
Effect of Temperature on the Development and Survival of Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae) Preying on Tetranychus urticae (Koch) (Acari: Tetranychidae)

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Their Preys
2.2. Development Experiment
2.3. Data Analysis and Model Development
- Effect of temperature on development time
- Development rate model
- (i)
- (ii)
- Logan-6 model [33]:
- (iii)
- Lactin-2 model [34]:
- (iv)
- (v)
- (vi)
- Briere-2 model [18]:
- Distribution of development completion
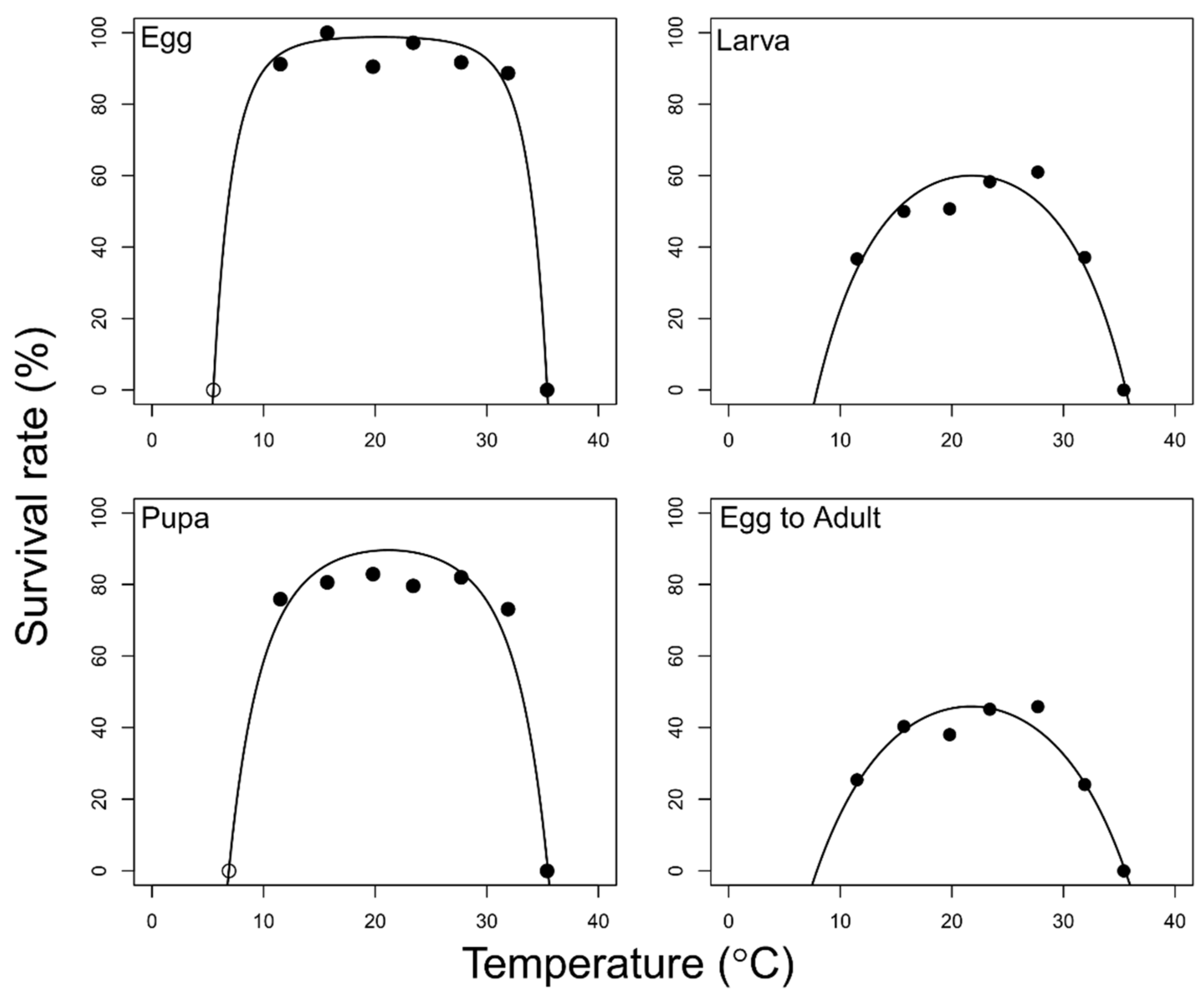
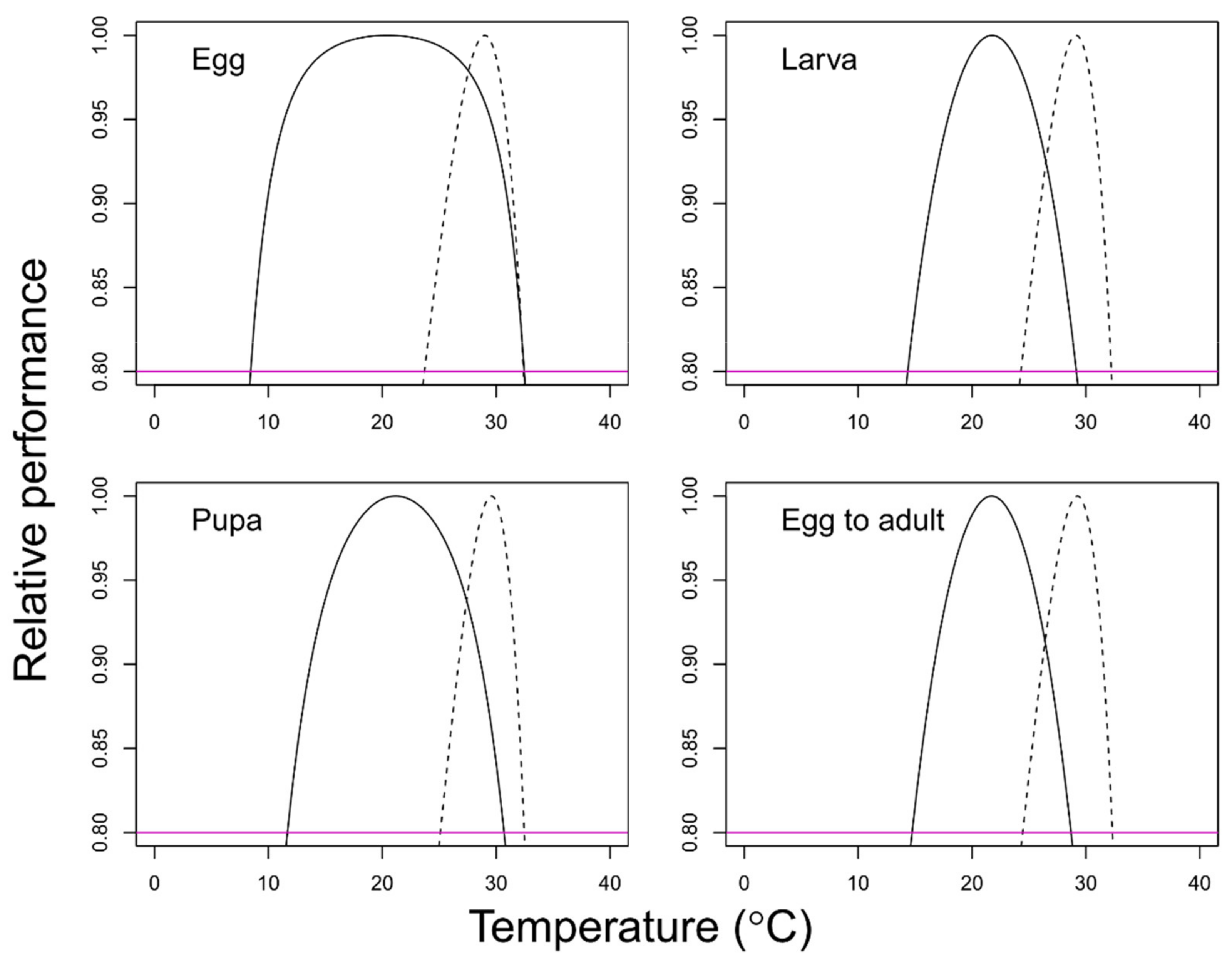
- Effect of temperature on survival
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CABI. Tetranychus urticae (two-spotted spider mite). In Invasive Species Compendium; CAB International: Wallingford, UK, 2019; Available online: https://www.cabi.org/isc/datasheet/53366 (accessed on 1 April 2021).
- Attia, S.; Grissa, K.L.; Lognay, G.; Bitume, E.; Hance, T.; Mailleux, A.C. A review of the major biological approaches to control the worldwide pest Tetranychus urticae (Acari: Tetranychidae) with special reference to natural pesticides. J. Pest Sci. 2013, 86, 361–386. [Google Scholar] [CrossRef]
- Kim, D.-S. Agro-ecosystem diversity and integrated mite pest management in fruit orchards: A review and future prospect. Korean J. Appl. Entomol. 2021, 60, 49–61. [Google Scholar]
- Goodwin, S.; Herron, G.; Gough, N.; Wellham, T.; Rophail, J.; Parker, R. Relationship between Insecticide-Acaricide Resistance and Field Control in Tetranychus urticae (Acari: Tetranychidae) Infesting Roses. J. Econ. Entomol. 1995, 88, 1106–1112. [Google Scholar] [CrossRef]
- Johnson, W.T.; Lyon, H.H. Insects that Feed on Trees and Shrubs, 2nd ed.; Cornell University Press: Ithaca, NY, USA, 1991. [Google Scholar]
- Georghiou, G.P.; Taylor, C.E. Genetic and Biological Influences in the Evolution of Insecticide Resistance. J. Econ. Entomol. 1977, 70, 319–323. [Google Scholar] [CrossRef]
- Choi, W.-I.; Lee, S.-G.; Park, H.-M.; Ahn, Y.-J. Toxicity of plant essential oils to Tetranychus urticae (Acari: Tetranychidae) and Phytoseiulus persimilis (Acari: Phytoseiidae). J. Econ. Entomol. 2004, 97, 553–558. [Google Scholar] [CrossRef] [PubMed]
- Pickett, C.H.; Gilstrap, F.E. Natural enemies associated with spider mites (Acari: Tetranychidae) infesting corn in the High Plains region of Texas. J. Kansas Entomol. Soc. 1986, 59, 524–536. [Google Scholar]
- Gillespie, D.R.; Roitberg, B.; Basalyga, E.; Johnstone, M.; Opit, G.; Rodgers, J.; Sawyer, N. Biology and Application of Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae) for Biological Control of Twospotted Spider Mites on Greenhouse Vegetable Crops; Technical Report 145 Agriculture and Agri-Food Canada; Pacific Agri-Food Research Centre: Agassiz, BC, Canada, 1998. [Google Scholar]
- Abe, J.; Ganaha-Kikumura, T.; Yukawa, J. Morphological features, distribution, prey mites, and life history traits of Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae) in Japan. Appl. Entomol. Zool. 2011, 46, 271–279. [Google Scholar] [CrossRef]
- Gagné, R.J. Revision of Tetranychid (Acarina) Mite Predators of the Genus Feltiella (Diptera: Cecidomyiidae). Ann. Entomol. Soc. Am. 1995, 88, 16–30. [Google Scholar] [CrossRef]
- Opit, G.P.; Roitberg, B.; Gillespie, D.R. The functional response and prey preference of Feltiella acarisuga (Vallot) (Diptera: Cecidomiidae) for two of its prey: Male and female twospotted spider mites, Tetranychus urticae Koch (Acari: Tetranychiidae). Can. Entomol. 1997, 129, 221–227. [Google Scholar] [CrossRef]
- Osborne, R.S.; Leppla, N.C.; Osborne, L.S. Predatory Gall Midge (unofficial common name), Feltiella acarisuga (Vallot) (Insecta: Diptera: Cecidomyiidae). EDIS 2002, 2004. [Google Scholar] [CrossRef]
- Mo, T.-L.; Liu, T.-X. Predation and life table of Feltiella acarisuga (Diptera: Cecidomyiidae) preying on eggs of Ttranychus urticae (Acari: Tetranychidae). Environ. Entomol. 2007, 35, 369–375. [Google Scholar] [CrossRef]
- Xiao, Y.; Osborne, L.S.; Chen, J.; Mckenzie, C.; Houben, K.; Irizarry, F. Evaluation of corn plant as potential banker plant for supporting predatory gall midge, Feltiella acarisuga (Diptera: Cecidomyiidae) against Tetranychus urticae (Acari: Tetranychidae) in greenhouse vegetable production. Crop Prot. 2011, 30, 1635–1642. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef]
- Taylor, F. Ecology and Evolution of Physiological Time in Insects. Am. Nat. 1981, 117, 1–23. [Google Scholar] [CrossRef]
- Briere, J.-F.; Pracros, P.; Le Roux, A.-Y.; Pierre, J.-S. A Novel Rate Model of Temperature-Dependent Development for Arthropods. Environ. Entomol. 1999, 28, 22–29. [Google Scholar] [CrossRef]
- Baek, S.; Son, Y.; Park, Y.-L. Temperature-dependent development and survival of Podisus maculiventris (Hemiptera: Pentatomidae): Implications for mass rearing and biological control. J. Pest Sci. 2014, 87, 331–340. [Google Scholar] [CrossRef]
- Agamy, E.A.; Gomma, W.O. Biological studies and food consumption on Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae) a predator of the spider mite, Tetranychus urticae (Koch) (Acari: Tetranychidae). Egypt. J. Biol. Pest Control 2002, 12, 87–89. [Google Scholar]
- Brødsgaard, H.F.; Jacobsen, S.; Enkegaard, A. Life-table characteristics of the predatory gall midge Feltiella acarisuga. Bull. OILB/SROC 1999, 22, 17–20. [Google Scholar]
- Svendsen, M.S.; Enkegaard, A.; Brødsgaard, H.F. Influence of humidity on the functional response of larvae of the fall midge (Feltiella acarisuga) feeding on spider mite eggs. Bull. OILB/SROC 1999, 22, 243–246. [Google Scholar]
- Kim, H.H.; Jeon, S.W.; Yoon, J.B.; Kim, D.H.; Yang, C.Y.; Kang, T.J. Effect of temperature on the biological attributes of the predatory gall midge, Feltiella acarisuga (Diptera: Cecidomyiidae). Korean J. Soil. Zool. 2015, 19, 15–21. [Google Scholar]
- Gillespie, D.; Opit, G.; Roitberg, B. Effects of Temperature and Relative Humidity on Development, Reproduction, and Predation in Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae). Biol. Control 2000, 17, 132–138. [Google Scholar] [CrossRef]
- Rafaei, G.S.; Mohamed, A.A. Biological characters of Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae) when fed on eggs of Tetranychus urticae Koach (Acari: Tetranychidae). Egypt. J. Agric. Res. 2013, 91, 119–123. [Google Scholar]
- Fratoni, S.; Duarte, M.V.A.; Vahansbeke, D.; Wäckers, F.L.; Dicke, M.; Pekas, A. A bittersweet meal: The impact of sugar solutions and honeydew on the fitness of two predatory gall midges. Biol. Control 2020, 140, 104098. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT User’s Guide, Cary version 9.3; SAS Institute: Cary, NC, USA, 2011. [Google Scholar]
- Davidson, J. On the Relationship between Temperature and Rate of Development of Insects at Constant Temperatures. J. Anim. Ecol. 1944, 13, 26–28. [Google Scholar] [CrossRef]
- Arnold, C.Y. The determination and significance of the base temperature in a linear heat unit system. Proc. Am. Soc. Hortic. Sci. 1959, 74, 430–445. [Google Scholar]
- Campbell, A.; Frazer, B.D.; Gilbert, N.; Gutierrez, A.P.; Mackauer, M. Temperature Requirements of Some Aphids and Their Parasites. J. Appl. Ecol. 1974, 11, 431–438. [Google Scholar] [CrossRef]
- Rosso, L.; Lobry, J.; Flandrois, J.-P. An Unexpected Correlation between Cardinal Temperatures of Microbial Growth Highlighted by a New Model. J. Theor. Biol. 1993, 162, 447–463. [Google Scholar] [CrossRef]
- Ratkowsky, D.A.; Reddy, G.V.P. Empirical Model with Excellent Statistical Properties for Describing Temperature-Dependent Developmental Rates of Insects and Mites. Ann. Entomol. Soc. Am. 2017, 110, 302–309. [Google Scholar] [CrossRef]
- Logan, J.A.; Wolkind, D.J.; Hoyt, S.C.; Tanigoshi, L.K. An analytic model for description of temperature dependent rate phenomena in Arthropods. Environ. Entomol. 1976, 5, 1133–1140. [Google Scholar] [CrossRef]
- Lactin, D.J.; Holliday, N.J.; Johnson, D.L.; Craigen, R. Improved rate model of temperature-dependent development by arthropods. Environ. Entomol. 1995, 24, 68–75. [Google Scholar] [CrossRef]
- Ratkowsky, D.A.; Lowry, R.K.; McMeekin, T.A.; Stokes, A.N.; Chandler, R.E. Model for bacterial culture growth rate throughout the entire biokinetic temperature range. J. Bacteriol. 1983, 154, 1222–1226. [Google Scholar] [CrossRef]
- Shi, P.; Ge, F.; Sun, Y.; Chen, C. A simple model for describing the effect of temperature on insect developmental rate. J. Asia Pac. Entomol. 2011, 14, 15–20. [Google Scholar] [CrossRef]
- Wang, L.F.; Shi, P.J.; Chen, C.; Xue, F.S. Effect of temperature on the development of Laodelphax striatellus (Homoptera: Delphacidae). J. Econ. Entomol. 2013, 106, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.Y.; Kropf, M.J.; McLaren, G.; Visperas, R.M. A nonlinear model for crop development as a function of temperature. Agric. For. Meteorol. 1995, 77, 1–16. [Google Scholar] [CrossRef]
- Shi, P.-J.; Reddy, G.V.P.; Chen, L.; Ge, F. Comparison of Thermal Performance Equations in Describing Temperature-Dependent Developmental Rates of Insects: (I) Empirical Models. Ann. Entomol. Soc. Am. 2016, 109, 211–215. [Google Scholar] [CrossRef]
- Lutterschmidt, W.I.; Hutchison, V.H. The critical thermal maximum: History and critique. Can. J. Zool. 1997, 75, 1561–1574. [Google Scholar] [CrossRef]
- Wagner, T.L.; Wu, H.I.; Sharpe, P.J.H.; Coulson, R.N. Modeling distribution of insect development rates: A literature review and application of the Weibull function. Environ. Entomol. 1984, 77, 475–487. [Google Scholar]
- Sánchez-Ramos, I.; Pascual, S.; Fernández, C.E.; Marcotegui, A.; González-Núňez, M. Effect of temperature on the survival and development of the immature stages of Monosteria unicostate (Hemiptera: Tingidae). Eur. J. Entomol. 2015, 112, 664–675. [Google Scholar] [CrossRef]
- Sharp, P.J.H.; DeMichele, D.W. Reaction kinetics of poikilotherm development. J. Theor. Biol. 1977, 64, 649–670. [Google Scholar] [CrossRef]
- Dixon, A.F.; Honěk, A.; Keil, P.; Kotela, M.A.A.; Šizling, A.L.; Jarošík, V. Relationship between the minimum and maximum temperature thresholds for development in insects. Funct. Ecol. 2009, 23, 257–264. [Google Scholar] [CrossRef]
- McCalla, K.A.; Keçeci, M.; Milosavljević, I.; Ratkowsky, D.A.; Hoddle, M.S. The Influence of temperature variation on life history parameters and thermal performance curves of Tamarixia radiata (Hymenoptera: Eulophidae), a parasitoid of the Asian citrus psyllid (Hemiptera: Liviidae). J. Econ. Entomol. 2019, 112, 1560–1574. [Google Scholar] [CrossRef] [PubMed]
- Milosavljević, I.; McCalla, K.A.; Morgan, D.J.W.; Hoddle, M.S. The effects of constant and fluctuating temperatures on development of Diaphorina citri (Hemiptera: Liviidae), the Asian citrus psyllid. J. Econ. Entomol. 2020, 113, 633–645. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temp (°C) | Egg | Larval | Pupal | Egg to Adult | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n 1 | d | s | n | d | s | n | d | s | n | d | s 4 | |
| 11.5 | 68 | 6.6 ± 0.10a 2 | 91.2 | 79 | 31.8 ± 2.05a | 36.7 | 29 | 17.8 ± 0.71a | 75.9 | 79 | 53.4 ± 1.88a | 25.4 |
| 15.7 | 99 | 4.2 ± 0.07b | 100.0 | 72 | 13.8 ± 0.56b | 50.0 | 36 | 10.7 ± 0.22b | 80.6 | 72 | 28.9 ± 0.59b | 40.3 |
| 19.8 | 84 | 2.8 ± 0.06c | 90.5 | 69 | 9.5 ± 0.28c | 50.7 | 35 | 6.5 ± 0.26c | 82.9 | 69 | 19.0 ± 0.27c | 38.0 |
| 23.4 | 36 | 2.3 ± 0.09d | 97.2 | 84 | 5.7 ± 0.22d | 58.3 | 49 | 4.7 ± 0.14d | 79.6 | 84 | 12.4 ± 0.27d | 45.1 |
| 27.7 | 72 | 1.8 ± 0.06e | 91.7 | 82 | 4.8 ± 0.14d | 61.0 | 50 | 4.0 ± 0.14d | 82.0 | 82 | 10.5 ± 0.16d | 45.8 |
| 31.9 | 53 | 1.7 ± 0.08e | 88.7 | 70 | 5.7 ± 0.12d | 37.1 | 26 | 3.5 ± 0.18d | 73.1 | 70 | 10.1 ± 0.24d | 24.1 |
| 35.4 | 139 | - 3 | 0.0 | 98 | - | 0.0 | 72 | - | 0.0 | 98 | - | 0.0 |
| Stage | Parameter (Mean ± SE) | Lower Development Threshold (°C) | Thermal Constant (DD) | |
|---|---|---|---|---|
| a | b | |||
| Egg | 0.024 ± 0.0007 | −0.131 ± 0.0158 | 5.5 | 41.7 |
| Larval | 0.011 ± 0.0010 | −0.105 ± 0.0207 | 9.5 | 90.9 |
| Pupal | 0.012 ± 0.0007 | −0.083 ± 0.0154 | 6.9 | 83.3 |
| Egg to adult | 0.005 ± 0.0004 | −0.041 ± 0.0075 | 8.2 | 200.0 |
| Model | Average Rank | Final Selection | ||||
|---|---|---|---|---|---|---|
| Egg | Larval | Pupal | Egg to Adult | |||
| LRF | 2533.6/43.4 | 23.7/35.8 | 705.2/44.5 | 50.9/39.8 | 4.5/1.8 | |
| Logan-6 | 67.8/35.4 | 5.1/35.0 | 21.2/35.0 | 1.4/35.1 | 2.5/1.0 | Selected |
| Lactin-2 | 2582.8/41.3 | 2.9/34.9 | 699.6/41.5 | 52.8/37.7 | 4.0/1.8 | |
| Performance-2 | 1228/36.9 | 4.9/33.4 | 631.0/38.7 | 0.6/35.1 | 2.5/1.5 | |
| Beta | 2842.8/43.8 | 2.5/35.3 | 780.5/45.1 | 51.2/38.9 | 4.5/1.8 | |
| Briere-2 | 2219.7/37.1 | 3.4/32.3 | 759.3/39.9 | 0.4/33.9 | 3.0/1.5 | |
| Model | Parameter | Egg | Larval | Pupal | Egg to Adult |
|---|---|---|---|---|---|
| LRF | 9.26 × 10−7 | 0.13 × 10−7 | 5.22 × 10−7 | 3.54 × 10−7 | |
| 19.0488 | 18.6848 | 20.7438 | 19.7650 | ||
| −5.3358 | 1.5729 | −2.9197 | −0.3839 | ||
| 43.4274 | 35.791 | 44.4095 | 39.8056 | ||
| Logan-6 | 0.1021 | 0.0433 | 0.0236 | 0.0087 | |
| 0.1496 | 0.1671 | 0.1774 | 0.1625 | ||
| 35.4276 | 35.0051 | 35.0338 | 35.0770 | ||
| 6.1817 | 5.7561 | 5.2619 | 5.5134 | ||
| Lactin-2 | 0.0851 | 0.1544 | 0.0939 | 0.1249 | |
| 42.7528 | 35.2245 | 42.7897 | 38.1585 | ||
| 11.0830 | 6.4580 | 10.4435 | 7.9811 | ||
| −0.2446 | −0.0312 | −0.1136 | −0.0221 | ||
| Performance-2 | 0.0252 | 0.0116 | 0.0131 | 0.0050 | |
| 5.6686 | 9.2970 | 7.7346 | 8.3325 | ||
| 0.3978 | 0.7557 | 0.3533 | 0.5603 | ||
| 36.9831 | 33.3749 | 38.7301 | 35.1266 | ||
| Beta | 0.5735 | 0.2142 | 0.2879 | 0.1012 | |
| 43.8128 | 35.3243 | 45.0534 | 38.8855 | ||
| 31.6761 | 28.6001 | 32.9301 | 30.1988 | ||
| −8.1044 | −18.4536 | −2.4553 | −10.7691 | ||
| Briere-2 | 2.88 × 10−4 | 2.58 × 10−4 | 1.22 × 10−4 | 0.93 × 10−4 | |
| −2.6248 | 6.3264 | 3.5766 | 4.1890 | ||
| 37.1227 | 32.3286 | 39.9349 | 33.9764 | ||
| 2.7946 | 4.6746 | 2.1764 | 3.9169 |
| Stage | Parameter | ||
| (Mean ± SE) | (Mean ± SE) | (Mean ± SE) | |
|---|---|---|---|
| Egg | 8.439 ± 0.5296 | −0.805 ± 0.1101 | 0.020 ± 0.0027 |
| Larval | 5.956 ± 0.3888 | −0.208 ± 0.0369 | 0.005 ± 0.0008 |
| Pupal | 7.665 ± 0.3879 | −0.501 ± 0.0584 | 0.012 ± 0.0014 |
| Egg to adult | 5.508 ± 0.2488 | −0.140 ± 0.0233 | 0.003 ± 0.0005 |
| Stage | Parameter | |
|---|---|---|
| α (Mean ± SE) | β (Mean ± SE) | |
| Egg | 1.038 ± 0.0108 | 7.839 ± 0.7049 |
| Larval | 1.078 ± 0.0082 | 5.433 ± 0.2982 |
| Pupal | 1.066 ± 0.0065 | 6.242 ± 0.3050 |
| Egg to adult | 1.039 ± 0.0043 | 10.534 ± 0.6176 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.-S.; Baek, S.; Kim, M.-J. Effect of Temperature on the Development and Survival of Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae) Preying on Tetranychus urticae (Koch) (Acari: Tetranychidae). Insects 2021, 12, 508. https://doi.org/10.3390/insects12060508
Choi Y-S, Baek S, Kim M-J. Effect of Temperature on the Development and Survival of Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae) Preying on Tetranychus urticae (Koch) (Acari: Tetranychidae). Insects. 2021; 12(6):508. https://doi.org/10.3390/insects12060508
Chicago/Turabian StyleChoi, Yong-Seok, Sunghoon Baek, and Min-Jung Kim. 2021. "Effect of Temperature on the Development and Survival of Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae) Preying on Tetranychus urticae (Koch) (Acari: Tetranychidae)" Insects 12, no. 6: 508. https://doi.org/10.3390/insects12060508
APA StyleChoi, Y.-S., Baek, S., & Kim, M.-J. (2021). Effect of Temperature on the Development and Survival of Feltiella acarisuga (Vallot) (Diptera: Cecidomyiidae) Preying on Tetranychus urticae (Koch) (Acari: Tetranychidae). Insects, 12(6), 508. https://doi.org/10.3390/insects12060508