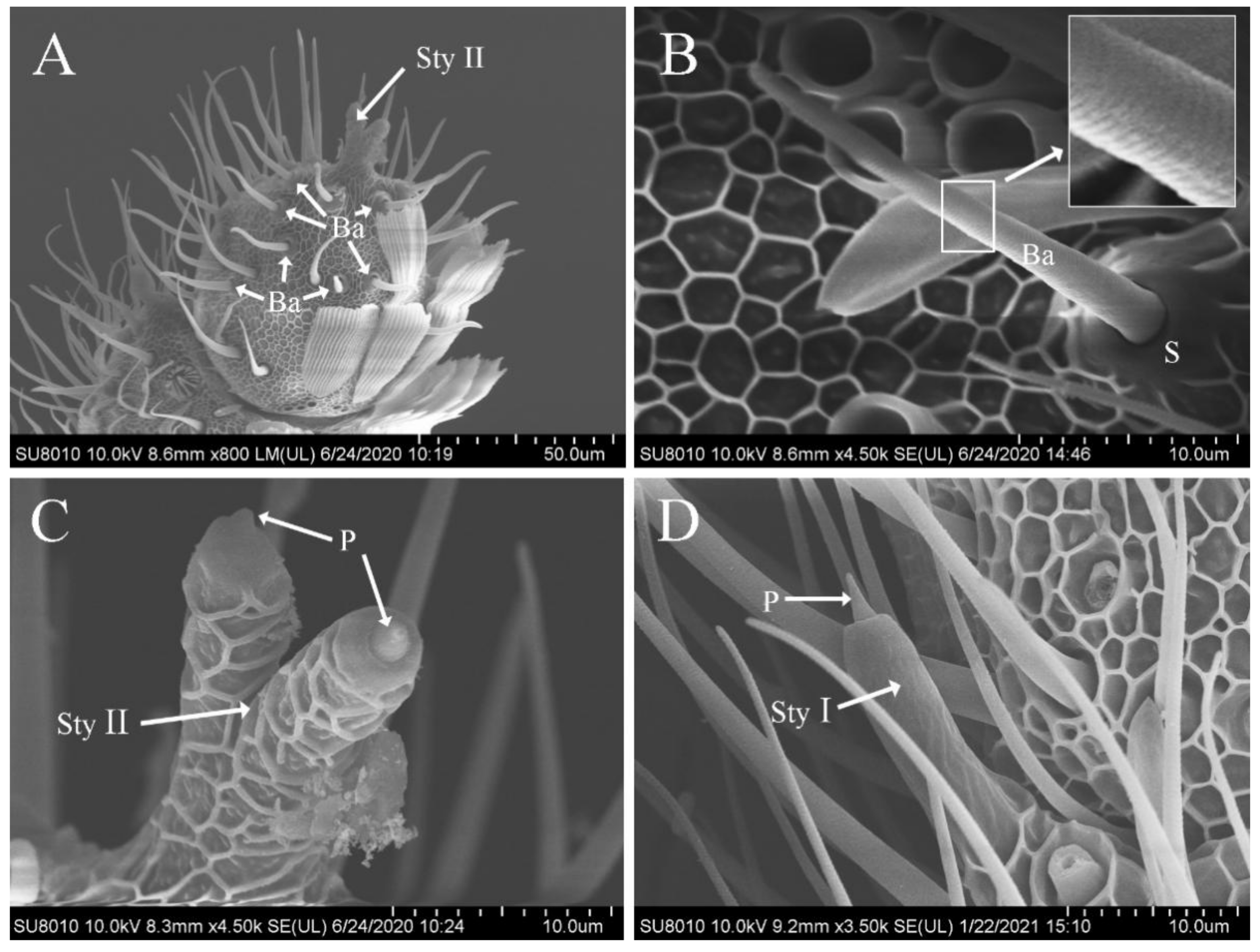
3.5. Features of Sensilla Basiconica
3.6. Features of Sensilla Styloconica
3.7. Features of Böhm’s Bristles
3.8. Features of Sensilla Squamiformia
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galizia, C.; Rössler, W. Parallel olfactory systems in insects: Anatomy and function. Annu. Rev. Entomol. 2010, 55, 399–420. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.R.; Guo, Y.Y.; Wu, K.M. Study on the ultrastructures of antennal sensilla in Helicoverpa armigera. Sci. Agric. Sin. 2002, 35, 1479–1482. [Google Scholar]
- Ma, R.Y.; Du, J.W. Insect antennae. Entomol. Knowl. 2000, 37, 179–183. [Google Scholar]
- Gao, J.Y.; Yu, B.Z.; Liu, S.W.; Wang, G.X.; Kuang, M.H.; Gao, Y.G. Population composition and niche of arthropod community in pine shoot tunnel bored by Dioryctria rubella. Chin. J. Ecol. 2010, 29, 363–369. [Google Scholar]
- Xin, H.P.; Zhang, J.T. Research advances on Dioryctria rubella Hampson. Mod. Agric. Sci. Technol. 2011, 8, 171–172. [Google Scholar]
- Wen, J.B. Effect of damage caused by Dioryctria rubella Hampson on the growth of Pinus tabulaeformis Carr. For. Pest Dis. 1998, 2, 1–2. [Google Scholar]
- Liang, J.S.; Wang, X.; Tan, X.H.; Tong, X.W.; Xia, Y.G.; Zhou, G.; Wang, X.L. Effect of damage by Dioryctria rubella on the growth of young Pinus massoniana forest and shoot cutting technique to control D. rubella. Hunan For. Sci. Technol. 2011, 1, 18–21. [Google Scholar]
- Schneider, D. Insect antennae. Ann. Rev. Entomol. 1964, 9, 103–122. [Google Scholar] [CrossRef]
- Steinbrecht, R.A.; Laue, M.; Maida, R.; Ziegelberger, G. Odorant-binding proteins and their role in the detection of plant odors. Entomol. Exp. Appl. 1996, 80, 15–18. [Google Scholar] [CrossRef]
- Forstner, M.; Breer, H.; Krieger, J. A receptor and binding protein interplay in the detection of a distinct pheromone component in the silkmoth Antheraea polyphemus. Int. J. Biol. Sci. 2009, 5, 745–757. [Google Scholar] [CrossRef] [Green Version]
- Forstner, M.; Gohl, T.; Breer, H.; Krieger, J. Candidate pheromone binding proteins of the silkmoth Bombyx mori. Invertebr. Neurosci. 2006, 6, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.J.; Tang, R.; Guo, H.; Ning, C.; Li, J.C.; Wu, H.; Wang, C.Z. Olfactory coding of intra- and interspecific pheromonal messages by the male Mythimna separata in North China. Insect Biochem. Mol. Biol. 2020, 125, 103439. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Wang, M.Q.; Zhang, G. Ultrastructural observations on antennal sensilla of Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Microsc. Res. Tech. 2011, 74, 113–121. [Google Scholar] [CrossRef]
- Maida, R.; Mameli, M.; Müller, B.; Krieger, J.; Steinbrecht, R.A. The expression pattern of four odorant-binding proteins in male and female silk moths, Bombyx mori. J. Neurocytol. 2005, 34, 149–163. [Google Scholar] [CrossRef]
- Anfora, G.; Vitagliano, S.; Larsson, M.C.; Witzgall, P.; Tasin, M.; Germinara, G.S.; De Cristofaro, A. Disruption of Phthorimaea operculella (Lepidoptera: Gelechiidae) oviposition by the application of host plant volatiles. Pest Manag. Sci. 2014, 70, 628–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wee, S.L.; Oh, H.W.; Park, K.C. Antennal sensillum morphology and electrophysiological responses of olfactory receptor neurons in trichoid sensilla of the diamondback moth (Lepidoptera: Plutellidae). Fla. Entomol. 2016, 99, 146–158. [Google Scholar] [CrossRef] [Green Version]
- Frank, D.L.; Leskey, T.C.; Bergh, J.C. Morphological characterization of antennal sensilla of the dogwood borer (Lepidoptera: Sesiidae). Ann. Entomol. Soc. Am. 2010, 103, 993–1002. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.Z.; Deng, C.P.; Xie, J.X.; Wu, L.J.; Sun, X.J.; Hao, C. Distribution patterns and morphology of sensilla on the antennae of Plutella xylostella (L.)—A scanning and transmission electron microscopic study. Micron 2017, 103, 1–11. [Google Scholar] [CrossRef]
- Hunger, T.; Steinbrecht, R.A. Functional morphology of a double-walled multiporous olfactory sensillum: The sensillum coeloconicum of Bombyx mori (Insecta, Lepidoptera). Tissue Cell 1998, 30, 14–29. [Google Scholar] [CrossRef]
- Shanbhag, S.R.; Singh, K.; Singh, R.N. Fine structure and primary sensory projections of sensilla located in the sacculus of the antenna of Drosophila melanogaster. Cell Tissue Res. 1995, 282, 237–249. [Google Scholar] [CrossRef]
- Gao, Y.; Luo, L.Z.; Hammond, A. Antennal morphology, structure and sensilla distribution in Microplitis pallidipes (Hymenoptera: Braconidae). Micron 2007, 38, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Roux, O.; van Baaren, J.; Gers, C.; Arvanitakis, L.; Legal, L. Antennal structure and oviposition behavior of the Plutella xylostella specialist parasitoid: Cotesia plutellae. Microsc. Res. Tech. 2005, 6, 36–44. [Google Scholar] [CrossRef]
- Altner, H.; Schaller-Selzer, L.; Stetter, H.; Wohlrab, I. Poreless sensilla with inflexible sockets: A comparative study of a fundamental type of insect sensilla probably comprising thermo- and hygroreceptors. Cell Tissue Res. 1983, 234, 279–307. [Google Scholar] [CrossRef] [PubMed]
- Cuperus, P.L. Distribution of antennal sense organs in male and female ermine moth, Yponomeuta vigintipunctatus (Retzius) (Lepidoptera: Yponomeutidae). Int. J. Insect Morphol. 1983, 12, 59–66. [Google Scholar] [CrossRef]
- Anderson, P.; Hallberg, E.; Subchev, M. Morphology of antennal sensilla auricillica and their detection of plant volatiles in the Herald moth, Scoliopteryx libatrix L. (Lepidoptera: Noctuidae). Arthropod Struct. Dev. 2000, 29, 33–41. [Google Scholar] [CrossRef]
- Wall, C. Morphology and histology of the antenna of Cydia nigricana (F.) (Lepidoptera: Tortricidae). Int. J. Insect Morphol. 1978, 7, 237–250. [Google Scholar] [CrossRef]
- Ebbinghaus, D.; Lösel, P.M.; Lindemann, M.; Scherkenbeck, J.; Zebitz, C.P.W. Detection of major and minor sex pheromone components by the male codling moth Cydia pomonella (Lepidoptera: Tortricidae). J. Insect Physiol. 1997, 44, 49–58. [Google Scholar] [CrossRef]
- Larsson, M.C.; Hallberg, E.; Kozlov, M.V.; Francke, W.; Hansson, B.S.; Löfstedt, C. Specialized olfactory receptor neurons mediating intra-and interspecific chemical communication in leafminer moths Eriocrania spp. (Lepidoptera: Eriocraniidae). J. Exp. Biol. 2002, 205, 989–998. [Google Scholar] [CrossRef]
- Chapman, R.F. Chemoreception: The significance of receptor numbers. Adv. Insect Physiol. 1982, 16, 247–356. [Google Scholar]
- Laue, M. Immunolocalization of general odorant-binding protein in antennal sensilla of moth caterpillars. Arthropod Struct. Dev. 2000, 29, 57–73. [Google Scholar] [CrossRef]
- Lopes, O.; Barata, E.N.; Mustaparta, H.; Araújo, J. Fine structure of antennal sensilla basiconica and their detection of plant volatiles in the eucalyptus woodborer, Phoracantha semipunctata Fabricius (Coleoptera: Cerambycidae). Arthropod Struct. Dev. 2002, 31, 1–13. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, C.C.; Yan, S.W.; Liu, Y.; Guo, M.B.; Dong, S.L.; Wang, G.R. An odorant receptor from the common cutworm (Spodoptera litura) exclusively tuned to the important plant volatile cis-3-hexenyl acetate. Insect Mol. Biol. 2013, 22, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Castrejón Gómez, V.R.; Nieto, G.; Valdes, J.; Castrejón, F.; Rojas, J.C. The antennal sensilla of Zamagiria dixolophella Dyar (Lepidoptera: Pyralidae). Ann. Entomol. Soc. Am. 2003, 96, 672–678. [Google Scholar] [CrossRef]
- Steinbrecht, R.A. The fine structure of thermo-/hygrosensitive sensilla in the silkmoth Bombyx mori: Receptor membrane substructure and sensory cell contacts. Cell Tissue Res. 1989, 255, 49–57. [Google Scholar] [CrossRef]
- Faucheux, M.J. Morphology and distribution of sensilla on the cephalic appendages, tarsi and ovipositor of the European sunflower moth, Homoeosoma nebulella Den. & Schiff. (Lepidoptera: Pyralidae). Int. J. Insect Morphol. 1991, 20, 291–307. [Google Scholar]
- Merivee, E.; Ploomi, A.; Rahi, M.; Bresciani, J.; Ravn, H.P.; Luik, A.; Sammelselg, V. Antennal sensilla of the ground beetle Bembidion properans Steph. (Coleoptera, Carabidae). Micron 2002, 33, 429–440. [Google Scholar] [CrossRef]
- Yang, H.; Yan, S.C.; Liu, D. Ultrastructural observations on antennal sensilla of Coleophora obducta (Meyrick) (Lepidoptera: Coleophoridae). Micron 2009, 40, 231–238. [Google Scholar] [CrossRef]
- McIver, S.B. Structure of cuticular mechanoreceptors of arthropods. Annu. Rev. Entomol. 1975, 20, 381–397. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Antennae Length (μm) | Number of Flagellum Flagellomeres | Scape Length (μm) | Scape Width (μm) | Pedicel Length (μm) | Pedicel Width (μm) | Flagellum Length (μm) |
|---|---|---|---|---|---|---|---|
| Male | 6774.20 ± 193.27 a | 62 ± 0.32 a | 329.64 ± 32.17 a | 131.97 ± 14.54 a | 63.69 ± 4.35 a | 54.22 ± 4.51 a | 6,419.35 ± 129.24 a |
| Female | 6345.24 ± 131.38 b | 61 ± 0.40 a | 281.41 ± 29.74 b | 112.6 ± 15.83 b | 53.39 ± 3.24 b | 49.63 ± 3.23 b | 6,145.23 ± 186.14 b |
| Types of Sensilla | Length (μm) | Width (μm) | ||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| Trichodea | 51.04 ± 13.49 a | 46.45 ± 5.93 b | 2.75 ± 0.44 a | 2.50 ± 0.52 a |
| Coeloconica | 8.85 ± 0.57 a | 9.34 ± 0.89 a | -- | -- |
| Auricillica | 17.06 ± 2.94 a | 17.43 ± 2.95 a | 3.80 ± 0.77 a | 2.97 ± 0.51 a |
| Basiconica | 30.89 ± 3.12 b | 35.10 ± 4.42 a | 2.23 ± 0.26 b | 2.91 ± 0.46 a |
| StyloconicaⅠ | 19.69 ± 1.99 a | 19.58 ± 1.60 a | 3.91 ± 0.50 a | 4.12 ± 0.34 a |
| StyloconicaⅡ | -- | -- | -- | -- |
| Böhm′s bristles | 12.06 ± 1.64 a | 11.19 ± 1.77 a | 1.41 ± 0.16 a | 1.56 ± 0.17 a |
| Squamiformia | 24.41 ± 2.05 a | 25.82 ± 3.10 b | 2.32 ± 0.25 a | 2.47 ± 0.26 b |
| Types of Sensilla | Male | Female |
|---|---|---|
| Trichodea | 2980.6 ± 360.78 a | 1821.3 ± 162.50 b |
| Coeloconica | 287.0 ± 29.83 a | 249.4 ± 16.58 b |
| Auricillica | 1128.0 ± 98.45 a | 872 ± 79.86 b |
| Basiconica | 216.0 ± 23.08 a | 194.2 ± 28.14 a |
| StyloconicaⅠ | -- | -- |
| StyloconicaⅡ | -- | -- |
| Böhm’s bristles | -- | -- |
| Squamiformia | -- | -- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Deng, C.; Lu, W.; Wu, S. Ultrastructure of Antennal Sensilla in Adults of Dioryctria rubella Hampson (Lepidoptera: Pyralidae). Insects 2021, 12, 821. https://doi.org/10.3390/insects12090821
Xu J, Deng C, Lu W, Wu S. Ultrastructure of Antennal Sensilla in Adults of Dioryctria rubella Hampson (Lepidoptera: Pyralidae). Insects. 2021; 12(9):821. https://doi.org/10.3390/insects12090821
Chicago/Turabian StyleXu, Jin, Caiping Deng, Wenfeng Lu, and Sanan Wu. 2021. "Ultrastructure of Antennal Sensilla in Adults of Dioryctria rubella Hampson (Lepidoptera: Pyralidae)" Insects 12, no. 9: 821. https://doi.org/10.3390/insects12090821
APA StyleXu, J., Deng, C., Lu, W., & Wu, S. (2021). Ultrastructure of Antennal Sensilla in Adults of Dioryctria rubella Hampson (Lepidoptera: Pyralidae). Insects, 12(9), 821. https://doi.org/10.3390/insects12090821