
GC/TOF-MS-Based Metabolomics Reveals Altered Metabolic Profiles in Wood-Feeding Termite Coptotermes formosanus Shiraki Digesting the Weed Mikania micrantha Kunth


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Termite Colony and Dietary Preparation
2.2. Bioassays
2.3. Metabolite Sample Collection and Preparation
2.4. GC/TOF-MS Analysis
2.5. Metabolomic Data Analysis
3. Results
3.1. ML Diets Increase Food Intake While Decreasing Survival Rates
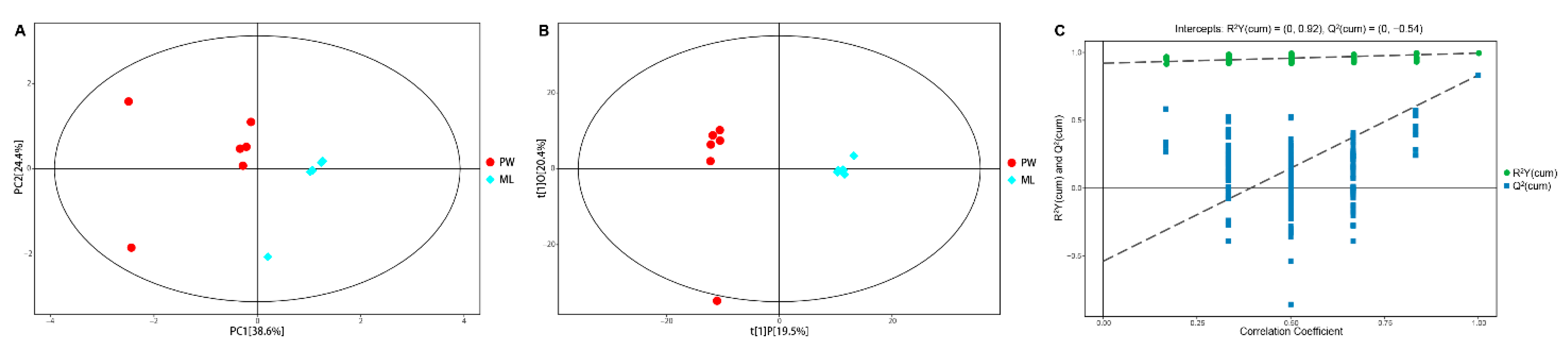
3.2. GC/TOF-MS Analyses Reveal Diet-Specific Metabolic Profiles
3.3. Metabolite Identification and Comparison
3.4. Metabolic Pathway Analyses of Differentially Abundant Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brune, A. Symbiotic digestion of lignocellulose in termite guts. Nat. Rev. Microbiol. 2014, 12, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Eggleton, P.; Tayasu, I. Feeding groups, lifetypes and the global ecology of termites. Ecol. Res. 2001, 16, 941–960. [Google Scholar] [CrossRef]
- Tarmadi, D.; Yoshimura, T.; Tobimatsu, Y.; Yamamura, M.; Umezawa, T. Effects of lignins as diet components on the physiological activities of a lower termite, Coptotermes formosanus Shiraki. J. Insect Physiol. 2017, 103, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Tarmadi, D.; Yoshimura, T.; Tobimatsu, Y.; Yamamura, M.; Miyamoto, T.; Miyagawa, Y.; Umezawa, T. The effects of various lignocelluloses and lignins on physiological responses of a lower termite, Coptotermes formosanus. J. Wood Sci. 2017, 63, 464–472. [Google Scholar] [CrossRef][Green Version]
- Sethi, A.; Slack, J.M.; Kovaleva, E.S.; Buchman, G.W.; Scharf, M.E. Lignin-associated metagene expression in a lignocellulose-digesting termite. Insect Biochem. Mol. Biol. 2013, 43, 91–101. [Google Scholar] [CrossRef]
- Karl, Z.J.; Scharf, M.E. Effects of five diverse lignocellulosic diets on digestive enzyme biochemistry in the termite Reticulitermes flavipes. Arch. Insect Biochem. Physiol. 2015, 90, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Rajarapu, S.P.; Scharf, M.E. Saccharification of Agricultural Lignocellulose Feedstocks and Protein-Level Responses by a Termite Gut-Microbe Bioreactor. Front. Energy Res. 2017, 5, 5. [Google Scholar] [CrossRef]
- Boucias, D.G.; Cai, Y.; Sun, Y.; Lietze, V.-U.; Sen, R.; Raychoudhury, R.; Scharf, M.E. The hindgut lumen prokaryotic microbiota of the termite Reticulitermes flavipes and its responses to dietary lignocellulose composition. Mol. Ecol. 2013, 22, 1836–1853. [Google Scholar] [CrossRef]
- Huang, X.-F.; Bakker, M.G.; Judd, T.M.; Reardon, K.F.; Vivanco, J.M. Variations in Diversity and Richness of Gut Bacterial Communities of Termites (Reticulitermes flavipes) Fed with Grassy and Woody Plant Substrates. Microb. Ecol. 2013, 65, 531–536. [Google Scholar] [CrossRef]
- Duarte, S.; Duarte, M.; Borges, P.A.V.; Nunes, L. Dietary-driven variation effects on the symbiotic flagellate protist communities of the subterranean termite Reticulitermes grassei Clément. J. Appl. Entomol. 2017, 141, 300–307. [Google Scholar] [CrossRef]
- Benjamino, J.; Lincoln, S.; Srivastava, R.; Graf, J. Low-abundant bacteria drive compositional changes in the gut microbiota after dietary alteration. Microbiome 2018, 6, 86. [Google Scholar] [CrossRef]
- Calusinska, M.; Marynowska, M.; Bertucci, M.; Untereiner, B.; Klimek, D.; Goux, X.; Sillam-Dussès, D.; Gawron, P.; Halder, R.; Wilmes, P.; et al. Integrative omics analysis of the termite gut system adaptation to Miscanthus diet identifies lignocellulose degradation enzymes. Commun. Biol. 2020, 3, 275. [Google Scholar] [CrossRef] [PubMed]
- Rajarapu, S.P.; Shreve, J.T.; Bhide, K.P.; Thimmapuram, J.; Scharf, M.E. Metatranscriptomic profiles of Eastern subterranean termites, Reticulitermes flavipes (Kollar) fed on second generation feedstocks. BMC Genom. 2015, 16, 332. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chong, J.; Yamamoto, M.; Xia, J. MetaboAnalystR 2.0: From Raw Spectra to Biological Insights. Metabolites 2019, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Nelson, L.J.; Cool, L.G.; Forschler, B.T.; Haverty, M.I. Correspondence of Soldier Defense Secretion Mixtures with Cuticular Hydrocarbon Phenotypes for Chemotaxonomy of the Termite Genus Reticulitermes in North America. J. Chem. Ecol. 2001, 27, 1449–1479. [Google Scholar] [CrossRef] [PubMed]
- Jirošová, A.; Jančařík, A.; Menezes, R.C.; Bazalová, O.; Dolejšová, K.; Vogel, H.; Jedlička, P.; Buček, A.; Brabcová, J.; Majer, P.; et al. Co-option of the sphingolipid metabolism for the production of nitroalkene defensive chemicals in termite soldiers. Insect Biochem. Mol. Biol. 2017, 82, 52–61. [Google Scholar] [CrossRef]
- Sillam-Dussès, D.; Krasulová, J.; Vrkoslav, V.; Pytelková, J.; Cvačka, J.; Kutalová, K.; Bourguignon, T.; Miura, T.; Šobotník, J. Comparative Study of the Labial Gland Secretion in Termites (Isoptera). PLoS ONE 2012, 7, e46431. [Google Scholar] [CrossRef]
- Geib, S.M.; Filley, T.R.; Hatcher, P.G.; Hoover, K.; Carlson, J.E.; Jimenez-Gasco, M.d.M.; Nakagawa-Izumi, A.; Sleighter, R.L.; Tien, M. Lignin degradation in wood-feeding insects. Proc. Natl. Acad. Sci. USA 2008, 105, 12932. [Google Scholar] [CrossRef] [PubMed]
- Ke, J.; Laskar, D.D.; Chen, S. Tetramethylammonium Hydroxide (TMAH) Thermochemolysis for Probing in Situ Softwood Lignin Modification in Each Gut Segment of the Termite. J. Agric. Food Chem. 2013, 61, 1299–1308. [Google Scholar] [CrossRef]
- Ke, J.; Laskar, D.D.; Singh, D.; Chen, S. In situ lignocellulosic unlocking mechanism for carbohydrate hydrolysis in termites: Crucial lignin modification. Biotechnol. Biofuels 2011, 4, 17. [Google Scholar] [CrossRef]
- Tarmadi, D.; Tobimatsu, Y.; Yamamura, M.; Miyamoto, T.; Miyagawa, Y.; Umezawa, T.; Yoshimura, T. NMR studies on lignocellulose deconstructions in the digestive system of the lower termite Coptotermes formosanus Shiraki. Sci. Rep. 2018, 8, 1290. [Google Scholar] [CrossRef]
- Tokuda, G.; Tsuboi, Y.; Kihara, K.; Saitou, S.; Moriya, S.; Lo, N.; Kikuchi, J. Metabolomic profiling of 13C-labelled cellulose digestion in a lower termite: Insights into gut symbiont function. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140990. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Ye, W.H.; Cao, H.L.; Feng, H.L. Mikania micrantha H. B. K. in China—An overview. Weed Res. 2004, 44, 42–49. [Google Scholar] [CrossRef]
- Borah, A.J.; Singh, S.; Goyal, A.; Moholkar, V.S. An assessment of the potential of invasive weeds as multiple feedstocks for biofuel production. RSC Adv. 2016, 6, 47151–47163. [Google Scholar] [CrossRef]
- Lim, C.J.; Arumugam, M.; Lim, C.K.; Ee, G.C.L. Mercerizing Extraction and Physicochemical Characterizations of Lignocellulosic Fiber from the Leaf Waste of Mikania micrantha Kunth ex H.B.K. J. Nat. Fibers 2020, 17, 726–737. [Google Scholar] [CrossRef]
- Dev, U.K.; Hossain, T.; Islam, Z. PHYTOCHEMICAL INVESTIGATION, ANTIOXIDANT ACTIVITY AND ANTIHELMINTIC ACTIVITY OF MIKANIA MICRANTHA LEAVES. World J. Pharm. Res. 2015, 4, 121–133. [Google Scholar]
- Ishak, A.H.; Shafie, N.H.; Esa, N.M.; Bahari, H.; Ismail, A. From weed to medicinal plant: Antioxidant capacities and phytochemicals of various extracts of Mikania micrantha. Int. J. Agric. Biol. 2018, 20, 561–568. [Google Scholar] [CrossRef]
- Sheam, M.M.; Haque, Z.; Nain, Z. Towards the antimicrobial, therapeutic and invasive properties of Mikania micrantha Knuth: A brief overview. J. Adv. Biotechnol. Exp. Ther. 2020, 3, 92–101. [Google Scholar] [CrossRef]
- Vargo, E.L.; Husseneder, C. Biology of Subterranean Termites: Insights from Molecular Studies of Reticulitermes and Coptotermes. Annu. Rev. Entomol. 2008, 54, 379–403. [Google Scholar] [CrossRef] [PubMed]
- Geng, A.; Cheng, Y.; Wang, Y.; Zhu, D.; Le, Y.; Wu, J.; Xie, R.; Yuan, J.S.; Sun, J. Transcriptome analysis of the digestive system of a wood-feeding termite (Coptotermes formosanus) revealed a unique mechanism for effective biomass degradation. Biotechnol. Biofuels 2018, 11, 24. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Wohlgemuth, G.; Lee, D.Y.; Lu, Y.; Palazoglu, M.; Shahbaz, S.; Fiehn, O. FiehnLib: Mass Spectral and Retention Index Libraries for Metabolomics Based on Quadrupole and Time-of-Flight Gas Chromatography/Mass Spectrometry. Anal. Chem. 2009, 81, 10038–10048. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Zhou, Y.; Qin, D.Q.; Zhang, P.W.; Chen, X.T.; Liu, B.J.; Cheng, D.M.; Zhang, Z.X. Integrated LC–MS and GC–MS-based untargeted metabolomics studies of the effect of azadirachtin on Bactrocera dorsalis larvae. Sci. Rep. 2020, 10, 2306. [Google Scholar] [CrossRef]
- Wang, X.; Yang, B.; Sun, H.; Zhang, A. Pattern Recognition Approaches and Computational Systems Tools for Ultra Performance Liquid Chromatography–Mass Spectrometry-Based Comprehensive Metabolomic Profiling and Pathways Analysis of Biological Data Sets. Anal. Chem. 2012, 84, 428–439. [Google Scholar] [CrossRef]
- Cai, S.; Guo, Y.; Li, Y. Intratree Variation in Viscoelastic Properties of Cell Walls of Masson Pine (Pinus Massoniana Lamb). J. Renew. Mater. 2022, 10, 119–133. [Google Scholar] [CrossRef]
- Pang, X.; Zhang, M.; Hou, Y.; Jiao, Y.; Cen, Y. Evaluation of plant protectants against pest insects. J. Appl. Ecol. 2000, 11, 108–110. [Google Scholar]
- Chou, J.-H.; Sheu, S.-Y.; Lin, K.-Y.; Chen, W.-M.; Arun, A.B.; Young, C.-C. Comamonas odontotermitis sp. nov., isolated from the gut of the termite Odontotermes formosanus. Int. J. Syst. Evol. Microbiol. 2007, 57, 887–891. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wu, J.; Wang, W.; Ding, P.; Feng, L. Proteomics analysis of aromatic catabolic pathways in thermophilic Geobacillus thermodenitrificans NG80-2. J. Proteom. 2012, 75, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, K.; Sharma, D.; Varney, R.; Simmons, B.; Isern, N.; Markillie, L.M.; Nicora, C.; Norbeck, A.; Taylor, R.; Aldrich, J.; et al. Evidence supporting dissimilatory and assimilatory lignin degradation in Enterobacter lignolyticus SCF1. Front. Microbiol. 2013, 4, 280. [Google Scholar] [CrossRef]
- Chen, H.; Si, Y.; Chen, Y.; Yang, H.; Chen, D.; Chen, W. NOx precursors from biomass pyrolysis: Distribution of amino acids in biomass and Tar-N during devolatilization using model compounds. Fuel 2017, 187, 367–375. [Google Scholar] [CrossRef]
- Liu, S.; Hu, C.; Lu, K. Manufacture of Mikania micrantha Vinegar and Investigation of its Repellent Activity for Forcipomyia taiwana. BioResources 2021, 16, 6831–6849. [Google Scholar]
- Mezrich, J.D.; Fechner, J.H.; Zhang, X.; Johnson, B.P.; Burlingham, W.J.; Bradfield, C.A. An interaction between kynurenine and the aryl hydrocarbon receptor can generate regulatory T cells. J. Immunol. 2010, 185, 3190–3198. [Google Scholar] [CrossRef] [PubMed]
- Zhuravlev, A.V.; Vetrovoy, O.V.; Savvateeva-Popova, E.V. Enzymatic and non-enzymatic pathways of kynurenines’ dimerization: The molecular factors for oxidative stress development. PLOS Comput. Biol. 2018, 14, e1006672. [Google Scholar] [CrossRef] [PubMed]
- Agudelo, L.Z.; Femenía, T.; Orhan, F.; Porsmyr-Palmertz, M.; Goiny, M.; Martinez-Redondo, V.; Correia, J.C.; Izadi, M.; Bhat, M.; Schuppe-Koistinen, I.; et al. Skeletal Muscle PGC-1α1 Modulates Kynurenine Metabolism and Mediates Resilience to Stress-Induced Depression. Cell 2014, 159, 33–45. [Google Scholar] [CrossRef]
- Wilinski, D.; Winzeler, J.; Duren, W.; Persons, J.L.; Holme, K.J.; Mosquera, J.; Khabiri, M.; Kinchen, J.M.; Freddolino, P.L.; Karnovsky, A.; et al. Rapid metabolic shifts occur during the transition between hunger and satiety in Drosophila melanogaster. Nat. Commun. 2019, 10, 4052. [Google Scholar] [CrossRef] [PubMed]
- Maffei, M.E. 5-Hydroxytryptophan (5-HTP): Natural Occurrence, Analysis, Biosynthesis, Biotechnology, Physiology and Toxicology. Int. J. Mol. Sci. 2021, 22, 181. [Google Scholar] [CrossRef]
- Neckameyer, W.S. A Trophic Role for Serotonin in the Development of a Simple Feeding Circuit. Dev. Neurosci. 2010, 32, 217–237. [Google Scholar] [CrossRef]
- Neckameyer, W.S.; Coleman, C.M.; Eadie, S.; Goodwin, S.F. Compartmentalization of neuronal and peripheral serotonin synthesis in Drosophila melanogaster. Genes Brain Behav. 2007, 6, 756–769. [Google Scholar] [CrossRef]
- Leandro, J.; Houten, S.M. The lysine degradation pathway: Subcellular compartmentalization and enzyme deficiencies. Mol. Genet. Metab. 2020, 131, 14–22. [Google Scholar] [CrossRef]
- Bao, Y.-Y.; Li, B.-L.; Liu, Z.-B.; Xue, J.; Zhu, Z.-R.; Cheng, J.-A.; Zhang, C.-X. Triazophos up-regulated gene expression in the female brown planthopper, Nilaparvata lugens. J. Insect Physiol. 2010, 56, 1087–1094. [Google Scholar] [CrossRef]
- Wan, P.-J.; Yuan, S.-Y.; Tang, Y.-H.; Li, K.-L.; Yang, L.; Fu, Q.; Li, G.-Q. Pathways of Amino Acid Degradation in Nilaparvata lugens (Stål) with Special Reference to Lysine-Ketoglutarate Reductase/Saccharopine Dehydrogenase (LKR/SDH). PLoS ONE 2015, 10, e0127789. [Google Scholar] [CrossRef] [PubMed]
- Huergo, L.F.; Dixon, R. The Emergence of 2-Oxoglutarate as a Master Regulator Metabolite. Microbiol. Mol. Biol. Rev. 2015, 79, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Gou, L.; Lee, J.; Yang, J.-M.; Park, Y.-D.; Zhou, H.-M.; Zhan, Y.; Lü, Z.-R. The effect of alpha-ketoglutaric acid on tyrosinase activity and conformation: Kinetics and molecular dynamics simulation study. Int. J. Biol. Macromol. 2017, 105, 1654–1662. [Google Scholar] [CrossRef] [PubMed]
- Vavricka, C.J.; Han, Q.; Mehere, P.; Ding, H.; Christensen, B.M.; Li, J. Tyrosine metabolic enzymes from insects and mammals: A comparative perspective. Insect Sci. 2014, 21, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Ipson, B.R.; Fisher, A.L. Roles of the tyrosine isomers meta-tyrosine and ortho-tyrosine in oxidative stress. Ageing Res. Rev. 2016, 27, 93–107. [Google Scholar] [CrossRef]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Barik, S. The Uniqueness of Tryptophan in Biology: Properties, Metabolism, Interactions and Localization in Proteins. Int. J. Mol. Sci. 2020, 21, 8776. [Google Scholar] [CrossRef]
- Oskouian, B.; Saba, J.D. Cancer Treatment Strategies Targeting Sphingolipid Metabolism. In Sphingolipids as Signaling and Regulatory Molecules; Chalfant, C., Poeta, M.D., Eds.; Springer: New York, NY, USA, 2010; pp. 185–205. [Google Scholar]
- Fyrst, H.; Herr, D.R.; Harris, G.L.; Saba, J.D. Characterization of free endogenous C14 and C16 sphingoid bases from Drosophila melanogaster. J. Lipid Res. 2004, 45, 54–62. [Google Scholar] [CrossRef]
- Revelles, O.; Espinosa-Urgel, M.; Molin, S.; Ramos, J.L. The davDT operon of Pseudomonas putida, involved in lysine catabolism, is induced in response to the pathway intermediate delta-aminovaleric acid. J. Bacteriol. 2004, 186, 3439–3446. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Similarity | Mass | VIP | p-Value | Fold Change |
|---|---|---|---|---|---|
| myo-Inositol | 847 | 318 | 2.0983 | 0.0000 | 2.6153 |
| Serine | 843 | 218 | 1.7328 | 0.0056 | 1.9249 |
| Methyl phosphate | 836 | 241 | 1.7052 | 0.0048 | 1.5119 |
| Isomaltose | 832 | 361 | 1.3743 | 0.0425 | 1.8170 |
| d-Arabitol | 823 | 217 | 1.3186 | 0.0317 | 1.5240 |
| Sorbitol | 822 | 307 | 1.3696 | 0.0281 | 1.5178 |
| Phenylalanine | 821 | 218 | 1.3800 | 0.0390 | 1.5033 |
| Ribose | 815 | 103 | 1.5596 | 0.0128 | 1.3338 |
| 3-Hydroxypyridine | 810 | 152 | 1.2158 | 0.0011 | 2.9223 |
| Palmitic acid | 806 | 313 | 1.9743 | 0.0003 | 1.4083 |
| Stearic acid | 788 | 341 | 2.1285 | 0.0000 | 1.7277 |
| 3-Hydroxybutyric acid | 754 | 147 | 1.1377 | 0.0208 | 2.4876 |
| Glutamic acid | 744 | 246 | 1.0792 | 0.0327 | 1.9142 |
| Threitol | 704 | 217 | 1.4558 | 0.0424 | 1.6505 |
| Ribitol | 670 | 217 | 1.5749 | 0.0147 | 5.4272 |
| Pyruvate | 670 | 174 | 1.7218 | 0.0002 | 2.7589 |
| N-Acetyl-d-galactosamine | 667 | 202 | 1.9062 | 0.0006 | 2.8560 |
| Glucose-1-phosphate | 660 | 217 | 1.6018 | 0.0146 | 1.5267 |
| Uridine | 657 | 168 | 1.6539 | 0.0058 | 1.5263 |
| Palatinose | 618 | 204 | 2.1142 | 0.0035 | 12.6492 |
| Ribonic acid, gamma-lactone | 609 | 117 | 1.8789 | 0.0015 | 1.5008 |
| Conduritol b epoxide | 572 | 217 | 1.5895 | 0.0101 | 1.5621 |
| 3-Hydroxy-3-methylglutaric acid | 555 | 247 | 1.3120 | 0.0466 | 1.4388 |
| Methyl heptadecanoate | 543 | 87 | 1.0250 | 0.0000 | 3.9387 |
| 4-Hydroxyphenylacetic acid | 530 | 164 | 1.9489 | 0.0002 | 2.3573 |
| Pelargonic acid | 509 | 215 | 1.9204 | 0.0040 | 5.8735 |
| Linoleic acid methyl ester | 503 | 163 | 1.0131 | 0.0342 | 1.9009 |
| Lactose | 389 | 57 | 1.3997 | 0.0304 | 1.3909 |
| 5-Aminovaleric acid lactam | 384 | 156 | 1.2540 | 0.0020 | 3.9391 |
| Nornicotine | 344 | 232 | 1.6329 | 0.0107 | 1.7724 |
| Melibiose | 333 | 332 | 1.0873 | 0.0471 | 1.8535 |
| 1,3-Diaminopropane | 326 | 292 | 1.1778 | 0.0132 | 4.4758 |
| 3-Hydroxynorvaline | 309 | 144 | 1.7495 | 0.0047 | 1.3766 |
| Dihydrotestosterone | 278 | 254 | 1.1360 | 0.0309 | 2.6046 |
| 3,5-Dihydroxyphenylglycine | 259 | 260 | 1.4267 | 0.0047 | 3.4809 |
| Saccharopine | 211 | 274 | 1.1976 | 0.0058 | 3.0467 |
| 4-Aminophenol | 156 | 240 | 1.2509 | 0.0464 | 3.0828 |
| Metabolites | Similarity | Mass | VIP | p-Value | Fold Change |
|---|---|---|---|---|---|
| Maltose | 818 | 204 | 2.2601 | 3.65 × 10−5 | 9.71 × 10−7 |
| Tyrosine | 814 | 218 | 1.9103 | 6.80 × 10−3 | 1.37 × 10−7 |
| 5-Aminovaleric acid | 695 | 174 | 2.2589 | 3.68 × 10−3 | 1.65 × 10−5 |
| 3-Phosphoglycerate | 662 | 227 | 1.3838 | 2.65 × 10−2 | 6.05 × 10−1 |
| 2-Oxoglutarate | 651 | 198 | 2.2552 | 2.00 × 10−3 | 1.97 × 10−5 |
| Tagatose | 646 | 415 | 1.8586 | 7.86 × 10−4 | 4.84 × 10−1 |
| 6-Phosphogluconic acid | 602 | 318 | 1.5976 | 8.79 × 10−3 | 7.27 × 10−1 |
| Ergosterol | 531 | 50 | 1.8937 | 5.81 × 10−3 | 2.70 × 10−5 |
| 5-Hydroxy-l-tryptophan | 511 | 290 | 1.9015 | 1.29 × 10−2 | 8.17 × 10−5 |
| Levoglucosan | 490 | 204 | 1.5614 | 3.36 × 10−5 | 1.61 × 10−1 |
| dAMP | 457 | 315 | 2.0136 | 4.30 × 10−3 | 3.66 × 10−1 |
| Progesterone | 422 | 119 | 1.5917 | 8.71 × 10−3 | 5.47 × 10−1 |
| Thymine | 403 | 270 | 1.6258 | 9.82 × 10−3 | 6.81 × 10−1 |
| Uridine monophosphate (UMP) | 396 | 211 | 1.1891 | 2.90 × 10−2 | 3.67 × 10−1 |
| Sphinganine | 376 | 217 | 1.0614 | 9.34 × 10−4 | 4.01 × 10−1 |
| 5-Dihydrocortisol | 369 | 370 | 1.5367 | 5.92 × 10−3 | 1.13 × 10−1 |
| Glycocyamine | 343 | 274 | 2.0481 | 1.22 × 10−5 | 4.90 × 10−1 |
| Hydantoin, 5-(4-hydroxybutyl)- 2 | 304 | 357 | 2.0694 | 5.55 × 10−5 | 4.49 × 10−1 |
| l-Kynurenine | 304 | 434 | 1.0513 | 2.04 × 10−2 | 5.13 × 10−1 |
| Dehydroabietic acid | 293 | 252 | 1.3628 | 3.19 × 10−2 | 4.98 × 10−3 |
| N-Acetyltryptophan | 266 | 290 | 1.7577 | 1.63 × 10−3 | 3.39 × 10−1 |
| Cholecalciferol | 254 | 213 | 1.7726 | 5.76 × 10−3 | 3.83 × 10−1 |
| Cyclohexylsulfamic acid | 250 | 303 | 1.5366 | 3.44 × 10−2 | 4.79 × 10−1 |
| 6-Methylmercaptopurine | 216 | 352 | 1.2724 | 4.23 × 10−2 | 7.08 × 10−1 |
| 1-Methyladenosine | 212 | 169 | 1.6781 | 7.04 × 10−3 | 5.32 × 10−1 |
| Pathway | −log10(p) | Impact | SDMs 1 |
|---|---|---|---|
| Glycine, serine and threonine metabolism | 4.2426 | 0.2793 | 1,3-diaminopropane↑, Pyruvate↑, Serine↑, Glycocyamine↓ |
| Tryptophan metabolism | 2.7729 | 0.2645 | 5-Hydroxy-l-tryptophan↓, l-Kynurenine↓ |
| Starch and sucrose metabolism | 5.8532 | 0.2516 | Glucose-1-phosphate↑, Isomaltose↑, Maltose↓ |
| Galactose metabolism | 3.8386 | 0.2514 | Glucose-1-phosphate↑, Lactose↑, Melibiose↑, myo-Inositol↑, N-Acetyl-d-galactosamine↑, Sorbitol↑, Tagatose↓ |
| Sphingolipid metabolism | 3.4365 | 0.2146 | Serine↑, Sphinganine↓ |
| Pyruvate metabolism | 3.7194 | 0.1654 | Pyruvate↑ |
| Pyrimidine metabolism | 2.8521 | 0.1417 | Uridine↑, Thymine↓, UMP↓ |
| Amino sugar and nucleotide sugar metabolism | 1.8373 | 0.1147 | Glucose-1-phosphate↑ |
| Citrate cycle (TCA cycle) | 6.5818 | 0.1049 | Pyruvate↑, 2-Oxoglutarate↓ |
| Glycolysis/Gluconeogenesis | 3.0713 | 0.0991 | Glucose-1-phosphate↑, Pyruvate↑ |
| Lysine degradation | 2.24 | 0.069 | Saccharopine↑, 5-Aminovaleric acid↓ |
| Alanine, aspartate and glutamate metabolism | 6.5818 | 0.0676 | Pyruvate↑, 2-Oxoglutarate↓ |
| Fructose and mannose metabolism | 1.5513 | 0.0459 | Sorbitol↑ |
| Fatty acid biosynthesis | 3.4695 | 0.0168 | Palmitic acid↑ |
| Purine metabolism | 2.9125 | 0.0025 | dAMP↓ |
| Tyrosine metabolism | 5.2416 | 0 | 4-Hydroxyphenylacetic acid↑, Pyruvate↑, Tyrosine↓ |
| Cysteine and methionine metabolism | 4.2426 | 0 | Pyruvate↑, Serine↑ |
| d-Glutamine and d-glutamate metabolism | 3.9902 | 0 | Glutamic acid↑, 2-Oxoglutarate↓ |
| Butanoate metabolism | 3.8176 | 0 | 3-Hydroxybutyric acid↑, 2-Oxoglutarate↓ |
| Fatty acid elongation | 3.4695 | 0 | Palmitic acid↑ |
| Pentose and glucuronate interconversions | 2.2258 | 0 | d-Arabitol↑, Glucose-1-phosphate↑ |
| Pentose phosphate pathway | 1.8912 | 0 | Ribose↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Hou, Y.; Zhang, S.; Chen, Y.; Zeng, W.; Li, Z. GC/TOF-MS-Based Metabolomics Reveals Altered Metabolic Profiles in Wood-Feeding Termite Coptotermes formosanus Shiraki Digesting the Weed Mikania micrantha Kunth. Insects 2021, 12, 927. https://doi.org/10.3390/insects12100927
Wu W, Hou Y, Zhang S, Chen Y, Zeng W, Li Z. GC/TOF-MS-Based Metabolomics Reveals Altered Metabolic Profiles in Wood-Feeding Termite Coptotermes formosanus Shiraki Digesting the Weed Mikania micrantha Kunth. Insects. 2021; 12(10):927. https://doi.org/10.3390/insects12100927
Chicago/Turabian StyleWu, Wenjing, Yahui Hou, Shijun Zhang, Yong Chen, Wenhui Zeng, and Zhiqiang Li. 2021. "GC/TOF-MS-Based Metabolomics Reveals Altered Metabolic Profiles in Wood-Feeding Termite Coptotermes formosanus Shiraki Digesting the Weed Mikania micrantha Kunth" Insects 12, no. 10: 927. https://doi.org/10.3390/insects12100927
APA StyleWu, W., Hou, Y., Zhang, S., Chen, Y., Zeng, W., & Li, Z. (2021). GC/TOF-MS-Based Metabolomics Reveals Altered Metabolic Profiles in Wood-Feeding Termite Coptotermes formosanus Shiraki Digesting the Weed Mikania micrantha Kunth. Insects, 12(10), 927. https://doi.org/10.3390/insects12100927