
Nootkatone Is an Effective Repellent against Aedes aegypti and Aedes albopictus

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Colonies
2.2. Zika Virus Infection of Mosquitoes
2.3. Bottle Bioassay: Mosquito Kill Assay
2.4. RIBB Assay
3. Results
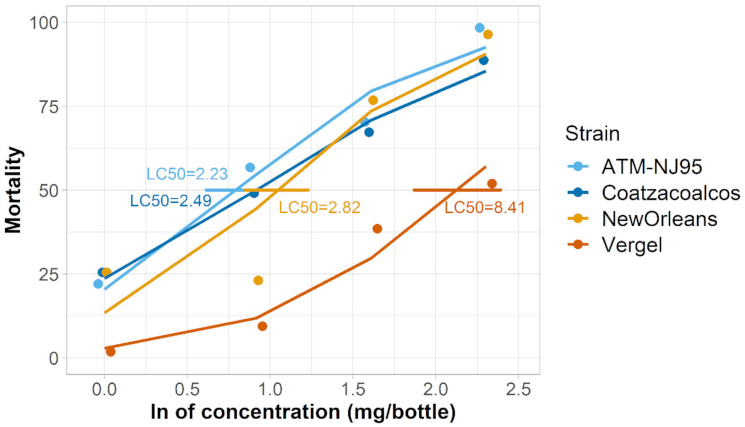
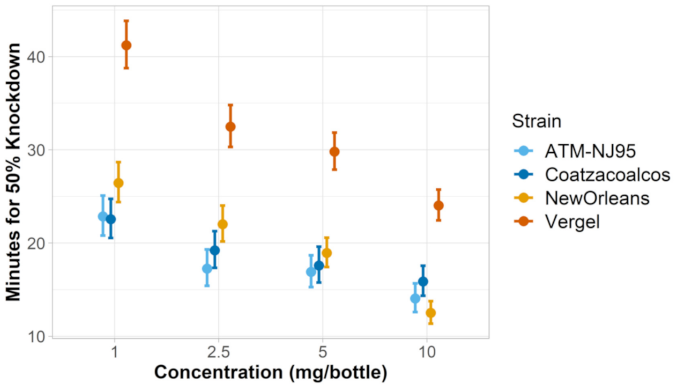
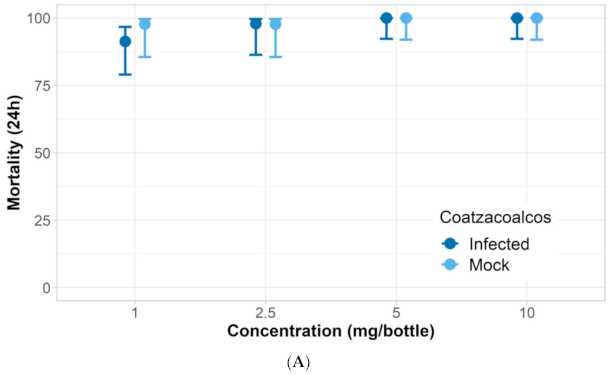
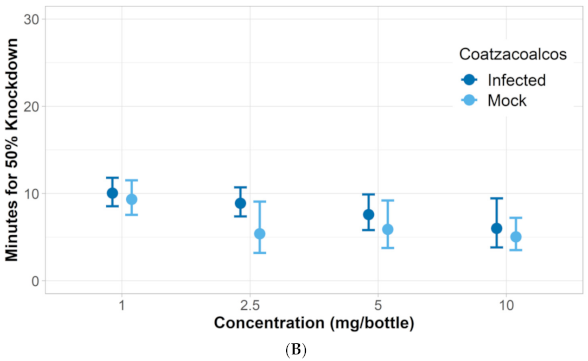
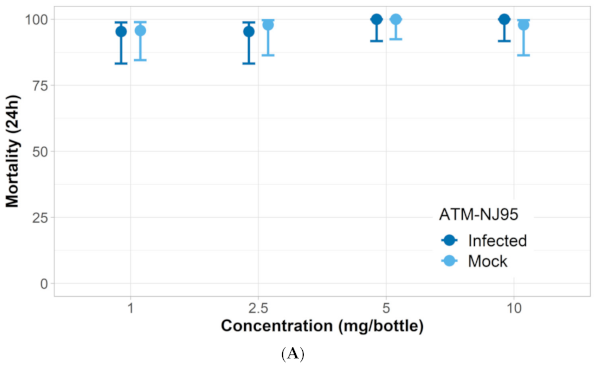
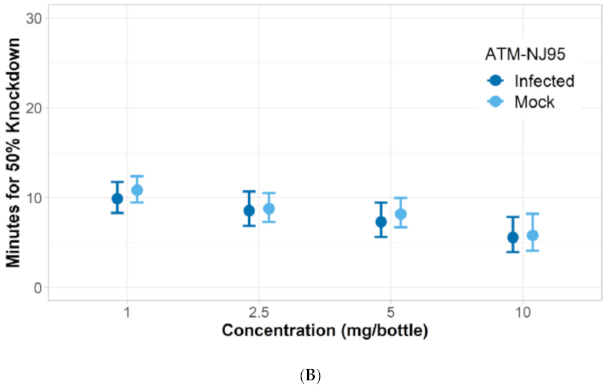
3.1. Verification of Permethrin Resistance
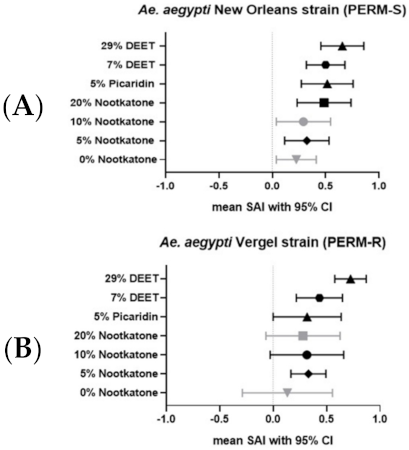
3.2. RIBB Analysis of Nootkatone as a Personal Repellent for A. aegypti Mosquitoes
4. Discussion
4.1. Nootkatone and Insecticide Bottle Bioassays
4.2. Nootkatone and Repellency Assays
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petersen, L.R.; Jamieson, D.J.; Powers, A.M.; Honein, M.A. Zika Virus. N. Engl. J. Med. 2016, 374, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Kindhauser, M.K.; Allen, T.; Frank, V.; Santhana, R.S.; Dye, C. Zika: The origin and spread of a mosquito-borne virus. Bull. World Heal. Organ. 2016, 94, 675–686C. [Google Scholar] [CrossRef] [PubMed]
- Vanlerberghe, V.; Toledo, M.E.; Rodriguez, M.; Gomez, D.; Baly, A.; Benitez, J.R.; Van Der Stuyft, P. Community involvement in dengue vector control: Cluster randomised trial. BMJ 2009, 338, b1959. [Google Scholar] [CrossRef]
- Brengues, C.; Hawkes, N.J.; Chandre, F.; McCarroll, L.; Duchon, S.; Guillet, P.; Manguin, S.; Morgan, J.C.; Hemingway, J. Pyrethroid and DDT cross-resistance in Aedes aegypti is correlated with novel mutations in the voltage-gated sodium channel gene. Med Veter. Èntomol. 2003, 17, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Harris, A.F.; Ranson, H.; Rajatileka, S. Pyrethroid Resistance in Aedes aegypti from Grand Cayman. Am. J. Trop. Med. Hyg. 2010, 83, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Saavedra-Rodriguez, K.; Urdaneta-Marquez, L.; Rajatileka, S.; Moulton, M.; Flores, A.E.; Fernandez-Salas, I.; Bisset, J.; Rodriguez, M.; McCall, P.J.; Donnelly, M.J.; et al. A mutation in the voltage-gated sodium channel gene associated with pyrethroid resistance in Latin American Aedes aegypti. Insect Mol. Biol. 2007, 16, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Pichler, V.; Bellini, R.; Veronesi, R.; Arnoldi, D.; Rizzoli, A.; Lia, R.P.; Otranto, D.; Montarsi, F.; Carlin, S.; Ballardini, M.; et al. First evidence of resistance to pyrethroid insecticides in Italian Aedes albopictus populations 26 years after invasion. Pest Manag. Sci. 2018, 74, 1319–1327. [Google Scholar] [CrossRef] [PubMed]
- Flores, A.E.; Ponce, G.; Silva, B.G.; Gutierrez, S.M.; Bobadilla, C.; Lopez, B.; Mercado, R.; Black, W.C. Wide Spread Cross Resistance to Pyrethroids in Aedes aegypti (Diptera: Culicidae) From Veracruz State Mexico. J. Econ. Èntomol. 2013, 106, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Panella, N.A.; Dolan, M.C.; Karchesy, J.J.; Xiong, Y.; Peralta-Cruz, J.; Khasawneh, M.; Montenieri, J.A.; Maupin, G.O. Use of novel compounds for pest control: Insecticidal and acaricidal activity of essential oil components from heartwood of Alaska yellow cedar. J. Med. Entomol. 2005, 42, 352–358. [Google Scholar] [CrossRef]
- U.S. Food & Drug Administration’s Code of Federal Regulations. 21CFR172.515. 2019. Available online: https://www.accessdata.fda.gov/scripts/cdrh/cfdocs/cfcfr/cfrsearch.cfm?fr=172.515 (accessed on 28 October 2020).
- MacLeod, W.D.; Buigues, N.M. Sesquiterpenes. I. Nootkatone, A New Grapefruit Flavor Constituent. J. Food Sci. 1964, 29, 565–568. [Google Scholar] [CrossRef]
- Robbins, P.J.; Cherniack, M.G. Review of the biodistribution and toxicity of the insect repellent N,N-diethyl-m-toluamide (DEET). J. Toxicol. Environ. Health 1986, 18, 503–525. [Google Scholar] [CrossRef]
- Maia, M.F.; Moore, S.J. Plant-based insect repellents: A review of their efficacy, development and testing. Malar. J. 2011, 10 (Suppl. S1), S11. [Google Scholar] [CrossRef]
- Zhu, B.C.R.; Henderson, G.; Chen, F.; Maistrello, L.; Laine, R.A. Nootkatone is a repellent for Formosan subterranean termite (Coptotermes formosanus). J. Chem. Ecol. 2001, 27, 523–531. [Google Scholar] [CrossRef]
- Vera-Maloof, F.Z.; Saavedra-Rodriguez, K.; Elizondo-Quiroga, A.E.; Lozano-Fuentes, S.; Iv, W.C.B. Coevolution of the Ile1,016 and Cys1,534 Mutations in the Voltage Gated Sodium Channel Gene of Aedes aegypti in Mexico. PLoS Neglected Trop. Dis. 2015, 9, e0004263. [Google Scholar] [CrossRef]
- Denham, S.; Eisen, L.; Beaty, M.; Beaty, B.J.; Black, W.C.; Saavedra-Rodriguez, K. Two Novel Bioassays to Assess the Effects of Pyrethroid-Treated Netting on Knockdown-Susceptible Versus Resistant Strains of Aedes aegypti. J. Am. Mosq. Control. Assoc. 2015, 31, 52–62. [Google Scholar] [CrossRef]
- Campbell, C.L.; Saavedra-Rodriguez, K.; Kubik, T.D.; Lenhart, A.; Lozano-Fuentes, S.; Black, W.C. Vgsc-interacting proteins are genetically associated with pyrethroid resistance in Aedes aegypti. PLoS ONE 2019, 14, e0211497. [Google Scholar] [CrossRef]
- Marcombe, S.; Farajollahi, A.; Healy, S.P.; Clark, G.G.; Fonseca, D.M. Insecticide Resistance Status of United States Populations of Aedes albopictus and Mechanisms Involved. PLoS ONE 2014, 9, e1019922. [Google Scholar] [CrossRef]
- WHO Pesticide Evaluation Scheme (WHOPES) (Ed.) Guidelines for Efficacy Testing of Insecticides for Indoor and Outdoor Ground-Applied Space Spray Applications; World Health Organization: Geneva, Switzerland, 2009; p. 83. [Google Scholar]
- WHO Pesticide Evaluation Scheme (WHOPES) (Ed.) Guidelines for Efficacy Testing of Mosquito Repellents for Human Skin; WHO Press: Geneva, Switzerland, 2009; p. 30. [Google Scholar]
- WHO Pesticide Evaluation Scheme (WHOPES) (Ed.) Guidelines for Efficacy Testing of Spatial Repellents, Control of Neglected Tropical Diseases; WHO Press: Geneva, Switzerland, 2013; pp. 1–48. [Google Scholar]
- Wagman, J.M.; Achee, N.L.; Grieco, J.P. Insensitivity to the Spatial Repellent Action of Transfluthrin in Aedes aegypti: A Heritable Trait Associated with Decreased Insecticide Susceptibility. PLoS Neglected Trop. Dis. 2015, 9, e0003726. [Google Scholar] [CrossRef]
- Saavedra-Rodriguez, K.; Ponce-Garcia, G.; Loroño-Pino, M.A.; García-Rejón, J.E.; Beaty, M.; Eisen, L.; Flores-Suarez, A.; Reyes-Solis, G.; Lozano-Fuentes, S.; Black, W.C.; et al. Local Evolution of Pyrethroid Resistance Offsets Gene Flow Among Aedes aegypti Collections in Yucatan State, Mexico. Am. J. Trop. Med. Hyg. 2015, 92, 201–209. [Google Scholar] [CrossRef]
- McAllister, J.C.; Adams, M.F. Mode of action for natural products isolated from essential oils of two trees is different from available mosquito adulticides. J. Med. Èntomol. 2010, 47, 1123–1126. [Google Scholar] [CrossRef]
- Hemingway, J.; Ranson, H. Insecticide Resistance in Insect Vectors of Human Disease. Annu. Rev. Èntomol. 2000, 45, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.T.; Smallegange, R.C.; Van Loon, J.J.A.; Ter Braak, C.J.F.; Takken, W. Interindividual variation in the attractiveness of human odours to the malaria mosquito Anopheles gambiae s. s. Med. Veter. Èntomol. 2006, 20, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Kuri-Morales, P.A.; Correa-Morales, F.; González-Acosta, C.; Sánchez-Tejeda, G.; Moreno-Garcia, M.; Dávalos-Becerril, E.; Juárez-Franco, M.F.; Benitez-Alva, J.I.; González-Roldán, J.F. Repellency of 29 Synthetic and Natural Commercial Topical Insect Repellents Against Aedes aegypti (Diptera: Culicidae) in Central Mexico. J. Med. Èntomol. 2017, 54, 1305–1311. [Google Scholar] [CrossRef] [PubMed]
- Flor-Weiler, L.B.; Behle, R.W.; Stafford, K.C., 3rd. Susceptibility of four tick species, Amblyomma americanum, Dermacentor variabilis, Ixodes scapularis, and Rhipicephalus sanguineus (Acari: Ixodidae), to nootkatone from essential oil of grapefruit. J. Med. Entomol. 2011, 48, 322–326. [Google Scholar] [CrossRef]
- Dolan, M.C.; Jordan, R.A.; Schulze, T.L.; Schulze, C.J.; Manning, M.C.; Ruffolo, D.; Schmidt, J.P.; Piesman, J.; Karchesy, J.J. Ability of Two Natural Products, Nootkatone and Carvacrol, to Suppress Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae) in a Lyme Disease Endemic Area of New Jersey. J. Econ. Èntomol. 2009, 102, 2316–2324. [Google Scholar] [CrossRef]
- Li, M.; Chapman, G.B. Why Do People Like Natural? Instrumental and Ideational Bases for the Naturalness Preference. J. Appl. Soc. Psychol. 2012, 42, 2859–2878. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mosquito | Strain | Presumed | Actual | Reference |
|---|---|---|---|---|
| Species | - | PERM-status * | PERM-status ** | - |
| Aedes aegypti | New Orleans | Susceptible | Susceptible | [5,6] |
| Aedes aegypti | Vergel | Resistant | Resistant | [15,16] |
| Aedes albopictus | ATM-NJ95 | Susceptible | Susceptible | [18] |
| Aedes albopictus | Coatzacoalcos | Unknown | Susceptible | - |
| Species and Strain Name | LC50 (mg/Bottle) | 95% CI (mg/Bottle) | Intercept | Slope | GOF | p Value | n |
|---|---|---|---|---|---|---|---|
| A. albopictus ATM-NJ95 | 2.24 | 1.85–2.71 | −1.36 | 1.69 | 3.56 | 0.130 | 252 |
| A. albopictus Coatzacoalcos | 2.50 | 1.95–3.20 | −1.17 | 1.28 | 0.48 | 0.850 | 217 |
| A. aegypti New Orleans | 2.83 | 2.34–3.42 | −1.87 | 1.79 | 13.38 | 0.00 | 214 |
| A. aegypti Vergel | 8.42 | 6.49–10.91 | −3.52 | 1.65 | 2.44 | 0.296 | 213 |
| Strain Name | LC50 (mg/Bottle) | 95% Confidence Interval (mg/Bottle) |
|---|---|---|
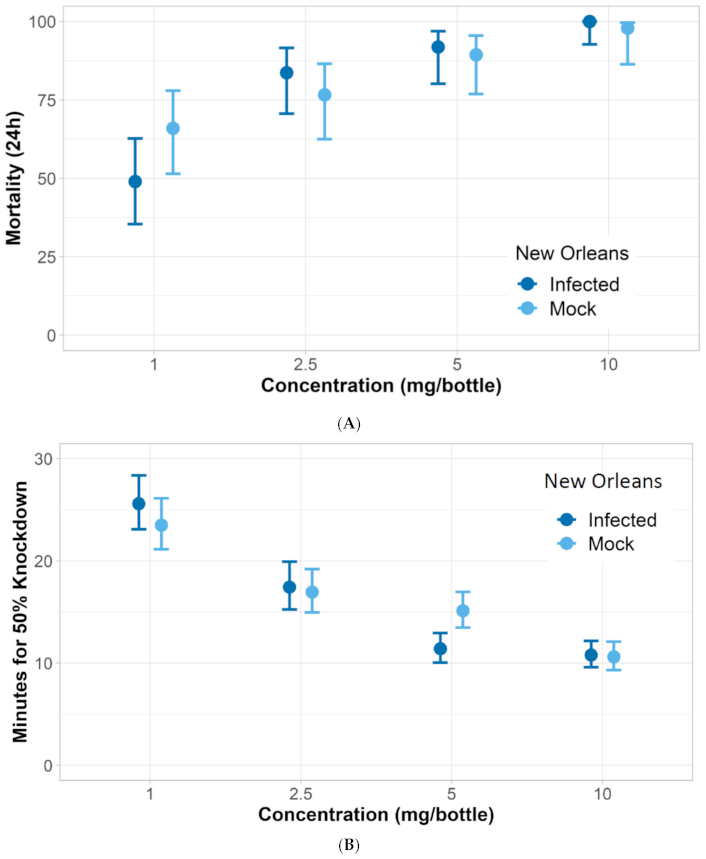
| A. aegypti New Orleans infected | 1.20 | 1.07–1.33 |
| A. aegypti New Orleans mock | 0.97 | 0.81–1.12 |
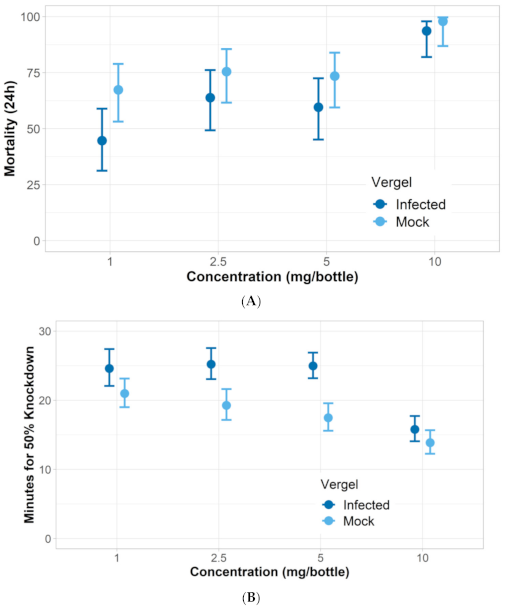
| A. aegypti Vergel infected | 1.87 | 1.68–2.06 |
| A. aegypti Vergel mock | 1.21 | 1.04–1.38 |
| A. albopictus Coatzacoalcos infected | 0.43 | 0.31–0.55 |
| A. albopictus Coatzacoalcos mock | 0.31 | 0.21–0.42 |
| A. albopictus ATM-NJ95 infected | 0.34 | 0.23–0.45 |
| A. albopictus ATM-NJ95 mock | 0.43 | 0.32–0.54 |
| A. aegypti New Orleans Strain (PERM-S) | A. aegypti Vergel Strain (PERM-R) | |||||||
|---|---|---|---|---|---|---|---|---|
| Repellent Formulation | Mean Proportion BF on Untreated Arm (95% CI) | Mean Proportion BF on Treated Arm (95% CI) | Difference in Means (95% CI) | % Reduction | Mean Proportion BF on Untreated Arm (95% CI) | Mean Proportion BF on Treated Arm (95% CI) | Difference in Means (95% CI) | % Reduction |
| 0% Nootkatone | 0.90 (0.84, 0.97) | 0.97 ˄,↑ (0.93, 1.02) | −0.03 (−0.09, −0.04) | −8% | 0.92 (0.83, 1.00) | 0.97 ˄ (0.93, 1.01) | −0.05 (−0.10,0.00) | −5% |
| 5% Nootkatone | 0.94 (0.88, 1.00) | 0.94 *,↑ (0.86, 1.03) | 0.00 (0.02, −0.03) | 0% | 0.90 (0.83, 0.96) | 0.65 *,˅↑ (0.41, 0.89) | 0.25 (0.42, 0.08) | 27% |
| 10% Nootkatone | 0.88 (0.69, 1.07) | 0.56 ˅,↑ (0.26, 0.87) | 0.32 (0.43, 0.20) | 36% | 0.89 (0.77, 1.01) | 0.42 ˅ (0.13, 0.70) | 0.47 (0.63, 0.31) | 53% |
| 20% Nootkatone | 0.95 (0.90, 1.00) | 0.72 ↑ (0.53, 0.91) | 0.23 (0.37, 0.09) | 24% | 0.95 (0.90, 1.00) | 0.45 ˅ (0.22, 0.69) | 0.50 (0.68, 0.32) | 53% |
| 5% Picaradin | 0.91 (0.85, 0.97) | 0.63 ↑ (0.41, 0.84) | 0.28 (0.44, 0.13) | 31% | 0.81 (0.58, 1.04) | 0.47 ˅ (0.19, 0.76) | 0.34 (0.39, 0.28) | 42% |
| 7% DEET | 0.94 (0.90, 0.98) | 0.57 ↑ (0.27, 0.88) | 0.37 (0.64, 0.10) | 39% | 0.92 (0.84, 1.01) | 0.49 ˅ (0.23, 0.75) | 0.43 (0.61, 0.25) | 47% |
| 29% DEET | 0.95 (0.91, 0.98) | 0.11 ↓ (−0.09, 0.32) | 0.84 (1.01, 0.67) | 88% | 0.89 (0.78, 1.00) | 0.18 ˅,↓ (−0.03, 0.38) | 0.71 (0.81, 0.62) | 80% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clarkson, T.C.; Janich, A.J.; Sanchez-Vargas, I.; Markle, E.D.; Gray, M.; Foster, J.R.; Black IV, W.C.; Foy, B.D.; Olson, K.E. Nootkatone Is an Effective Repellent against Aedes aegypti and Aedes albopictus. Insects 2021, 12, 386. https://doi.org/10.3390/insects12050386
Clarkson TC, Janich AJ, Sanchez-Vargas I, Markle ED, Gray M, Foster JR, Black IV WC, Foy BD, Olson KE. Nootkatone Is an Effective Repellent against Aedes aegypti and Aedes albopictus. Insects. 2021; 12(5):386. https://doi.org/10.3390/insects12050386
Chicago/Turabian StyleClarkson, Taylor C., Ashley J. Janich, Irma Sanchez-Vargas, Erin D. Markle, Megan Gray, John R. Foster, William C. Black IV, Brian D. Foy, and Ken E. Olson. 2021. "Nootkatone Is an Effective Repellent against Aedes aegypti and Aedes albopictus" Insects 12, no. 5: 386. https://doi.org/10.3390/insects12050386
APA StyleClarkson, T. C., Janich, A. J., Sanchez-Vargas, I., Markle, E. D., Gray, M., Foster, J. R., Black IV, W. C., Foy, B. D., & Olson, K. E. (2021). Nootkatone Is an Effective Repellent against Aedes aegypti and Aedes albopictus. Insects, 12(5), 386. https://doi.org/10.3390/insects12050386