
Serosurvey of Nonhuman Primates in Costa Rica at the Human–Wildlife Interface Reveals High Exposure to Flaviviruses

 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Protocol
2.2. Serological Screening
3. Results
3.1. Serological Screening
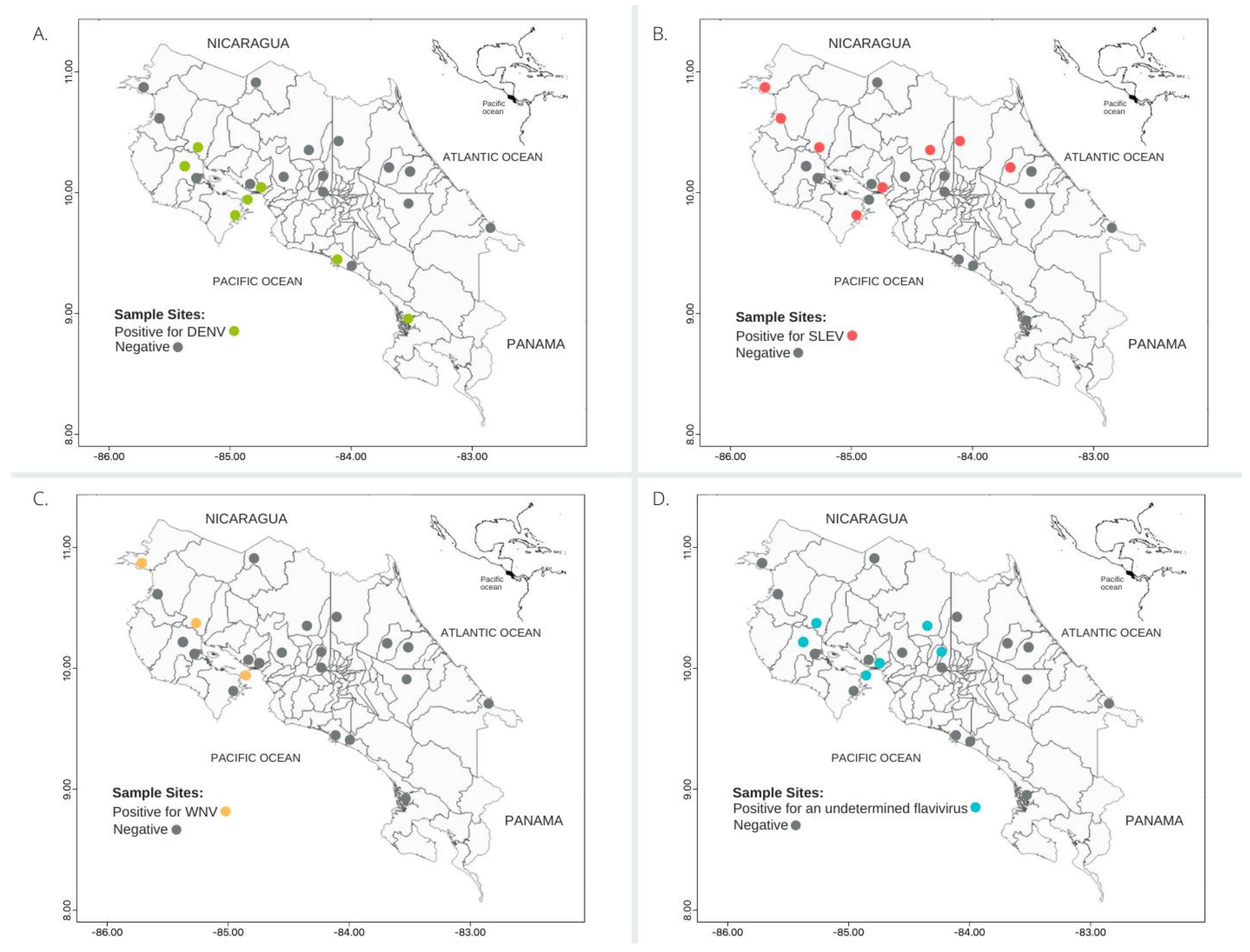
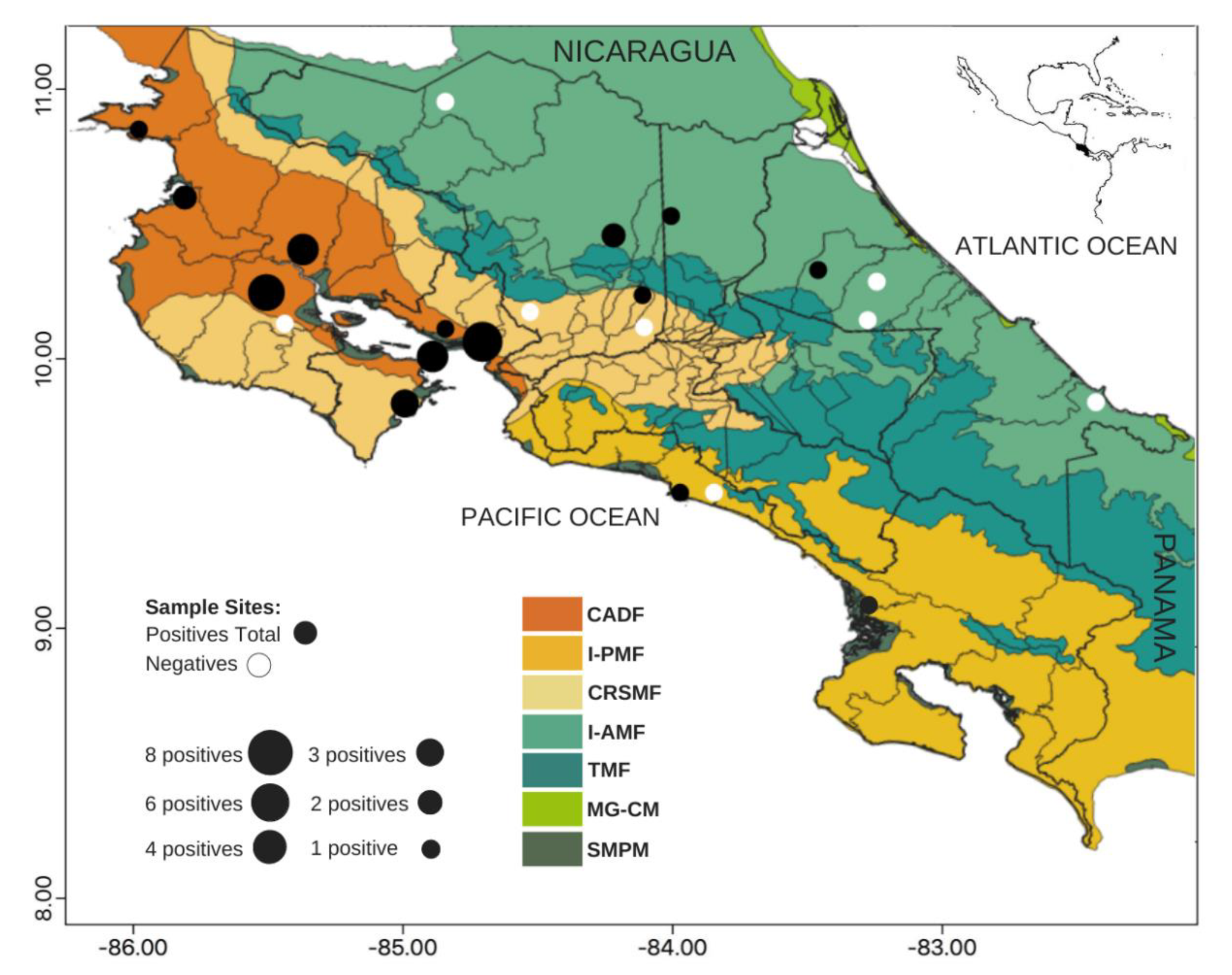
3.2. Geographical and Eco-Regional Distribution of Seropositives
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| NPs | neotropical primates |
| DENV | dengue virus |
| SLEV | Saint Louis encephalitis virus |
| WNV | West Nile virus |
| ZIKV | Zika virus |
| YFV | yellow fever virus |
| Micro-PRNT | micro plaque reduction neutralization test |
| HI | hemagglutination inhibition |
| CF | complement fixation |
| mg/kg | milligram per kilogram |
| m | meter |
| mL | milliliter |
| µL | microliter |
| PFU | plaque-forming units |
| mm | millimeter |
| °C | centigrade |
| RPM | revolutions per minute |
| min | minutes |
| MEM | minimal essential medium |
| CMC | carboxymethyl cellulose |
References
- Holbrook, M.R. Historical Perspectives on Flavivirus Research. Viruses 2017, 9, 97. [Google Scholar] [CrossRef]
- Kuno, G.; Gwong-Jen, J.; Tsuchiya, C.K.R.; Karabatsos, N.; Cropp, B. Phylogeny of the genus flavivirus. J. Virol. 1998, 72, 73–83. [Google Scholar] [CrossRef]
- Cleton, N.; Koopmans, M.; Reimerink, J.; Godeke, G.J.; Reusken, C. Come fly with me: Review of clinically important arboviruses for global travelers. J. Clin. Virol. 2012, 55, 191–203. [Google Scholar] [CrossRef]
- Musso, D.; Rodriguez-Morales, A.J.; Levi, J.E.; Cao-Lormeau, V.M.; Gubler, D.J. Unexpected outbreaks of arbovirus infections: Lessons learned from the Pacific and tropical America. The Lancet Infectious Diseases. Lancet Infect Dis. 2018, 355–361. [Google Scholar] [CrossRef]
- Araújo, P.A.; Freitas, M.O.; Oliveira Chiang, J.; Arruda Silva, F.; Leal Chagas, L.; Mansour Casseb, S.; Patroca Silva, S.; Pinto Nunes-Neto, J.; Rosa-Júnior, J.W.; Sena Nascimento, B.; et al. Investigation about the Occurrence of Transmission Cycles of Arbovirus in the Tropical Forest, Amazon Region. Viruses 2019, 11, 774. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, N.D.; Escalante, A.A.; Karesh, W.B.; Kilbourn, A.; Spielman, A.; Spielman, A.; Lal, A.A. Wild primate populations in emerging infectious disease research: The missing link? Emerg. Infect Dis. 1998, 4, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Valentine, M.; Cuin Murdock, C.; Kelly, P.J. Sylvatic cycles of arboviruses in non-human primates. Parasites Vectors 2019, 12, 463. [Google Scholar] [CrossRef] [PubMed]
- Pandit, P.S.; Doyle, M.M.; Smart, K.M.; Young, C.C.W.; Drape, G.W.; Johnson, C.K. Predicting wildlife reservoirs and global vulnerability to zoonotic Flaviviruses. Nat. Commun. 2018, 9, 5425. [Google Scholar] [CrossRef]
- Chippaux, J.P.; Chippaux, A. Yellow fever in Africa and the Americas: A historical and epidemiological perspective. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 20. [Google Scholar] [CrossRef]
- Hanley, K.A.; Monath, T.P.; Weaver, S.C.; Rossi, S.L.; Richman, R.L.; Vasilakis, N. Fever versus fever: The role of host and vector susceptibility and interspecific competition in shaping the current and future distributions of the sylvatic cycles of dengue virus and yellow fever virus. Infect. Genet. Evol. 2013, 19, 292–311. [Google Scholar] [CrossRef]
- Morales, M.A.; Fabbri, C.M.; Zunino, G.E.; Kowalewski, M.M.; Luppo, V.C.; Enría, D.A.; Levis, S.C.; Calderon, G.E. Detection of the mosquito-borne flaviviruses, West Nile, dengue, Saint Louis encephalitis, Ilheus, Bussuquara, and yellow fever in free-ranging black howlers (Alouatta caraya) of northeastern Argentina. PLoS Negl. Trop. Dis. 2017, 11, e0005351. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Soto, A.; Torres, M.C.; Lima de Mendonca, M.C.; Mares-Guia, M.A.; dos Santos Rodrigues, C.D.; Fabri, A.A.; dos Santos, C.C.; Machado, E.S.; Fischer, C.; Ribeiro, R.M.; et al. Evidence for multiple sylvatic transmission cycles during the 2016-2017 yellow fever virus outbreak. Braz. Clin. Microbiol Infect. 2018, 24, 1019-e1. [Google Scholar] [CrossRef] [PubMed]
- Dolz, G.; Chaves, A.; Gutiérrez-Espeleta, G.; Ortiz-Malavasi, E.; Bernal, S.; Herrero, M.V. Distribution of Flavivirus during time in wild non-human primates from lowlands from Costa Rica. PLoS ONE 2019, 14, e0219271. [Google Scholar] [CrossRef] [PubMed]
- Catenacci, L.S.; Ferreira, M.; Martins, L.C.; De Vleeschouwer, K.M.; Cassano, C.R.; Oliveira, L.C.; Canale, G.; Deem, S.L.; Tello, J.S.; Parker, P.; et al. Surveillance of arboviruses in primates and sloths in the Atlantic Forest, Bahia, Brazil. EcoHealth 2018, 15, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Soto, A.; Carneiro, I.D.O.; Fischer, C.; Feldmann, M.; Kümmerer, B.M.; Silva, N.S.; Santos, U.G.; Souza, B.F.D.C.D.; Liborio, F.D.A.; Valença Montenegro, M.M.; et al. Limited evidence for infection of urban and peri-urban nonhuman primates with Zika and chikungunya viruses in Brazil. mSphere 2018, 3, e00523-17. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Barrett, A.D.T. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Vinson, J.E.; Park, A.W. Vector-borne parasite invasion in communities across space and time. Proc. R. Soc. B 2019, 286, 20192614. [Google Scholar] [CrossRef] [PubMed]
- Lounibos, L.P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef]
- Mena, N.; Troyo, A.; Bonilla-Carrión, R.; Calderón-Arguedas, Ó. Factores asociados con la incidencia de dengue en Costa Rica. Rev. Panam. Salud. Publica 2011, 29, 234–242. [Google Scholar] [CrossRef]
- Varela, N. Introducción al Estudio de la salud de los Primates Neotropicales. 2007, Volume 56. Available online: https://www.veterinariosvs.org/introduccion-al-estudio-de-los-primates-neotropicales/ (accessed on 14 June 2021).
- West, G.; Heard, D.; Caulkett, N. Zoo Animal and Wildlife Immobilization and Anesthesia; John Wiley & Sons: Hoboken, NJ, USA, 2007; ISBN-10: 081381183X. [Google Scholar]
- Glander, K.E.; Fedigan, L.M.; Fedigan, L.; Chapman, C. Field methods for capture and measurement of three monkey spe- cies in Costa Rica. Folia Primatol. 1991, 57, 70–82. [Google Scholar] [CrossRef]
- Simões, M.; Camacho, L.A.; Yamamura, A.M.; Miranda, E.H.; Cajaraville, A.C.; da Silva Freire, M. Evaluation of accuracy and reliability of the plaque reduction neutralization test (micro-PRNT) in detection of yellow fever virus antibodies. Biologicals 2012, 40, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Guy, B.; Guirakhoo, F.; Barban, V.; Higgs, S.; Monath, T.P.; Lang, J. Preclinical and clinical development of YFV 17D-based chimeric vaccines against dengue, West Nile and Japanese encephalitis viruses. Vaccine 2010, 28, 632–649. [Google Scholar] [CrossRef]
- Azkarate, J.; Dunn, J.C.; Domingo Balcells, C.; Veà Baró, J. A demographic history of a population of howler monkeys (Alouatta palliata) living in a fragmented landscape in Mexico. PeerJ 2017, 5, e3547. [Google Scholar] [CrossRef] [PubMed]
- Maness, N.J.; Schouest, B.; Singapuri, A.; Dennis, M.; Gilbert, M.H.; Bohm, R.P. Postnatal Zika virus infection of nonhuman primate infants born to mothers infected with homologous Brazilian Zika virus. Sci. Rep. 2019, 9, 12802. [Google Scholar] [CrossRef] [PubMed]
- Díaz, L.A.; Ré, V.; Almirón, W.R.; Farías, A.; Vázquez, A.; Sanchez-Seco, M.P.; Aguilar, J.; Spinsanti, L.; Konigheim, B.; Visintin, A.; et al. Genotype III Saint Louis encephalitis virus outbreak, Argentina. Emerg. Infects Dis. 2006, 12, 1752–1754. [Google Scholar] [CrossRef]
- Heinen, L.B.; Zuchi, N.; Serra, O.P.; Cardoso, B.F.; Gondim, B.H.; Dos Santos, M.A.; Souto, F.J.; Paula, D.A.; Dutra, V.; Dezengrini-Slhessarenko, R. Saint Louis Encephalitis virus in Mato Grosso, Central-Western Brazil. Rev. Inst. Med. Trop. Sao Paulo 2015, 57, 215–220. [Google Scholar] [CrossRef]
- Medlin, S.; Deardorff, E.R.; Hanley, C.S.; Vergneau-Grosset, C.; Siudak-Campfield, A.; Dallwig, R.; Travassos da Rosa, A.; Tesh, R.B.; Martin, M.P.; Weaver, S.C.; et al. Serosurvey of Selected Arboviral Pathogens in Free-Ranging, Two-Toed Sloths (Choloepus Hoffmanni) And Three-Toed Sloths (Bradypus Variegatus) In Costa Rica, 2005–2007. J. Wildl Dis. 2016, 52, 883–892. [Google Scholar] [CrossRef]
- Kopp, A.; Gillespie, T.R.; Hobelsberger, D.; Estrada, A.; Harper, J.M.; Miller, R.A.; Eckerle, I.; Muller, M.A.; Podsiadlowski, L.; Leendertz, F.H.; et al. Provenance and geographic spread of St. Louis encephalitis virus. mBio 2013, 4, e00322-13. [Google Scholar] [CrossRef] [PubMed]
- Contigiani, M.S.; Fernandez, C.; Spinsanti, L.I.; D́ıaz, G.E. Prevalence of Flavivirus antibodies in Alouatta caraya primate autochthonous of Argentina. Medicina 2000, 60, 348–350. [Google Scholar]
- Svoboda, W.K.; Martins, L.C.; Malanski, L. deS.; Shiozawa, M.M.; Spohr, K.A.; Hilst, C.L.; Aguiar, L.M.; Ludwig, G.; Passos, F.deC.; Silva, L.R.; et al. Serological evidence for Saint Louis encephalitis virus in free-ranging New World monkeys and horses within the upper Paraná River basin region, Southern Brazil. Rev. Soc. Bras Med. Trop. 2014, 47, 280–286. [Google Scholar]
- Reisen, W.K. Epidemiology of St. Louis encephalitis virus. Adv. Virus Res. 2003, 61, 139–183. [Google Scholar]
- de Thoisy, B.; Dussart, P.; Kazanji, M. Wild terrestrial rainforest mammals as potential reservoirs for flaviviruses (yellow fever, dengue 2 and St Louis encephalitis viruses) in French Guiana. Trans. R Soc. Trop. Med. Hyg. 2004, 98, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Troyo, A.; Porcelain, S.L.; Calderón-Arguedas, O.; Chadee, D.D.; Beier, J.C. Dengue in Costa Rica: The gap in local scientific research. Rev. Panam. Salud. Publica 2006, 20, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Soto-Garita, C.; Somogyi, T.; Vicente-Santos, A.; Corrales-Aguilar, E. Molecular Characterization of Two Major Dengue Outbreaks in Costa Rica. Am. J. Trop. Med. Hyg. 2016, 95, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Health Ministry. Análisis de la Situación de Salud: Dengue. 2015. Available online: https://www.ministeriodesalud.go.cr/index.php/vigilancia-de-la-salud/analisis-de-situacion-de-salud?limit=20&limitstart=20 (accessed on 26 March 2021).
- Marin, R.; Diaz, M. Sitios de cría del Aedes aegypti en la region Pacífico Central de Costa Rica. Rev. Costarr. Salud. Publica 2012, 2, 21. [Google Scholar]
- de Figueiredo, M.L.; de C. Gomes, A.; Amarilla, A.A.; de S. Leandro, A.; de S. Orrico, A.; de Araujo, R.F.; do S. M. Castro, J.; Durigon, E.L.; Aquino, V.H.; Figueiredo, L.T. Mosquitoes infected with dengue viruses in Brazil. Virol. J. 2010, 7, 152. [Google Scholar] [CrossRef]
- Harris, M.; Caldwell, J.M.; Mordecai, E.A. Climate drives spatial variation in Zika epidemics in Latin America. Proc. R. Soc. B 2019, 286, 20191578. [Google Scholar] [CrossRef]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile Virus. Biomed. Res. Int. 2015, 376230. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Cerdeña, C.N.; Valiente-Banuet, L.; Sánchez-Cordero, V.; Stephens, C.R.; Ramsey, J.M. Trypanosoma cruzi reservoir—triatomine vector co-occurrence networks reveal meta-community effects by synanthropic mammals on geographic dispersal. PeerJ 2017, 5, e3152. [Google Scholar] [CrossRef] [PubMed]
- Mourão, M.P.G.; Bastos, M.S.; Gimaque, J.B.L.; Mota, B.R.; Souza, G.S.; Grimmer, G.H.N.; Galusso, E.S.; Arruda, E.; Figueiredo, L.T.M. Outbreak of Oropouche fever in Manaus, Western Brazilian Amazon, 2007-2008. Emerg. Infect. Dis. 2009, 15, 2063–2064. [Google Scholar] [CrossRef] [PubMed]
- Mourão, M.P.G.; Mota, B.R.; Souza, G.S.; Bastos, M.S.; Gimaque, J.B.L.; Galusso, E.S.; Kramer, V.M.; Figueiredo, L.T.M. Mayaro fever in Manaus, Brazil, 2007–2008. Vec. Born. Zoon. Dis. 2012, 12, 42–46. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Specie | n | DENV | SLEV | WNV | FLAVIRUS * |
|---|---|---|---|---|---|
| Alouatta palliata | 68 | 8 (0.12, 0.06–0.22) | 10 (0.15, 0.08–0.25) | 3 (0.04, 0.02–0.12) | 10 (0.15, 0.08–0.25) |
| Ateles geoffroyi | 14 | 2 (0.14, 0.04–0.4) | 1 (0.07, 0.01–0.31) | 0 (0, 0–0.22) | 3 (0.21, 0.08–0.48) |
| Saimiri oerstedii | 3 | 1 | 0 | 0 | 0 |
| Cebus imitator | 1 | 0 | 0 | 0 | 0 |
| Age | |||||
| adult | 62 | 11 (0.18, 0.10–0.29) | 10 (0.16, 0.09–0.27) | 3 (0.05, 0.02–0.13) | 11 (0.18, 0.10–0.29) |
| juvenile | 13 | 0 (0, 0–0.23) | 1 (0.08, 0.01–0.33) | 0 (0, 0–0.23) | 2 (15, 0.04–0.42) |
| not determined | 11 | 0 (0, 0–0.26) | 0 (0, 0–0.26) | 0 (0, 0–0.26) | 0 (0, 0–0.26) |
| Sex | |||||
| female | 46 | 3 (0.07, 0.02–0.18) | 7 (0.15, 0.08–0.28) | 0 (0, 0–0.08) | 5 (0.11, 0.05–0.23) |
| male | 38 | 8 (0.21, 0.11–0.36) | 4 (0.11, 0.04–0.24) | 3 (0.08, 0.03–0.21) | 8 (0.21, 0.11–0.36) |
| not determined | 2 | 0 | 0 | 0 | 0 |
| Animal Identification | Ecoregion | Species | Year | PRNT Titer | |||||
|---|---|---|---|---|---|---|---|---|---|
| DENV-1 | DENV-2 | DENV-3 | DENV-4 | SLEV | WNV | ||||
| AP-03 | I-AMF | A. palliata | 2000 | Neg | Neg | Neg | Neg | 1:160 * | Neg |
| AP-05 | CADF | A. palliata | 2000 | Neg | Neg | Neg | Neg | 1:160 * | Neg |
| AP-06 | CADF | A. palliata | 2001 | Neg | Neg | Neg | Neg | 1:80 * | Neg |
| AP-07 | CADF | A. palliata | 2001 | Neg | Neg | Neg | Neg | 1:40 * | Neg |
| AP-08 | CADF | A. palliata | 2001 | Neg | Neg | Neg | Neg | 1:40 * | Neg |
| AP-30 | CADF | A. palliata | 2001 | Neg | Neg | Neg | Neg | 1:40 * | Neg |
| AP-22 | CADF | A. palliata | 2001 | 1:80 † | 1:20 † | Neg | Neg | Neg | Neg |
| AP-12 | CADF | A. palliata | 2001 | Neg | 1:20 * | Neg | Neg | Neg | Neg |
| AP-19 | CADF | A. palliata | 2001 | Neg | 1:20 * | Neg | Neg | Neg | Neg |
| AP-27 | CADF | A. palliata | 2001 | Neg | Neg | Neg | Neg | Neg | 1:20 * |
| AP-09 | CADF | A. palliata | 2001 | 1:40 † | 1:20 † | Neg | Neg | Neg | Neg |
| AP-10 | CADF | A. palliata | 2001 | 1:20 † | 1:40 † | Neg | Neg | Neg | 1:20 † |
| AP-11 | CADF | A. palliata | 2001 | 1:40 † | 1:40 † | Neg | Neg | Neg | Neg |
| AP-58 | SMPM | A. palliata | 2002 | 1:20 * | Neg | Neg | Neg | Neg | Neg |
| AP-101 | CADF | A. palliata | 2003 | Neg | Neg | Neg | Neg | 1:160 * | 1:20 * |
| AP-144 | CADF | A. palliata | 2006 | 1:40 † | 1:20 † | 1:40 † | Neg | Neg | Neg |
| AP-142 | CADF | A. palliata | 2006 | Neg | Neg | Neg | Neg | Neg | 1:20 * |
| AP-147 | I-AMF | A. palliata | 2006 | Neg | Neg | Neg | Neg | 1:40 * | Neg |
| AG-50 | CRSMF | At. geoffroyi | 2006 | 1:20 † | Neg | Neg | Neg | 1:80 † | Neg |
| AG-45 | CRSMF | At. geoffroyi | 2006 | 1:20 * | Neg | Neg | Neg | Neg | Neg |
| AG-46 | CRSMF | At. geoffroyi | 2006 | Neg | Neg | Neg | Neg | 1:20 * | Neg |
| SM-1 | I-PMF | S. oerstedii | 2006 | 1:20 * | Neg | Neg | Neg | Neg | Neg |
| AG-55 | I-AMF | At. geoffroyi | 2006 | Neg | 1:20 † | Neg | Neg | 1:20 † | Neg |
| AG-55-1 | I-AMF | At. geoffroyi | 2006 | Neg | Neg | Neg | Neg | 1:80 † | 1:20 † |
| FP-101 | CADF | A. palliata | 2014 | 1:640 * | 1:20 * | Neg | Neg | Neg | Neg |
| FP-107 | CADF | A. palliata | 2014 | >1:1280 * | 1:20 † | Neg | Neg | Neg | 1:40 † |
| FP-108 | CADF | A. palliata | 2014 | 1:80 † | 1:20 † | Neg | Neg | Neg | 1:20 † |
| FPX-25 | CADF | A. palliata | 2014 | Neg | 1:20 * | Neg | Neg | Neg | Neg |
| FPX-21 | CADF | A. palliata | 2014 | Neg | 1:20 * | Neg | Neg | Neg | Neg |
| FP-109 | CADF | A. palliata | 2014 | Neg | 1:20 * | Neg | Neg | Neg | Neg |
| FP-102 | CADF | A. palliata | 2014 | 1:40 † | 1:20 † | Neg | Neg | Neg | Neg |
| FP-103 | CADF | A. palliata | 2014 | 1:80 † | 1:20 † | Neg | Neg | Neg | 1:40 † |
| MP-45 | CADF | A. palliata | 2015 | Neg | Neg | Neg | Neg | 1:80 * | Neg |
| MP-48 | CADF | A. palliata | 2015 | Neg | Neg | Neg | Neg | 1:40 * | Neg |
| MG-32 | CRSMF | At. geoffroyi | 2015 | Neg | 1:20 * | Neg | Neg | Neg | Neg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaves, A.; Piche-Ovares, M.; Ibarra-Cerdeña, C.N.; Corrales-Aguilar, E.; Suzán, G.; Moreira-Soto, A.; Gutiérrez-Espeleta, G.A. Serosurvey of Nonhuman Primates in Costa Rica at the Human–Wildlife Interface Reveals High Exposure to Flaviviruses. Insects 2021, 12, 554. https://doi.org/10.3390/insects12060554
Chaves A, Piche-Ovares M, Ibarra-Cerdeña CN, Corrales-Aguilar E, Suzán G, Moreira-Soto A, Gutiérrez-Espeleta GA. Serosurvey of Nonhuman Primates in Costa Rica at the Human–Wildlife Interface Reveals High Exposure to Flaviviruses. Insects. 2021; 12(6):554. https://doi.org/10.3390/insects12060554
Chicago/Turabian StyleChaves, Andrea, Martha Piche-Ovares, Carlos N. Ibarra-Cerdeña, Eugenia Corrales-Aguilar, Gerardo Suzán, Andres Moreira-Soto, and Gustavo A. Gutiérrez-Espeleta. 2021. "Serosurvey of Nonhuman Primates in Costa Rica at the Human–Wildlife Interface Reveals High Exposure to Flaviviruses" Insects 12, no. 6: 554. https://doi.org/10.3390/insects12060554
APA StyleChaves, A., Piche-Ovares, M., Ibarra-Cerdeña, C. N., Corrales-Aguilar, E., Suzán, G., Moreira-Soto, A., & Gutiérrez-Espeleta, G. A. (2021). Serosurvey of Nonhuman Primates in Costa Rica at the Human–Wildlife Interface Reveals High Exposure to Flaviviruses. Insects, 12(6), 554. https://doi.org/10.3390/insects12060554