
Surveillance Studies Reveal Diverse and Potentially Pathogenic-Incriminated Vector Mosquito Species across Major Botswana Touristic Hotspots

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
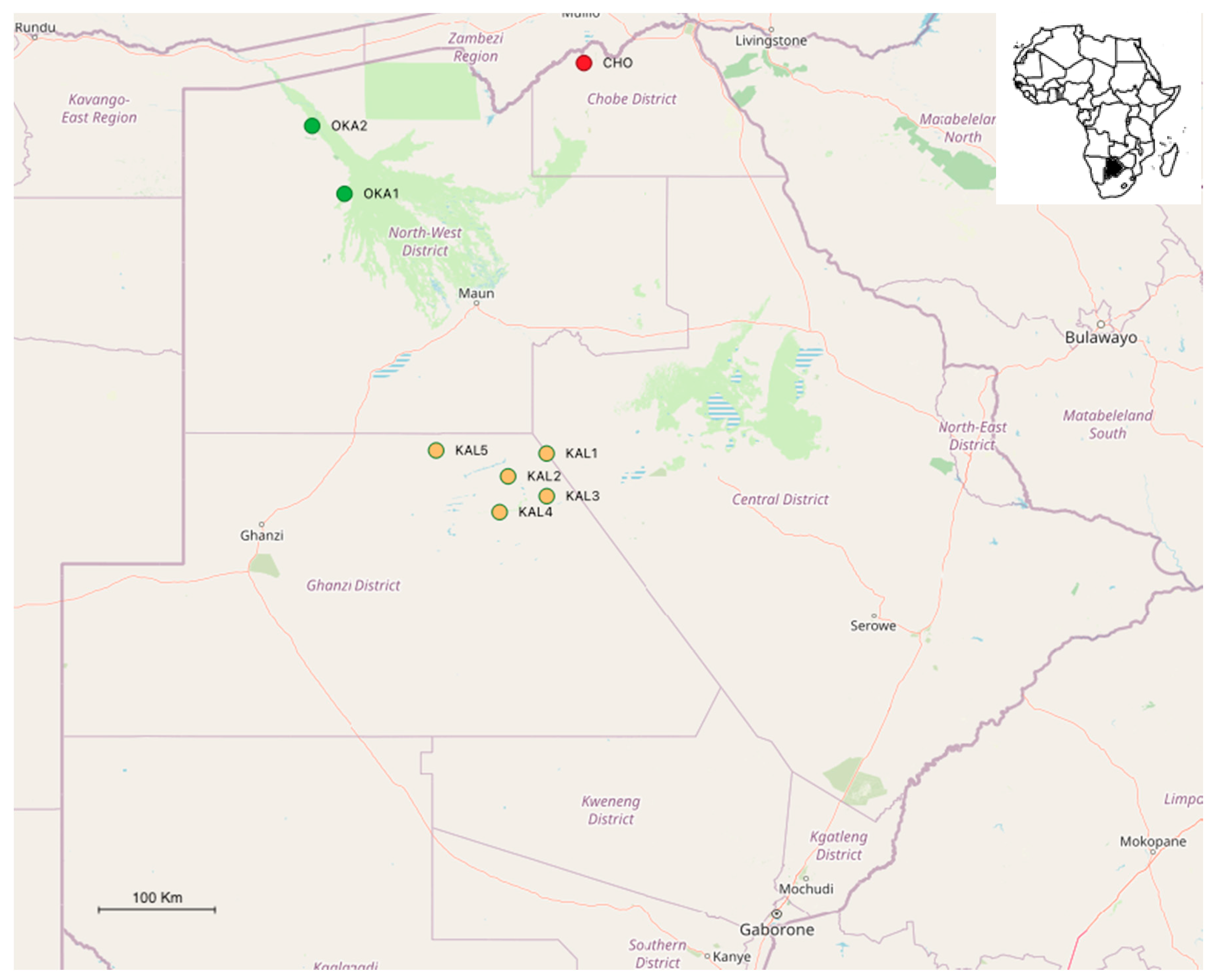
2.1. Study Site and Mosquito Collection
2.2. Sample Identification and Analyses
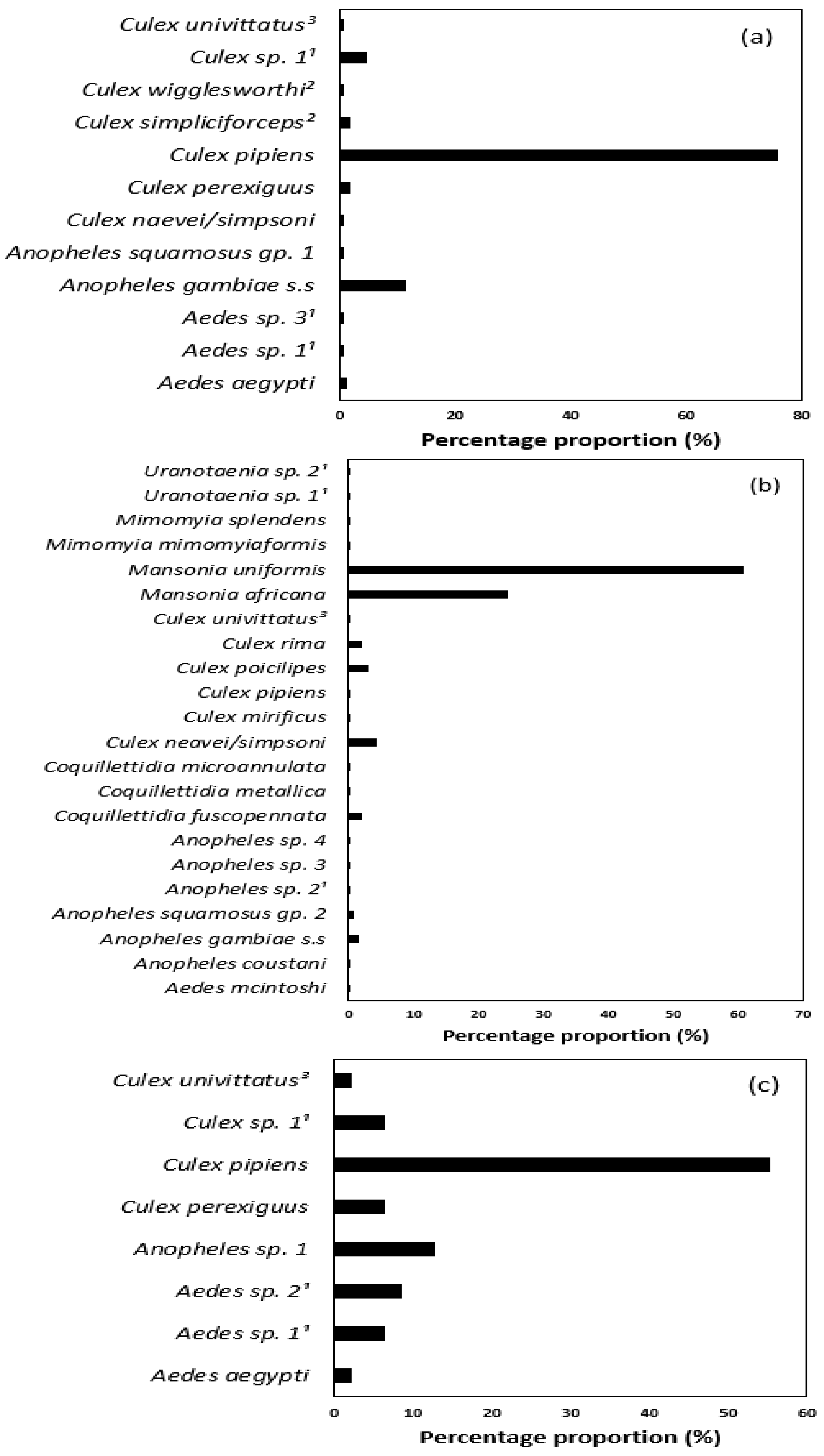
3. Results
4. Discussions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Portillo, A.; Ruiz-Arrondo, I.; Oteo, J.A. Arthropods as vectors of transmissible diseases in Spain. Med. Clin. 2018, 151, 450–459. [Google Scholar] [CrossRef]
- Santos, J.C.; de Almeida, W.R.; Fernandes, G.W. Arthropods: Why It Is So Crucial to Know Their Biodiversity? In Measuring Arthropod Biodiversity; Springer International Publishing: Cham, Switzerland, 2021; pp. 3–11. [Google Scholar]
- Lebl, K.; Zittra, C.; Silbermayr, K.; Obwaller, A.; Berer, D.; Brugger, K.; Rubel, F. Mosquitoes (Diptera: Culicidae) and their relevance as disease vectors in the city of Vienna, Austria. Parasitol. Res. 2015, 114, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Rizzoli, A.; Tagliapietra, V.; Cagnacci, F.; Marini, G.; Arnoldi, D.; Rosso, F.; Rosà, R. Parasites and wildlife in a changing world: The vector-host-pathogen interaction as a learning case. Int. J. Parasitol. Parasites Wildl. 2019, 9, 394–401. [Google Scholar] [CrossRef]
- Azari-Hamidian, S.; Norouzi, B.; Harbach, R.E. A detailed review of the mosquitoes (Diptera: Culicidae) of Iran and their medical and veterinary importance. Acta Trop. 2019, 194, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Rubio-Palis, Y.; Chareonviriyaphap, T.; Coetzee, M.; Hay, S.I. A global map of dominant malaria vectors. Parasites Vectors 2012, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Carlson, J.S.; Walther, E.; TroutFryxell, R.; Staley, S.; Tell, L.A.; Sehgal, R.N.; Cornel, A.J. Identifying avian malaria vectors: Sampling methods influence outcomes. Parasites Vectors 2015, 8, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, R.; Arum, S.; Chepkorir, E.; Mosomtai, G.; Tigoi, C.; Sigei, F.; Evander, M. Distribution and abundance of key vectors of Rift Valley fever and other arboviruses in two ecologically distinct counties in Kenya. PLoS Negl. Trop. Dis. 2017, 11, e0005341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braack, L.; De Almeida, A.P.G.; Cornel, A.J.; Swanepoel, R.; De Jager, C. Mosquito-borne arboviruses of African origin: Review of key viruses and vectors. Parasites Vectors. 2018, 11, 1–26. [Google Scholar] [CrossRef]
- Nkumama, I.N.; O’Meara, W.P.; Osier, F.H. Changes in malaria epidemiology in Africa and new challenges for elimination. Trends Parasitol. 2017, 33, 128–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Essl, F. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Chaves, L.S.M.; Fry, J.; Malik, A.; Geschke, A.; Sallum, M.A.M.; Lenzen, M. Global consumption and international trade in deforestation-associated commodities could influence malaria risk. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schrama, M.; Hunting, E.R.; Beechler, B.R.; Guarido, M.M.; Govender, D.; Nijland, W.; Gorsich, E.E. Human practices promote presence and abundance of disease-transmitting mosquito species. Sci. Rep. 2020, 10, 1–6. [Google Scholar] [CrossRef]
- Diagne, C.; Leroy, B.; Vaissière, A.C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Renault, D.; Angulo, E.; Cuthbert, R.; Haubrock, P.J.; Capinha, C.; Kramer, A.M.; Courchamp, F. The magnitude, diversity, and distribution of the economic costs of invasive terrestrial invertebrates worldwide. Biol. Invasions 2021, 23. [Google Scholar] [CrossRef]
- Carvajal-Lago, L.; Ruiz-López, M.J.; Figuerola, J.; Martínez-de la Puente, J. Implications of diet on mosquito life history traits and pathogen transmission. Environ. Res. 2021, 195, 110893. [Google Scholar] [CrossRef] [PubMed]
- Cunze, S.; Kochmann, J.; Koch, L.K.; Klimpel, S. Niche conservatism of Aedes albopictus and Aedes aegypti-two mosquito species with different invasion histories. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munyao, V.; Karisa, J.; Munyao, C.M.; Ngari, M.; Menza, N.; Peshu, N.; Mwangangi, J. Surveillance of Culicine Mosquitoes in Six Villages of Taita-Taveta County, Kenya, With Host Determinations From Blood-Fed Females. J. Med. Entomol. 2020, 57, 1972–1982. [Google Scholar] [CrossRef] [PubMed]
- Joannides, J. Species Composition and Risk of Transmission of Aedes-Borne Arboviruses around the Mole Game Reserve in Northern Ghana. Ph.D. Thesis, University of Ghana, Accra, Ghana, 2019. [Google Scholar]
- Mbacham, W.F.; Ayong, L.; Guewo-Fokeng, M.; Makoge, V. Current Situation of Malaria in Africa. In Malaria Control and Elimination; Humana Press: New York, NY, USA, 2019; pp. 29–44. [Google Scholar]
- Willcox, A.C.; Collins, M.H.; Jadi, R.; Keeler, C.; Parr, J.B.; Mumba, D.; Meshnick, S.R. Seroepidemiology of dengue, Zika, and yellow fever viruses among children in the Democratic Republic of the Congo. Am. J. Trop. Med. Hyg. 2018, 99, 756–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwanyika, G.O.; Mboera, L.E.; Rugarabamu, S.; Ngingo, B.; Sindato, C.; Lutwama, J.J.; Misinzo, G. Dengue Virus Infection and Associated Risk Factors in Africa: A Systematic Review and Meta-Analysis. Viruses 2021, 13, 536. [Google Scholar] [CrossRef]
- Dyer, O. African malaria deaths set to dwarf covid-19 fatalities as pandemic hits control efforts, WHO warns. BMJ 2020, 371, m4711. [Google Scholar] [CrossRef]
- Stevenson, J.C.; Norris, D.E. Implicating cryptic and novel anophelines as malaria vectors in Africa. Insects 2017, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Kgoroebutswe, T.K.; Ramatlho, P.; Reeder, S.; Makate, N.; Paganotti, G.M. Distribution of Anopheles mosquito species, their vectorial role and profiling of knock-down resistance mutations in Botswana. Parasitol. Res. 2020, 119, 1201–1208. [Google Scholar] [CrossRef]
- Tawe, L.; Ramatlho, P.; Waniwa, K.; Muthoga, C.W.; Makate, N.; Ntebela, D.S.; Paganotti, G.M. Preliminary survey on Anopheles species distribution in Botswana shows the presence of Anopheles gambiae and Anopheles funestus complexes. Malar. J. 2017, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Leechor, C. Developing Tourism in Botswana: Progress and Challenges; Botswana Documents; Botswana institute for Development Policy Analysis: Gaborone, Botswana, 2017. [Google Scholar]
- Stone, T.M.; Mbaiwa, J.E.; Stone, L.S. Tourism in Botswana in the last 50 years: A review. Botsw. Notes Rec. 2017, 49, 57–72. [Google Scholar]
- Magole, L.I.; Gojamang, O. The dynamics of tourist visitation to national parks and game reserves in Botswana. Botsw. Notes Rec. 2005, 37, 80–96. [Google Scholar]
- Pachka, H.; Annelise, T.; Alan, K.; Power, T.; Patrick, K.; Véronique, C.; Ferran, J. Rift Valley fever vector diversity and impact of meteorological and environmental factors on Culex pipiens dynamics in the Okavango Delta, Botswana. Parasites Vectors 2016, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cornel, A.J.; Lee, Y.; Almeida, A.P.G.; Johnson, T.; Mouatcho, J.; Venter, M.; Braack, L. Mosquito community composition in South Africa and some neighboring countries. Parasites Vectors 2018, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karypidou, M.C.; Almpanidou, V.; Tompkins, A.M.; Mazaris, A.D.; Gewehr, S.; Mourelatos, S.; Katragkou, E. Projected shifts in the distribution of malaria vectors due to climate change. Clim. Chang. 2020, 163, 2117–2133. [Google Scholar] [CrossRef]
- Buxton, M.; Wasserman, R.J.; Nyamukondiwa, C. Disease Vector Relative Spatio-Temporal Abundances to Water Bodies and Thermal Fitness Across Malaria Endemic Semi-Arid Areas. J. Med. Entomol. 2021, 58, 682–691. [Google Scholar] [CrossRef]
- Bango, Z.A.; Tawe, L.; Muthoga, C.W.; Paganotti, G.M. Past and current biological factors affecting malaria in the low transmission setting of Botswana: A review. Infect. Genet. Evol. 2020, 85, 104458. [Google Scholar] [CrossRef]
- Abdulla-Khan, R. Bionomics and cytogenetics of Anopheles seretsei Abdulla-Khan, Coetzee, and Hunt, a new species from northern Botswana. J. Am. Mosq. Control. Assoc. 1998, 14, 253–255. [Google Scholar]
- Buxton, M.; Lebani, K.; Nyamukondiwa, C.; Wasserman, R.J. First record of Aedes (Stegomyia) aegypti (Linnaeus, 1762) (Diptera: Culicidae) in Botswana. Bioinvasions Rec. 2019, 8, 551–557. [Google Scholar] [CrossRef]
- Buxton, M.; Cuthbert, R.N.; Dalu, T.; Nyamukondiwa, C.; Wasserman, R.J. Cattle-induced eutrophication favours disease-vector mosquitoes. Sci. Total Environ. 2020, 715, 136952. [Google Scholar] [CrossRef]
- Johnson, T.; Braack, L.; Guarido, M.; Venter, M.; Gouveia Almeida, A.P. Mosquito community composition and abundance at contrasting sites in northern South Africa, 2014–2017. J. Vector Ecol. 2020, 45, 104–117. [Google Scholar] [CrossRef]
- Mishra, N.B.; Crews, K.A.; Miller, J.A.; Meyer, T. Mapping vegetation morphology types in southern Africa savanna using MODIS time-series metrics: A case study of central Kalahari, Botswana. Land 2015, 4, 197–215. [Google Scholar] [CrossRef] [Green Version]
- Ramberg, L.; Hancock, P.; Lindholm, M.; Meyer, T.; Ringrose, S.; Sliva, J.; Vander Post, C. Species diversity of the Okavango delta, Botswana. Aquat. Sci. 2006, 68, 310–337. [Google Scholar] [CrossRef]
- Makhabu, S.W. Resource partitioning within a browsing guild in a key habitat, the Chobe Riverfront, Botswana. J. Trop. Ecol. 2005, 21, 641–649. [Google Scholar] [CrossRef]
- Jori, F.; Mokospasetso, M.; Etter, E.; Munstermann, S.; Newman, S.H.; Michel, A. Preliminary Assessment of Bovine Tuberculosis at the Livestock/Wildlife Interface in two Protected Areas of Northern Botswana. Transbound. Emerg. Dis. 2013, 60, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jupp, P.G. Mosquitoes of Southern Africa: Culicinae and toxorhynchitinae; Ekogilde Publishers: Hartebeespoort, South Africa, 1996. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Khoobdel, M.; Keshavarzi, D.; Mossa-Kazemi, S.H.; Sobati, H. Species diversity of mosquitoes of the Genus Culex (Diptera, Culicidae) in the coastal areas of the Persian Gulf. AIMS Public Health 2019, 6, 99. [Google Scholar] [CrossRef] [PubMed]
- Weetman, D.; Kamgang, B.; Badolo, A.; Moyes, C.L.; Shearer, F.M.; Coulibaly, M.; McCall, P.J. Aedes mosquitoes and Aedes-borne arboviruses in Africa: Current and future threats. Int. J. Environ. Res. Public Health 2018, 15, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprygin, A.; Pestova, Y.; Wallace, D.B.; Tuppurainen, E.; Kononov, A.V. Transmission of lumpy skin disease virus: A short review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.P.; Reuman, D.C.; Lutomiah, J.; Peterson, A.T.; Linthicum, K.J.; Britch, S.C.; Sang, R. Predicting Abundances of Aedes mcintoshi, a primary Rift Valley fever virus mosquito vector. PLoS ONE 2019, 14, e0226617. [Google Scholar] [CrossRef] [PubMed]
- Goupeyou-Youmsi, J.; Rakotondranaivo, T.; Puchot, N.; Peterson, I.; Girod, R.; Vigan-Womas, I.; Bourgouin, C. Differential contribution of Anopheles coustani and Anopheles arabiensis to the transmission of Plasmodium falciparum and Plasmodium vivax in two neighbouring villages of Madagascar. Parasites Vectors 2020, 13, 1–16. [Google Scholar] [CrossRef]
- Tantely, L.M.; Boyer, S.; Fontenille, D. A review of mosquitoes associated with Rift Valley fever virus in Madagascar. Am. J. Trop. Med. Hyg. 2015, 92, 722–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreaux, A.M.; Stone, C.M.; Barreaux, P.; Koella, J.C. The relationship between size and longevity of the malaria vector Anopheles gambiae (ss) depends on the larval environment. Parasites Vectors 2018, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Socolovschi, C.; Pages, F.; Ndiath, M.O.; Ratmanov, P.; Raoult, D. Rickettsia species in African Anopheles mosquitoes. PLoS ONE 2012, 7, e48254. [Google Scholar] [CrossRef] [Green Version]
- Fornadel, C.M.; Norris, L.C.; Franco, V.; Norris, D.E. Unexpected anthropophily in the potential secondary malaria vectors Anopheles coustani sl and Anopheles squamosus in Macha, Zambia. Vector Borne Zoonotic Dis. 2011, 11, 1173–1179. [Google Scholar] [CrossRef] [Green Version]
- Mutebi, J.P.; Crabtree, M.B.; Kading, R.C.; Powers, A.M.; Lutwama, J.J.; Miller, B.R. Mosquitoes of western Uganda. J. Med. Entomol. 2014, 49, 1289–1306. [Google Scholar] [CrossRef]
- Njabo, K.Y.; Cornel, A.J.; Bonneaud, C.; Toffelmier, E.; Sehgal, R.N.M.; Valkiūnas, G.; Smith, T.B. Nonspecific patterns of vector, host and avian malaria parasite associations in a central African rainforest. Mol. Ecol. 2011, 20, 1049–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Njabo, K.Y.; Cornel, A.J.; Sehgal, R.N.; Loiseau, C.; Buermann, W.; Harrigan, R.J.; Smith, T.B. Coquillettidia (Culicidae, Diptera) mosquitoes are natural vectors of avian malaria in Africa. Malar. J. 2009, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fall, A.G.; Diaïté, A.; Lancelot, R.; Tran, A.; Soti, V.; Etter, E.; Bouyer, J. Feeding behaviour of potential vectors of West Nile virus in Senegal. Parasites Vectors 2011, 4, 1–7. [Google Scholar] [CrossRef]
- Ferraguti, M.; Heesterbeek, H.; Martínez-de la Puente, J.; Jiménez-Clavero, M.Á.; Vázquez, A.; Ruiz, S.; Figuerola, J. The role of different Culex mosquito species in the transmission of West Nile virus and avian malaria parasites in Mediterranean areas. Transbound. Emerg. Dis. 2020, 68, 920–930. [Google Scholar] [CrossRef]
- Woods, J.A. Lumpy Skin Disease Virus. In Virus Infections of Ruminants; Elsevier Science publishers: Amsterdam, The Netherlands, 1990; pp. 53–67. [Google Scholar]
- Traoré, M.; Ouattara, A.F.; Coulibaly, D.; GN, C.; Gbalégba, D.S.T.; Kwadjo, K.E.; Doumbia, M. Assessment of the risk of mosquitoes nuisance and appearance of vector-borne diseases in rubber production areas in Southern Côte d’Ivoire. J. Entomol. Zool. Stud. 2019, 7, 322–329. [Google Scholar]
- Ughasi, J.; Bekard, H.E.; Coulibaly, M.; Adabie-Gomez, D.; Gyapong, J.; Appawu, M.; Boakye, D.A. Mansonia africana and Mansonia uniformis are vectors in the transmission of Wuchereria bancrofti lymphatic filariasis in Ghana. Parasites Vectors 2012, 5, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Osei-Poku, J.; Han, C.; Mbogo, C.M.; Jiggins, F.M. Identification of Wolbachia strains in mosquito disease vectors. PLoS ONE 2012, 7, e49922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traore-Lamizana, M.; Zeller, H.G.; Mondo, M.; Hervy, J.P.; Adam, F.; Digoutte, J.P. Isolations of West Nile and Bagaza viruses from mosquitoes (Diptera: Culicidae) in central Senegal (Ferlo). J. Med. Entomol. 1994, 31, 934–938. [Google Scholar] [CrossRef]
- Sabesan, S.; Kumar, N.P.; Krishnamoorthy, K.; Panicker, K.N. Seasonal abundance and biting behaviour of Mansonia annulifera. M. uniformis and M. indiana and their relative role in the transmission of malayan filariasis in shertallai (Kerala state). Indian J. Med. Res. 1991, 93, 253–258. [Google Scholar]
- Ridha, M.R.; Rahayu, N.; Hairani, B.; Perwitasari, D.; Kusumaningtyas, H. Biodiversity of mosquitoes and Mansonia uniformis as a potential vector of Wuchereria bancrofti in Hulu Sungai Utara District, South Kalimantan, Indonesia. Vet. World 2020, 13, 2815. [Google Scholar] [CrossRef]
- Zittra, C.; Vitecek, S.; Obwaller, A.G.; Rossiter, H.; Eigner, B.; Zechmeister, T.; Waringer, J.; Fuehrer, H.P. Landscape structure affects distribution of potential disease vectors (Diptera: Culicidae). Parasites Vectors 2017, 10, 205. [Google Scholar] [CrossRef] [Green Version]
- Johnson, P.H.; Russell, R.C. Colonization of Coquillettidia linealis (Skuse) with reference to other Coquillettidia and Mansonia species. J. Vector Ecol. 2019, 44, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Dida, G.O.; Anyona, D.N.; Abuom, P.O.; Akoko, D.; Adoka, S.O.; Matano, A.S.; Ouma, C. Spatial distribution and habitat characterization of mosquito species during the dry season along the Mara River and its tributaries, in Kenya and Tanzania. Infect. Dis. Poverty 2018, 7, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatem, A.J. Mapping population and pathogen movements. Int. Health 2014, 6, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Chirebvu, E.; Chimbari, M.J. Characteristics of Anopheles arabiensis larval habitats in Tubu village, Botswana. J. Vector Ecol. 2015, 40, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Ong, J.; Aik, J.; Ng, L.C. Adult Aedes abundance and risk of dengue transmission. PLoS Negl. Trop. Dis. 2021, 15, e0009475. [Google Scholar] [CrossRef] [PubMed]
- Kline, D.L. Traps and trapping techniques for adult mosquito control. J. Am. Mosq. Control. Assoc. 2006, 22, 490–496. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Anopheles | Aedes | Culex | Mansonia |
|---|---|---|---|
| An. arabiensis | Ae. mcintoshi | Cx. pipiens | Ma. uniformis |
| An. parensis | Ae. aegypti | Cx. poicillipes | |
| An. longipalpis type C | Cx. neavei | ||
| An. leesoni | Cx. antennatus | ||
| An. quadriannulatus | |||
| An. funestus s.s | |||
| An. rivulorum | |||
| An. pharoensis | |||
| An. nili | |||
| An. rufipes | |||
| An. distinctus | |||
| An. squamosus | |||
| An. ziemanni | |||
| An. demeilloni | |||
| An. marshalli | |||
| An. vaneedeni | |||
| An. rhodesiensis | |||
| An. seretsei | |||
| Refs [26,31,33,34,35] | [31,36] | [30,31,37] | [31] |
| Location | Site | Sampling Dates | Station Code | GPS Coordinates |
|---|---|---|---|---|
| Chobe enclave | VanThuyne-Ridge Research Centre | 18–22nd March 2019 | CHO | 18°06′44.5″ S 24°18′58.4″ E |
| 8–12th February 2020 | CHO | |||
| Okavango | Etsha | 3–5th February 2020 | OKA1 | 19°08′18.1″ S 22°20′04.1″ E |
| Nxamasere | 6–7th February 2020 | OKA2 | 18°36′22.6″ S 22°03′59.1″ E | |
| Central Kalahari Game Reserve | Matswere Gate | 12th March 2019 | CKGR1 | 21°09′23.4″ S 24°00′25.7″ E |
| Sunday Pan | 13th March 2019 | CKGR2 | 21°19′58.3″ S 23°41′18.4″ E | |
| Kalahari Plains Lodge | 14th March 2019 | CKGR3 | 21°29′07.9″ S 24°00′30.1″ E | |
| Letiahau | 15th March 2019 | CKGR4 | 21°36′30.3″ S 23°37′05.4″ E | |
| Motopi | 16th March 2019 | CKGR5 | 21°07′57.9″ S 23°05′34.8″ E |
| Genus | Species | Nb. Specimens | Identification (morph/mol.) | GenBank ID (% Identity) | ||
|---|---|---|---|---|---|---|
| CKGR | OKA | CHO | ||||
| Aedes | Ae. aegypti | 1 | 0 | 2 | morph/mol. | MK533633 (99.2–99.8) |
| Ae. mcintoshi | 0 | 3 | 0 | mol. | LC473695 (97.9–99.8) | |
| Ae. sp. 1 1 | 3 | 0 | 1 | morph. | - | |
| Ae. sp. 2 1 | 4 | 0 | 0 | morph. | - | |
| Ae. sp. 3 1 | 0 | 0 | 1 | morph. | - | |
| Anopheles | An. coustani | 0 | 10 | 0 | mol. | MK585951 (98.3–99.7) |
| An. gambiae s.s | 0 | 74 | 20 | morph/mol. | NC_002084 (99.7–100) | |
| An. squamosus gp. 1 | 0 | 0 | 1 | mol. | MK776750 (99.7) | |
| An. squamosus gp. 2 | 0 | 34 | 0 | mol. | MK533644 (99.1) | |
| An. sp. 1 | 6 | 0 | 0 | morph. | An. sp. 1 MT741511 (100) | |
| An. sp. 2 1 | 0 | 1 | 0 | morph. | - | |
| An. sp. 3 | 0 | 1 | 0 | morph. | An. sp. Mali 1 MK585979 (99.4) | |
| An. sp. 4 | 0 | 1 | 0 | morph. | An. sp. 15 MK776739 (98.7) | |
| Coquillettidia | Cq. fuscopennata | 0 | 111 | 0 | morph/mol. | LC473712 (98.3–99.1) |
| Cq. metallica | 0 | 4 | 0 | morph/mol. | LC473709 (99.8) | |
| Cq. microannulata | 0 | 2 | 0 | morph/mol. | LC473713 (99.2–99.4) | |
| Culex | Cx. neavei/simpsoni | 0 | 229 | 1 | morph/mol. | Cx. neavei LC473635 (97.4–98.6) Cx. simpsoni KU187061 (96.9–98.0) |
| Cx. mirificus | 0 | 1 | 0 | mol. | LC473643 (100) | |
| Cx. perexiguus | 3 | 0 | 3 | mol. | KU380423 (99.7–100) | |
| Cx. pipiens | 26 | 10 | 131 | morph/mol. | MZ206334 (99.7–100) | |
| Cx. poicilipes | 0 | 164 | 0 | morph/mol. | LC473618 (98.2–99.7) | |
| Cx. rima | 0 | 111 | 0 | morph/mol. | KU380462 (99.2–99.7) | |
| Cx. simpliciforceps2 | 0 | 0 | 3 | morph. | Cx. sp. LC507872 (98.8–99.1) | |
| Cx. wigglesworthi2 | 0 | 0 | 1 | morph. | Cx. rima LC473615 (97.3) | |
| Cx. sp. 1 1 | 3 | 0 | 8 | mol. | Cx. sp. 16GH LC507872 (98.9–99.8) Cx. cinereus LC473617 (97.0) | |
| Cx. univittatus3 | 1 | 8 | 1 | morph/mol. | LC102144 (99.0–99.7) | |
| Mansonia | Ma. africana | 0 | 1290 | 0 | morph/mol. | KU380478 (98.5–99.1) |
| Ma. uniformis | 0 | 3207 | 0 | morph/mol. | KU187168 (99.8–100) | |
| Mimomyia | Mi. mimomyiaformis | 0 | 1 | 0 | mol. | LC473719 (100) |
| Mi. splendens | 0 | 2 | 0 | morph/mol. | KU380391 (99.0–99.1) | |
| Uranotaenia | Ur. sp. 1 1 | 0 | 1 | 0 | morph. | - |
| Ur. sp. 2 1 | 0 | 1 | 0 | morph. | - | |
| TOTAL | 47 | 5266 | 173 | |||
| Species | Main Associated Diseases | Preferred Hosts: Humans (H), Livestock (L), Wildlife (W) | References |
|---|---|---|---|
| Aedes aegypti | Dengue, yellow fever, Zika, Chikungunya Lumpy skin disease | H, L, W | [46,47] |
| Aedes mcintoshi | Rift Valley fever | L, W | [48] |
| Anopheles coustani | Human malaria Rift Valley fever, West Nile fever, Chikungunya | H, L, W | [49] |
| Anopheles gambiae s.s | Human malaria | H, L, W | [50,51,52] |
| Rift Valley fever Cat-flea typhus Spotted fever | |||
| Anopheles squamosus gp. 1 Anopheles squamosus gp. 2 | Human malaria Rift Valley fever | H, W | [53] |
| Coquillettidia fuscopennata | Avian malaria, Chikungunya, yellow fever, Sindbis fever | H, W | [54,55,56] |
| Coquillettidia metallica | Avian malaria, West Nile fever, Rift Valley fever | H, W | [54,55,56] |
| Coquillettidia microannulata | Avian malaria | W | [55,56] |
| Culex naevei | West Nile fever, avian malaria | W | [55,57] |
| Culex perexiguus | West Nile fever, avian malaria | H, W | [58] |
| Culex pipiens | Rift Valley fever, West Nile fever, Filariasis, Encephalitis, Avian malaria | H, L, W | [50,58] |
| Culex mirificus | Lumpy skin disease | L, W | [47,59] |
| Culex poicilipes | Rift Valley fever, avian malaria | L, W | [55,57] |
| Culex rima | West Nile fever, encephalitis, avian malaria, filariasis | H, L, W | [60] |
| Culex simpliciforceps | [50,55] | ||
| Culex sp. 1 | Rift Valley fever, avian malaria | H, L, W | |
| Culex sp. 2 | |||
| Culex univittatus | West Nile fever, Wesselsbron | H, L, W | [54] |
| Mansonia africana | Rift Valley fever, lymphatic filariasis, West Nile fever | H, W | [50,54,61,62] |
| Mansonia uniformis | Rift Valley fever, lymphatic filariasis, Avian malaria, West Nile fever, Zika, Chikungunya | H, W | [50,54,55,61,62] |
| Mimomyia mimomyiaformis Mimomyia splendens | West Nile fever | H, W | [63] |
| Uranotaenia sp. 1 Uranotaenia sp. 2 | Wesselsbron | L | [54] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buxton, M.; Nyamukondiwa, C.; Wasserman, R.J.; Othenin-Girard, V.; Pigeault, R.; Christe, P.; Glaizot, O. Surveillance Studies Reveal Diverse and Potentially Pathogenic-Incriminated Vector Mosquito Species across Major Botswana Touristic Hotspots. Insects 2021, 12, 913. https://doi.org/10.3390/insects12100913
Buxton M, Nyamukondiwa C, Wasserman RJ, Othenin-Girard V, Pigeault R, Christe P, Glaizot O. Surveillance Studies Reveal Diverse and Potentially Pathogenic-Incriminated Vector Mosquito Species across Major Botswana Touristic Hotspots. Insects. 2021; 12(10):913. https://doi.org/10.3390/insects12100913
Chicago/Turabian StyleBuxton, Mmabaledi, Casper Nyamukondiwa, Ryan J. Wasserman, Victor Othenin-Girard, Romain Pigeault, Philippe Christe, and Olivier Glaizot. 2021. "Surveillance Studies Reveal Diverse and Potentially Pathogenic-Incriminated Vector Mosquito Species across Major Botswana Touristic Hotspots" Insects 12, no. 10: 913. https://doi.org/10.3390/insects12100913
APA StyleBuxton, M., Nyamukondiwa, C., Wasserman, R. J., Othenin-Girard, V., Pigeault, R., Christe, P., & Glaizot, O. (2021). Surveillance Studies Reveal Diverse and Potentially Pathogenic-Incriminated Vector Mosquito Species across Major Botswana Touristic Hotspots. Insects, 12(10), 913. https://doi.org/10.3390/insects12100913