
Patterns of Genetic Diversity and Mating Systems in a Mass-Reared Black Soldier Fly Colony


 , , , and
, , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Preparation
2.2. Genotyping
2.3. Genetic Data Analysis
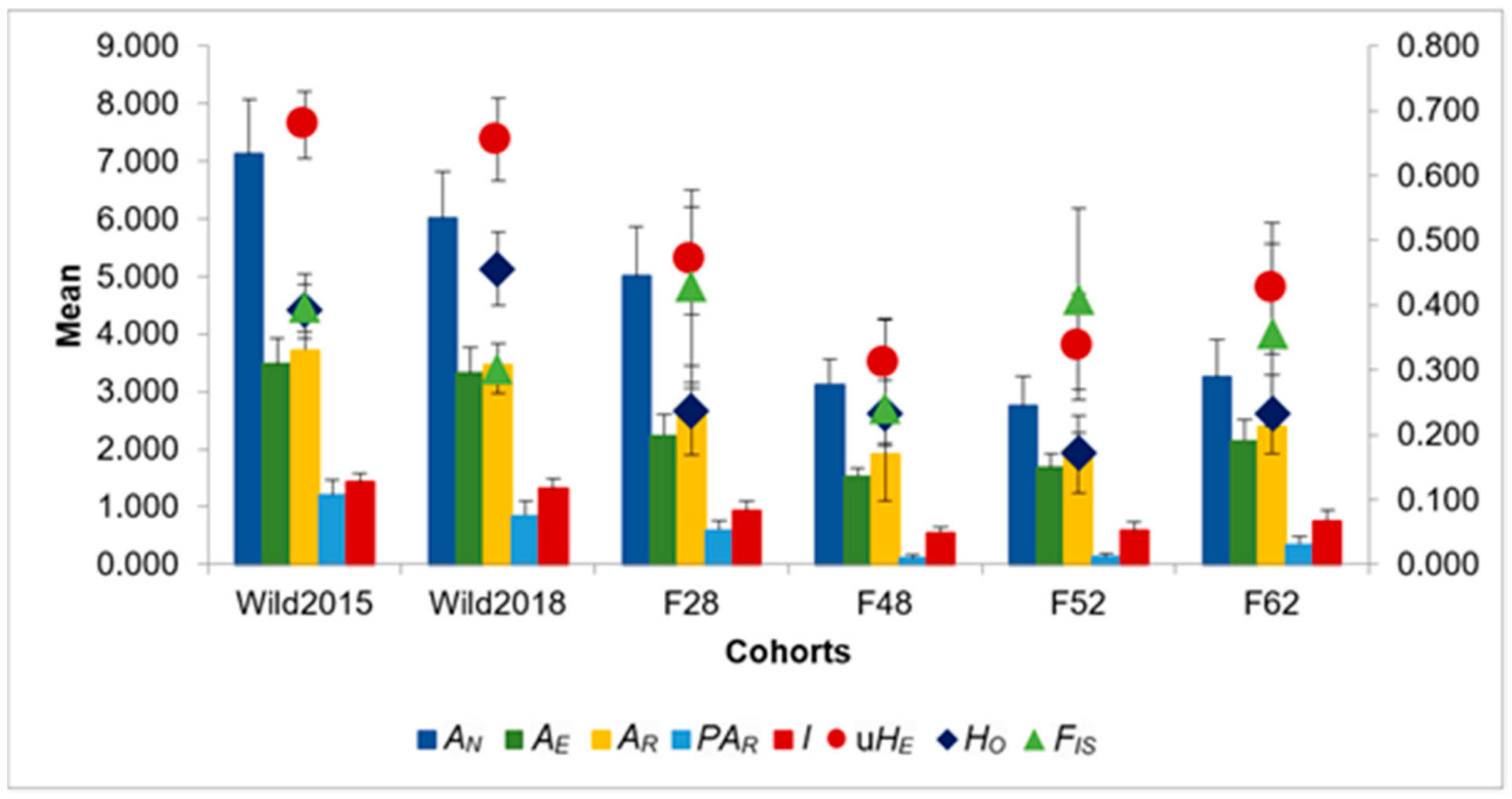
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; ISBN 9789251075951. [Google Scholar]
- Oonincx, D.G.A.B.; Van Broekhoven, S.; Van Huis, A.; Van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Yong, H.I.; Kim, Y.; Kim, H.; Choi, Y.-S. Edible insects as a protein source: A review of public perception, processing technology, and research trends. Food Sci. Anim. Resour. 2019, 39, 521–540. [Google Scholar] [CrossRef]
- Alliedmarketresearch Edible Insects Market by Product Type (Whole Insect, Insect Powder, Insect Meal, Insect Type (Crickets, Black Soldier Fly, Mealworms), Application (Animal Feed, Protein Bar and Shakes, Bakery, Confectionery, Beverages)—Global Forecast to 2030. Available online: https://www.researchandmarkets.com/reports/4757400/edible-insects-market-by-product-type-whole (accessed on 23 February 2021).
- Magalhães, R.; Sánchez-López, A.; Leal, R.S.; Martínez-Llorens, S.; Oliva-Teles, A.; Peres, H. Black soldier fly (Hermetia illucens) pre-pupae meal as a fish meal replacement in diets for European seabass (Dicentrarchus labrax). Aquaculture 2017, 476, 79–85. [Google Scholar] [CrossRef]
- Onsongo, V.O.; Osuga, I.M.; Gachuiri, C.K.; Wachira, A.M.; Miano, D.M.; Tanga, C.M.; Ekesi, S.; Nakimbugwe, D.; Fiaboe, K.K.M. Insects for income generation through animal feed: Effect of dietary replacement of soybean and fish meal with black soldier fly meal on broiler growth and economic performance. J. Econ. Entomol. 2018, 111, 1966–1973. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kortner, T.M.; Chikwati, E.M.; Belghit, I.; Lock, E.J.; Krogdahl, Å. Total replacement of fish meal with black soldier fly (Hermetia illucens) larvae meal does not compromise the gut health of Atlantic salmon (Salmo salar). Aquaculture 2020, 520, 734967. [Google Scholar] [CrossRef]
- Kaya, C.; Generalovic, T.N.; Ståhls, G.; Hauser, M.; Samayoa, A.C.; Nunes-Silva, C.G.; Roxburgh, H.; Wohlfahrt, J.; Ewusie, E.A.; Kenis, M.; et al. Global population genetic structure and demographic trajectories of the black soldier fly, Hermetia illucens. BMC Biol. 2021, 19, 94. [Google Scholar] [CrossRef]
- Salomone, R.; Saija, G.; Mondello, G.; Giannetto, A.; Fasulo, S.; Savastano, D. Environmental impact of food waste bioconversion by insects: Application of Life Cycle Assessment to process using Hermetia illucens. J. Clean. Prod. 2017, 140, 890–905. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Shelomi, M. Review of black soldier fly (Hermetia illucens) as animal feed and human food. Foods 2017, 6, 91. [Google Scholar] [CrossRef]
- Rhode, C.; Badenhorst, R.; Hull, K.L.; Greenwood, M.P.; Bester-van der Merwe, A.E.; Andere, A.A.; Picard, C.J.; Richards, C. Genetic and phenotypic consequences of early domestication in black soldier flies (Hermetia illucens). Anim. Genet. 2020, 51, 752–762. [Google Scholar] [CrossRef]
- Bijlsma, R.; Bundgaard, J.; Boerema, A.C. Does inbreeding affect the extinction risk of small populations? Predictions from Drosophila. J. Evol. Biol. 2000, 13, 502–514. [Google Scholar] [CrossRef]
- Mattila, H.R.; Seeley, T.D. Genetic diversity in honey bee colonies enhances productivity and fitness. Science 2007, 317, 362–364. [Google Scholar] [CrossRef]
- Hughes, A.R.; Inouye, B.D.; Johnson, M.T.J.; Underwood, N.; Vellend, M. Ecological consequences of genetic diversity. Ecol. Lett. 2008, 11, 609–623. [Google Scholar] [CrossRef]
- Ekroth, A.K.E.; Rafaluk-Mohr, C.; King, K.C. Host genetic diversity limits parasite success beyond agricultural systems: A meta-analysis. Proc. R. Soc. B Biol. Sci. 2019, 286. [Google Scholar] [CrossRef]
- James, J.W. The founder effect and response to artificial selection. Genet. Res. 1971, 16, 241–250. [Google Scholar] [CrossRef]
- Olesen, I.; Gjedrem, T.; Bentsen, H.B.; Gjerde, B.; Rye, M. Breeding programs for sustainable aquaculture. J. Appl. Aquac. 2003, 13, 179–204. [Google Scholar] [CrossRef]
- Gamborg, C.; Sandøe, P. Sustainability in farm animal breeding: A review. Livest. Prod. Sci. 2005, 92, 221–231. [Google Scholar] [CrossRef]
- Flint, A.P.F.; Woolliams, J.A. Precision animal breeding. Philos. Trans. R. Soc. B Biol. Sci. 2008, 363, 573–590. [Google Scholar] [CrossRef]
- Cardellino, R.A.; Boyazoglu, J. Research opportunities in the field of animal genetic resources. Livest. Sci. 2009, 120, 166–173. [Google Scholar] [CrossRef]
- Gavrilets, S.; Hastings, A. Founder effect speciation: A theoretical reassessment. Am. Nat. 1996, 147, 466–491. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Elam, D.R. Population genetic conseqences of small population size: Implications for plant conservation. Annu. Rev. Ecol. Syst. 1993, 24, 217–242. [Google Scholar] [CrossRef]
- Mignon-Grasteau, S.; Boissy, A.; Bouix, J.; Faure, J.M.; Fisher, A.D.; Hinch, G.N.; Jensen, P.; Le Neindre, P.; Mormède, P.; Prunet, P.; et al. Genetics of adaptation and domestication in livestock. Livest. Prod. Sci. 2005, 93, 3–14. [Google Scholar] [CrossRef]
- Hedrick, P.W.; Garcia-dorado, A. Understanding inbreeding depression, purging, and genetic rescue. Trends Ecol. Evol. 2016, 31, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Briscoe, D.A.; Malpica, J.M.; Robertson, A.; Smith, G.J.; Frankham, R.; Banks, R.G.; Barker, J.S.F. Rapid loss of genetic variation in large captive populations of Drosophila flies: Implications for the genetic management of captive populations. Conserv. Biol. 1992, 6, 416–425. [Google Scholar] [CrossRef]
- Cvijović, I.; Good, B.H.; Desai, M.M. The effect of strong purifying selection on genetic diversity. Genetics 2018, 209, 1235–1278. [Google Scholar] [CrossRef]
- Heber, S.; Briskie, J.V.; Apiolaza, L.A. A test of the “genetic rescue” technique using bottlenecked donor populations of Drosophila melanogaster. PLoS ONE 2012, 7, e43113. [Google Scholar] [CrossRef]
- Whiteley, A.R.; Fitzpatrick, S.W.; Funk, W.C.; Tallmon, D.A. Genetic rescue to the rescue. Trends Ecol. Evol. 2015, 30, 42–49. [Google Scholar] [CrossRef]
- Tallmon, D.A.; Luikart, G.; Waples, R.S. The alluring simplicity and complex reality of genetic rescue. Trends Ecol. Evol. 2004, 19, 489–496. [Google Scholar] [CrossRef]
- Kronenberger, J.A.; Gerberich, J.C.; Fitzpatrick, S.W.; Broder, E.D.; Angeloni, L.M.; Funk, W.C. An experimental test of alternative population augmentation scenarios. Conserv. Biol. 2018, 32, 838–848. [Google Scholar] [CrossRef]
- Baruch, O.; Mendel, Z.; Scharf, I.; Harari, A.R. Mating system, mate choice and parental care in a bark beetle. Bull. Entomol. Res. 2017, 107, 611–619. [Google Scholar] [CrossRef]
- Vargo, E.L. Diversity of termite breeding systems. Insects 2019, 10, 52. [Google Scholar] [CrossRef]
- Cotton, A.J.; Cotton, S.; Small, J.; Pomiankowski, A. Male mate preference for female eyespan and fecundity in the stalk-eyed fly, Teleopsis dalmanni. Behav. Ecol. 2015, 26, 376–385. [Google Scholar] [CrossRef]
- Gerofotis, C.D.; Yuval, B.; Ioannou, C.S.; Nakas, C.T.; Papadopoulos, N.T. Polygyny in the olive fly—Effects on male and female fitness. Behav. Ecol. Sociobiol. 2015, 69, 1323–1332. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Padoa-Schioppa, E.; Wang, J.; Garner, T.W.J. Polygyny, census and effective population size in the threatened frog, Rana latastei. Anim. Conserv. 2010, 13, 82–89. [Google Scholar] [CrossRef]
- Kvarnemo, C. Why do some animals mate with one partner rather than many? A review of causes and consequences of monogamy. Biol. Rev. 2018, 93, 1795–1812. [Google Scholar] [CrossRef]
- Arnqvist, R.A.N.; Nilsson, T. The evolution of polyandry: Multiple mating and female fitness in insects. Anim. Behav. 2000, 60, 145–164. [Google Scholar] [CrossRef]
- Jaffé, R. An updated guide to study polyandry in social insects. Sociobiology 2014, 61, 1–8. [Google Scholar] [CrossRef]
- Taylor, M.L.; Price, T.A.R.; Wedell, N. Polyandry in nature: A global analysis. Trends Ecol. Evol. 2014, 29, 376–383. [Google Scholar] [CrossRef]
- Flintham, E.O.; Yoshida, T.; Smith, S.; Pavlou, H.J.; Goodwin, S.F.; Carazo, P.; Wigby, S. Interactions between the sexual identity of the nervous system and the social environment mediate lifespan in Drosophila melanogaster. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C. Lekking behavior of the black soldier fly (Diptera: Stratiomyidae). Florida Entomol. 2001, 84, 729. [Google Scholar] [CrossRef]
- Jones, T.M.; Hamilton, J.G.C. A role for pheromones in mate choice in a lekking sandfly. Anim. Behav. 1998, 56, 891–898. [Google Scholar] [CrossRef]
- Shelly, T.; Edu, J.; Pahio, E. Mate choice by lekking males: Evidence from the Mediterranean fruit fly from field cage trials (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2012, 105, 368–376. [Google Scholar] [CrossRef]
- Mir, S.H.; Mir, G.M. Lekking behaviour and male-male rivalry in the melon fly Bactrocera cucurbitae (Coquillett) (Diptera: Tephritidae). J. Insect Behav. 2016, 29, 379–384. [Google Scholar] [CrossRef]
- Alcock, J. A large male competitive advantage in a lekking fly, Hermetia comstockiwilliston (Diptera: Stratiomyidae). Psyche 1990, 97, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Jarrige, A.; Riemann, D.; Goubault, M.; Schmoll, T. Strategic sperm allocation in response to perceived sperm competition risk in a lekking insect. Anim. Behav. 2015, 109, 81–87. [Google Scholar] [CrossRef]
- Mayo, O. A century of Hardy-Weinberg equilibrium. Twin Res. Hum. Genet. 2008, 11, 249–256. [Google Scholar] [CrossRef]
- Baniel, A. Assortative Mating. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T.K., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2018; ISBN 9783319478296. [Google Scholar]
- Hedrick, P.W.; Tuttle, E.M.; Gonser, R.A. Negative-assortative mating in the white-throated sparrow. J. Hered. 2018, 109, 223–231. [Google Scholar] [CrossRef]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. Selected life-history traits of black soldier flies (Diptera: Stratiomyidae) reared on three artificial diets. Ann. Entomol. Soc. Am. 2002, 95, 379–386. [Google Scholar] [CrossRef]
- Oliveira, F.; Doelle, K.; Smith, R. External morphology of Hermetia illucens Stratiomyidae: Diptera (L.1758) based on electron microscopy. Annu. Res. Rev. Biol. 2016, 9, 1–10. [Google Scholar] [CrossRef]
- Nakamura, S.; Ichiki, R.T.; Shimoda, M.; Morioka, S. Small-scale rearing of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae), in the laboratory: Low-cost and year-round rearing. Appl. Entomol. Zool. 2016, 51, 161–166. [Google Scholar] [CrossRef]
- Bertinetti, C.; Samayoa, A.C.; Hwang, S.Y. Effects of feeding adults of Hermetia illucens (Diptera: Stratiomyidae) on longevity, oviposition, and egg hatchability: Insights into optimizing egg production. J. Insect Sci. 2019, 19, 1–7. [Google Scholar] [CrossRef]
- Bruno, D.; Bonelli, M.; Cadamuro, A.G.; Reguzzoni, M.; Grimaldi, A.; Casartelli, M.; Tettamanti, G. The digestive system of the adult Hermetia illucens (Diptera: Stratiomyidae): Morphological features and functional properties. Cell Tissue Res. 2019, 378, 221–238. [Google Scholar] [CrossRef]
- Samayoa, A.C.; Chen, W.T.; Hwang, S.Y. Survival and development of Hermetia illucens (Diptera: Stratiomyidae): A biodegradation agent of organic waste. J. Econ. Entomol. 2016, 109, 2580–2585. [Google Scholar] [CrossRef]
- Saghai-Maroof, M.A.; Soliman, K.M.; Jorgensen, R.A.; Allard, R.W. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 1984, 81, 8014–8018. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, X. Comparison of methods for DNA extraction from a single chironomid for PCR analysis. Pak. J. Zool. 2012, 44, 421–426. [Google Scholar]
- Park, S.D.E. Microsatellite Toolkit 2001. Available online: http://animalgenomics.ucd.ie/sdepark/ms-toolkit (accessed on 28 July 2017).
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Brookfield, J.F.Y. A simple new method for estimating null allele frequency from heterozygote deficiency. Mol. Ecol. Notes 1996, 5, 453–455. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Kalinowski, S.T. HP-RARE 1.0: A computer program for performing rarefaction on measures of allelic richness. Mol. Ecol. Notes 2005, 5, 187–189. [Google Scholar] [CrossRef]
- Addinsoft XLSTAT Statistical and Data Analysis Solution 2021. Available online: https://xlstat.com (accessed on 5 January 2021).
- Queller, D.C.; Goodnight, K.F. Estimating relatedness using genetic markers. Evolution 1989, 43, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator v2: Re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour. 2014, 14, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Piry, S.; Luikart, G.; Cornuet, J.M. BOTTLENECK: A computer program for detecting recent reduction in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Vandeputte, M.; Mauger, S.; Dupont-Nivet, M. An evaluation of allowing for mismatches as a way to manage genotyping errors in parentage assignment by exclusion. Mol. Ecol. Notes 2006, 6, 265–267. [Google Scholar] [CrossRef]
- Jones, O.R.; Wang, J. COLONY: A program for parentage and sibship inference from multilocus genotype data. Mol. Ecol. Resour. 2010, 10, 551–555. [Google Scholar] [CrossRef]
- Wahlund, S. Zusammensetzung von populationen und korrelationserscheinungen vom standpunkt der vererbungslehre aus betrachtet. Hereditas 1928, 11, 65–106. [Google Scholar] [CrossRef]
- Park, S.; Choi, H.; Choi, J.; Jeong, G. Population structure of the exotic black soldier fly, Hermetia illucens (Diptera: Stratiomyidae) in Korea. Korean J. Environ. Ecol. 2017, 31, 520–528. [Google Scholar] [CrossRef]
- Vandewoestijne, S.; Nève, G.; Baguette, M. Spatial and temporal population genetic structure of the butterfly Aglais urticae L. (Lepidoptera, Nymphalidae). Mol. Ecol. 1999, 8, 1539–1543. [Google Scholar] [CrossRef]
- Okeyo, W.A.; Saarman, N.P.; Mengual, M.; Dion, K.; Bateta, R.; Mireji, P.O.; Okoth, S.; Ouma, J.O.; Ouma, C.; Ochieng, J.; et al. Temporal genetic differentiation in Glossina pallidipes tsetse fly populations in Kenya. Parasit. Vectors 2017, 10, 1–13. [Google Scholar] [CrossRef]
- Nei, M. Definition and estimation of fixation indices. Evolution 1986, 40, 643–645. [Google Scholar] [CrossRef]
- Neel, M.C.; McKelvey, K.; Ryman, N.; Lloyd, M.W.; Short Bull, R.; Allendorf, F.W.; Schwartz, M.K.; Waples, R.S. Estimation of effective population size in continuously distributed populations: There goes the neighborhood. Heredity 2013, 111, 189–199. [Google Scholar] [CrossRef]
- Whitlock, M.C. Fixation of new alleles and the extinction of small populations: Drift load, beneficial alleles, and sexual selection. Evolution 2000, 54, 1855–1861. [Google Scholar] [CrossRef]
- Waples, R.S.; Antao, T.; Luikart, G. Effects of overlapping generations on linkage disequilibrium estimates of effective population size. Genetics 2014, 197, 769–780. [Google Scholar] [CrossRef]
- Wang, J.; Santiago, E.; Caballero, A. Prediction and estimation of effective population size. Heredity 2016, 117, 193–206. [Google Scholar] [CrossRef]
- Sánchez-Montes, G.; Ariño, A.H.; Vizmanos, J.L.; Wang, J.; Martínez-Solano, Í. Effects of sample size and full sibs on genetic diversity characterization: A case study of three syntopic Iberian pond-breeding amphibians. J. Hered. 2017, 108, 535–543. [Google Scholar] [CrossRef]
- Gilchrist, A.S.; Cameron, E.C.; Sved, J.A.; Meats, A.W. Genetic consequences of domestication and mass rearing of pest fruit fly Bactrocera tryoni (Diptera: Tephritidae). J. Econ. Entomol. 2012, 105, 1051–1056. [Google Scholar] [CrossRef]
- Francuski, L.; Djurakic, M.; Ludoški, J.; Hurtado, P.; Pérez-Bañón, C.; Ståhls, G.; Rojo, S.; Milankov, V. Shift in phenotypic variation coupled with rapid loss of genetic diversity in captive populations of Eristalis tenax (Diptera: Syrphidae): Consequences for rearing and potential commercial use. J. Econ. Entomol. 2014, 107, 821–832. [Google Scholar] [CrossRef]
- Taylor, H.R. The use and abuse of genetic marker-based estimates of relatedness and inbreeding. Ecol. Evol. 2015, 5, 3140–3150. [Google Scholar] [CrossRef]
- Harris, A.M.; DeGiorgio, M. An unbiased estimator of gene diversity with improved variance for samples containing related and inbred individuals of any ploidy. G3 Genes Genomes Genet. 2017, 7, 671–691. [Google Scholar] [CrossRef]
- Sheppard, D.C.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. Rearing methods for the black soldier fly (Diptera: Stratiomyidae). J. Med. Entomol. 2002, 39, 695–698. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; Sheppard, D.C. Factors influencing mating and oviposition of black soldier flies (Diptera: Stratiomyidae) in a colony. J. Entomol. Sci. 2002, 37, 345–352. [Google Scholar] [CrossRef]
- Dortmans, B.M.A.; Diener, S.; Verstappen, B.M.; Zurbrügg, C. Black Soldier Fly Biowaste Processing—A Step-by-Step Guide; Donahue, P., Ed.; Eawag-Swiss Federal Institute of Aquatic Science and Technology: Dübendorf, Switzerland, 2017; ISBN 9783906484662.
- Frankham, R.; Bradshaw, C.J.A.; Brook, B.W. Genetics in conservation management: Revised recommendations for the 50/500 rules, Red List criteria and population viability analyses. Biol. Conserv. 2014, 170, 56–63. [Google Scholar] [CrossRef]
- Barmentlo, S.H.; Meirmans, P.G.; Luijten, S.H.; Triest, L.; Oostermeijer, J.G.B. Outbreeding depression and breeding system evolution in small, remnant populations of Primula vulgaris: Consequences for genetic rescue. Conserv. Genet. 2018, 19, 545–554. [Google Scholar] [CrossRef]
- Lack, J.B.; Monette, M.J.; Johanning, E.J.; Sprengelmeyer, Q.D.; Pool, J.E. Decanalization of wing development accompanied the evolution of large wings in high-altitude Drosophila. Proc. Natl. Acad. Sci. USA 2016, 113, 1014–1019. [Google Scholar] [CrossRef]
- Slatkin, M. Linkage disequilibrium—Understanding the evolutionary past and mapping the medical future. Nat. Rev. Genet. 2008, 9, 477–485. [Google Scholar] [CrossRef]
- Engqvist, L.; Cordes, N.; Schwenniger, J.; Bakhtina, S.; Schmoll, T. Female remating behavior in a lekking moth. Ethology 2014, 120, 662–671. [Google Scholar] [CrossRef]
- Shelly, T.E. Sexual selection on leks: A fruit fly primer. J. Insect Sci. 2018, 18, 1–16. [Google Scholar] [CrossRef]
- Le Page, S.; Sepil, I.; Flintham, E.; Pizzari, T.; Carazo, P.; Wigby, S. Male relatedness and familiarity are required to modulate male-induced harm to females in Drosophila. Proc. R. Soc. B Biol. Sci. 2017, 284, 11–14. [Google Scholar] [CrossRef] [PubMed]
- Kvarnemo, C.; Simmons, L.W. Polyandry as a mediator of sexual selection before and after mating. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Wild2015 | Wild2018 | F28 | F48 | F52 | F62 |
|---|---|---|---|---|---|---|
| Sample size (n) | 30 | 30 | 30 | 30 | 30 | 29 |
| Ne | 22.1 | 30.0 | 59.0 | 56.7 | 22.6 | 24.4 |
| (14.7–36.6) | (16.4–82.4) | (21.7–∞) | (14.9–∞) | (5.3–∞) | (7.4–∞) | |
| Wilcoxon test | ||||||
| IAM | ||||||
| HE excess | 0.230 | 0.004 ** | 0.680 | 0.727 | 0.148 | 0.039 * |
| HE deficiency | 0.809 | 0.998 | 0.371 | 0.320 | 0.945 | 0.973 |
| SMM | ||||||
| HE excess | 0.902 | 0.986 | 1.000 | 0.986 | 0.852 | 0.711 |
| HE deficiency | 0.125 | 0.020 * | 0.002 ** | 0.020 * | 0.188 | 0.344 |
| TPM | ||||||
| HE excess | 0.727 | 0.371 | 0.990 | 0.973 | 0.406 | 0.289 |
| HE deficiency | 0.320 | 0.680 | 0.014 * | 0.037 * | 0.656 | 0.766 |
| Wild2015 | Wild2018 | F28 | F48 | F52 | |
|---|---|---|---|---|---|
| Wild2015 | |||||
| Wild2018 | 0.062 ** | ||||
| F28 | 0.094 ** | 0.162 ** | |||
| F48 | 0.139 ** | 0.225 ** | 0.112 ** | ||
| F52 | 0.096 ** | 0.172 ** | 0.033 ** | 0.047 ** | |
| F62 | 0.160 ** | 0.163 ** | 0.082 ** | 0.201 ** | 0.103 ** |
| Source of Variation | Variation (%) | Fixation Index |
|---|---|---|
| Amongst groups | 6.87 | FST = 0.150 ** |
| Amongst generations within groups | 8.10 | FSC = 0.087 ** |
| Within groups | 85.03 | FCT = 0.069 * |
| Population | Mean Pairwise Relatedness |
| Source population (S) | −0.034 (−0.094–0.126) |
| Candidate parents (F0) | 0.093 (−0.421–0.347) |
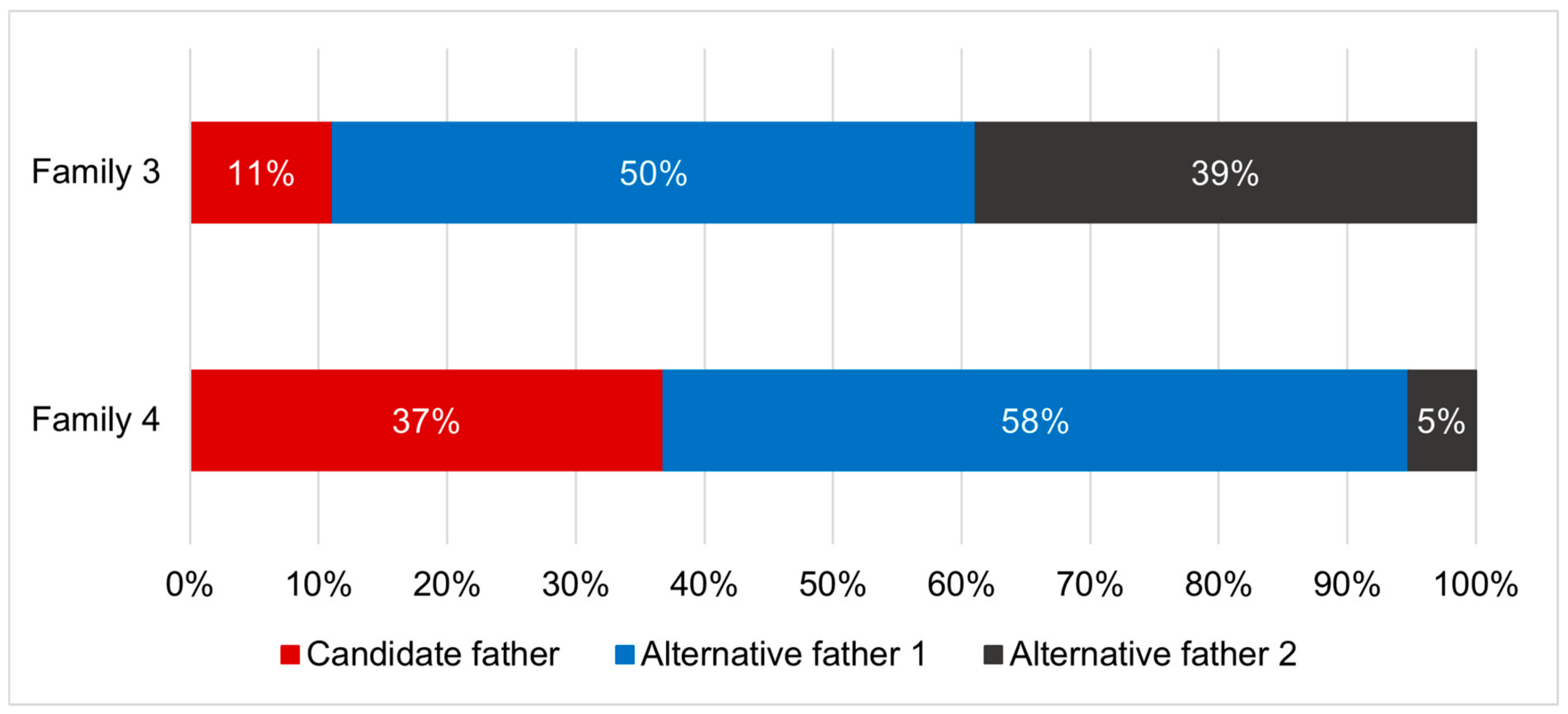
| Parent Pair | Pairwise Relatedness |
| Family 1 | 0.455 |
| Family 2 | 0.455 |
| Family 3 | 1.000 |
| Family 4 | −0.430 |
| Family 5 | 0.700 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoffmann, L.; Hull, K.L.; Bierman, A.; Badenhorst, R.; Bester-van der Merwe, A.E.; Rhode, C. Patterns of Genetic Diversity and Mating Systems in a Mass-Reared Black Soldier Fly Colony. Insects 2021, 12, 480. https://doi.org/10.3390/insects12060480
Hoffmann L, Hull KL, Bierman A, Badenhorst R, Bester-van der Merwe AE, Rhode C. Patterns of Genetic Diversity and Mating Systems in a Mass-Reared Black Soldier Fly Colony. Insects. 2021; 12(6):480. https://doi.org/10.3390/insects12060480
Chicago/Turabian StyleHoffmann, Lelanie, Kelvin L. Hull, Anandi Bierman, Rozane Badenhorst, Aletta E. Bester-van der Merwe, and Clint Rhode. 2021. "Patterns of Genetic Diversity and Mating Systems in a Mass-Reared Black Soldier Fly Colony" Insects 12, no. 6: 480. https://doi.org/10.3390/insects12060480
APA StyleHoffmann, L., Hull, K. L., Bierman, A., Badenhorst, R., Bester-van der Merwe, A. E., & Rhode, C. (2021). Patterns of Genetic Diversity and Mating Systems in a Mass-Reared Black Soldier Fly Colony. Insects, 12(6), 480. https://doi.org/10.3390/insects12060480