
Prediction of Spatiotemporal Invasive Risk of the Red Import Fire Ant, Solenopsis invicta (Hymenoptera: Formicidae), in China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Species Overview
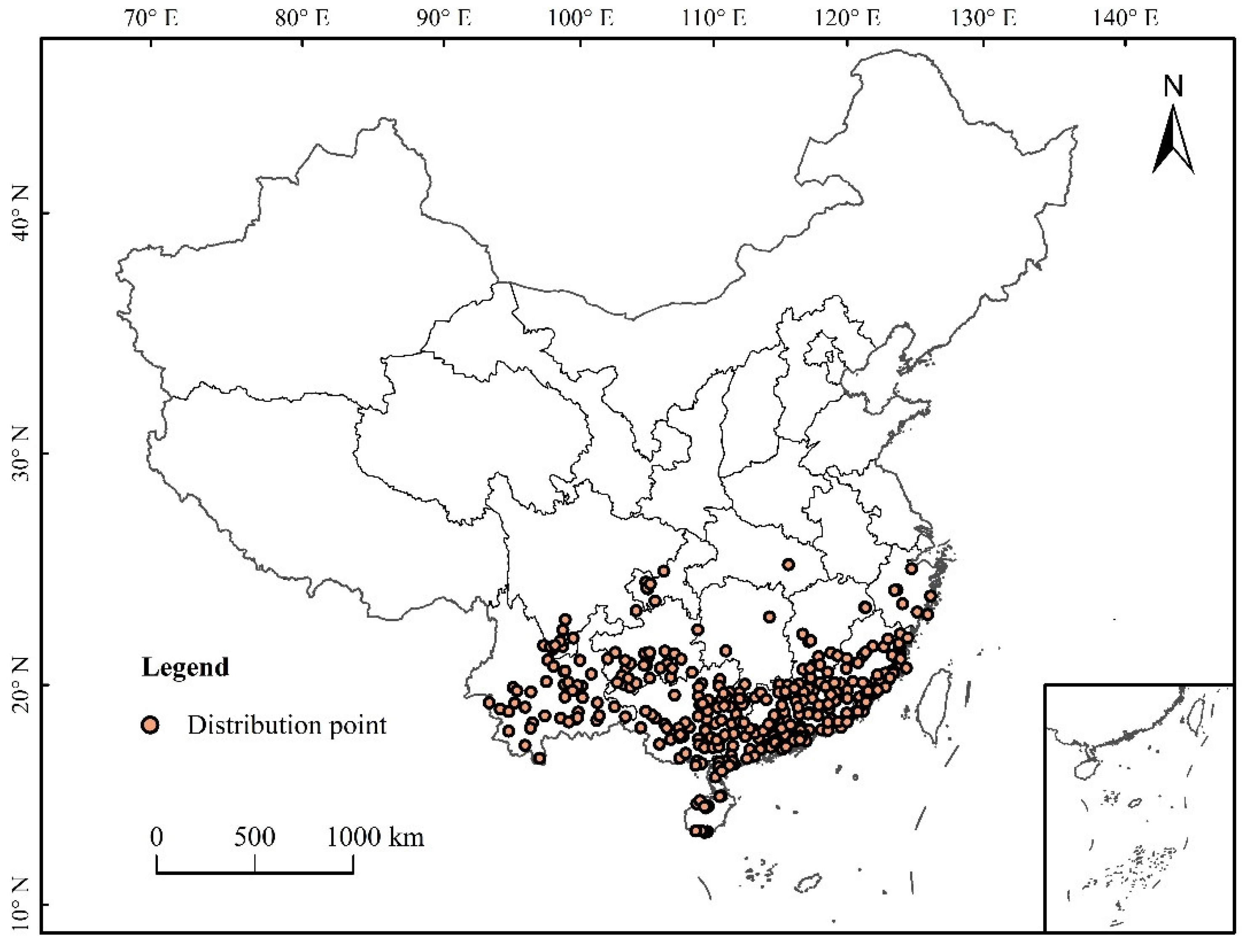
2.2. Data and Processing of Species Distribution
2.3. Environmental Factors and Processing
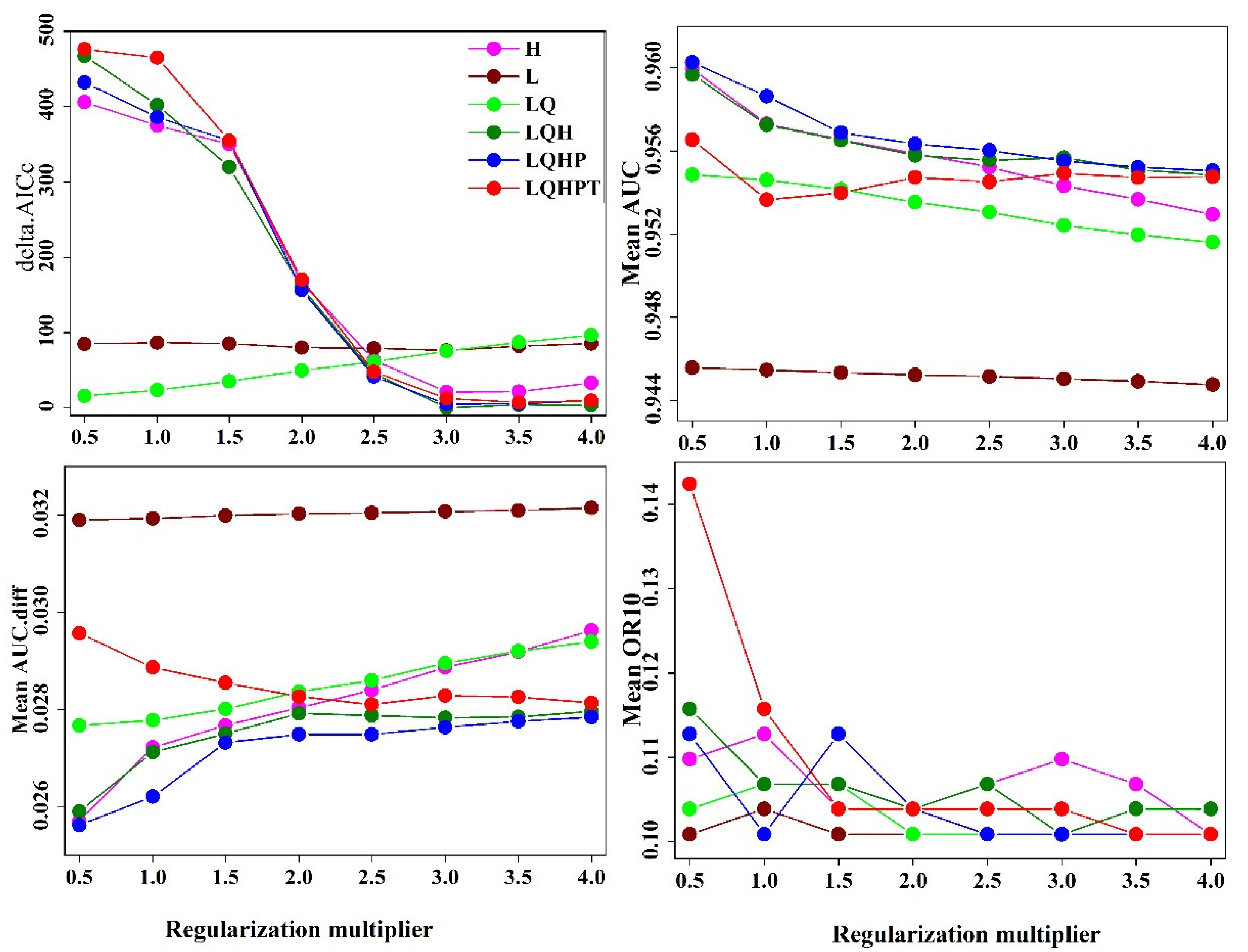
2.4. MaxEnt Model Construction and Parameter Optimization
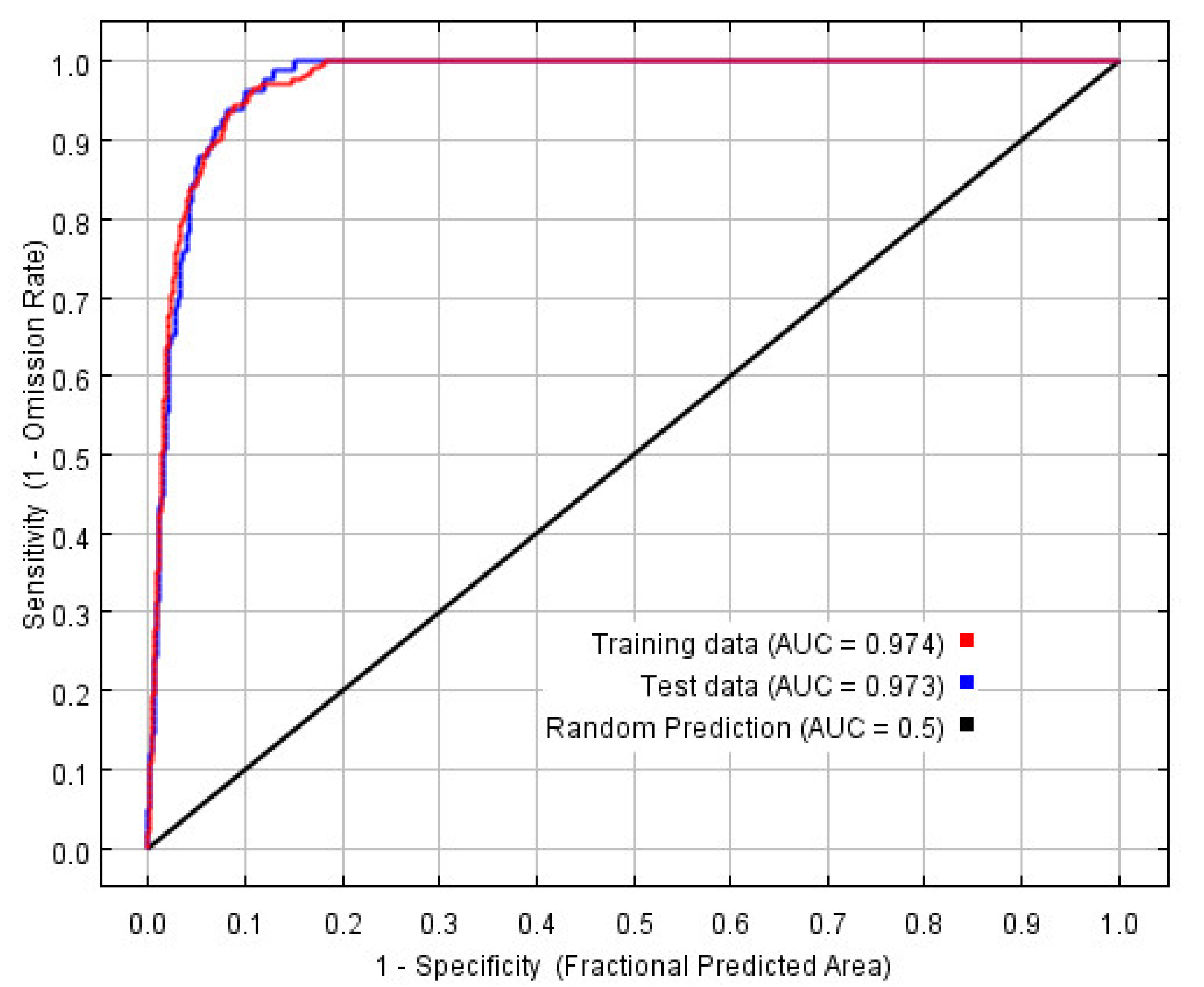
2.5. MaxEnt Model Evaluation
3. Results
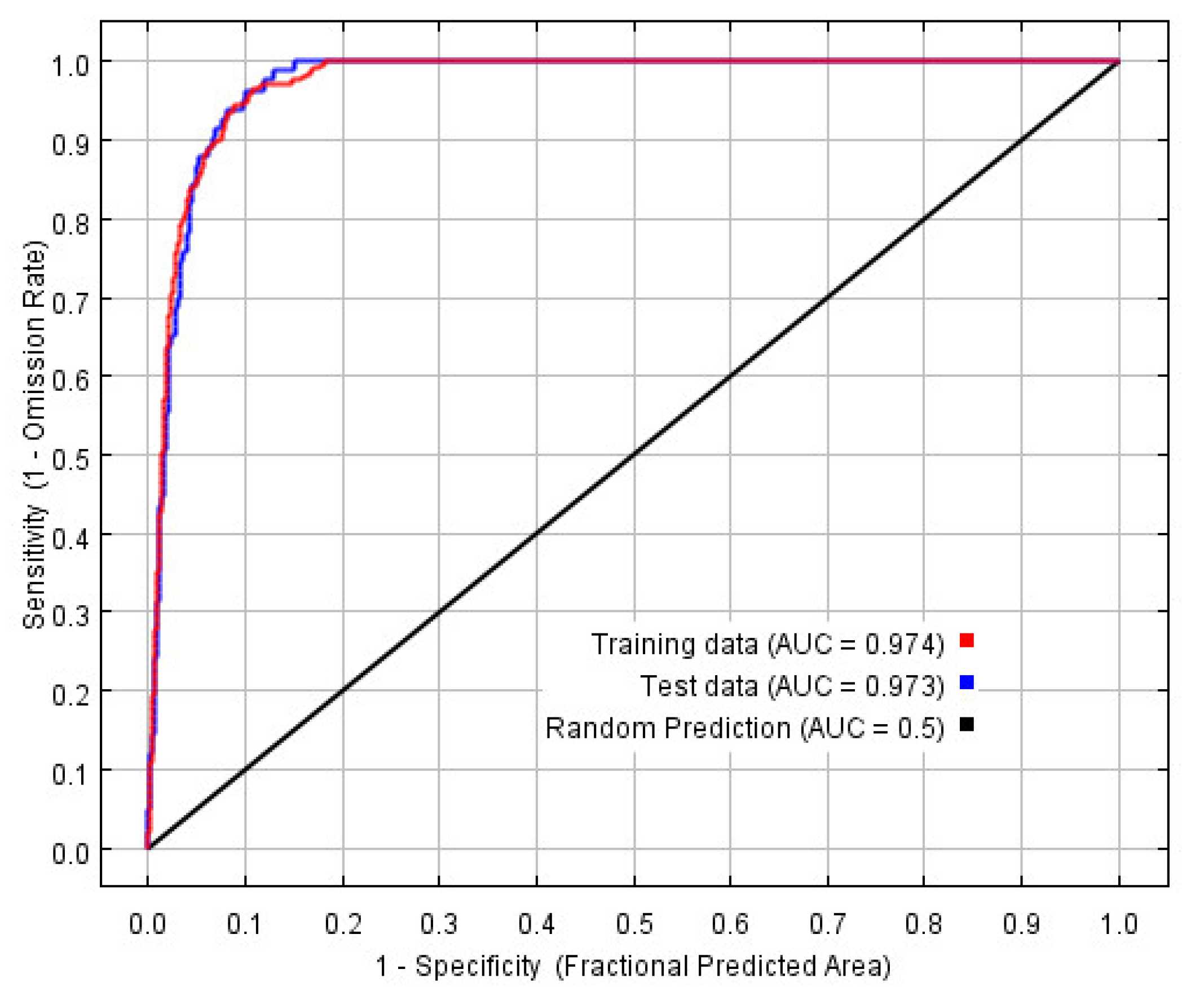
3.1. Optimal Model and Accuracy Evaluation
3.2. Dominant Environmental Factors
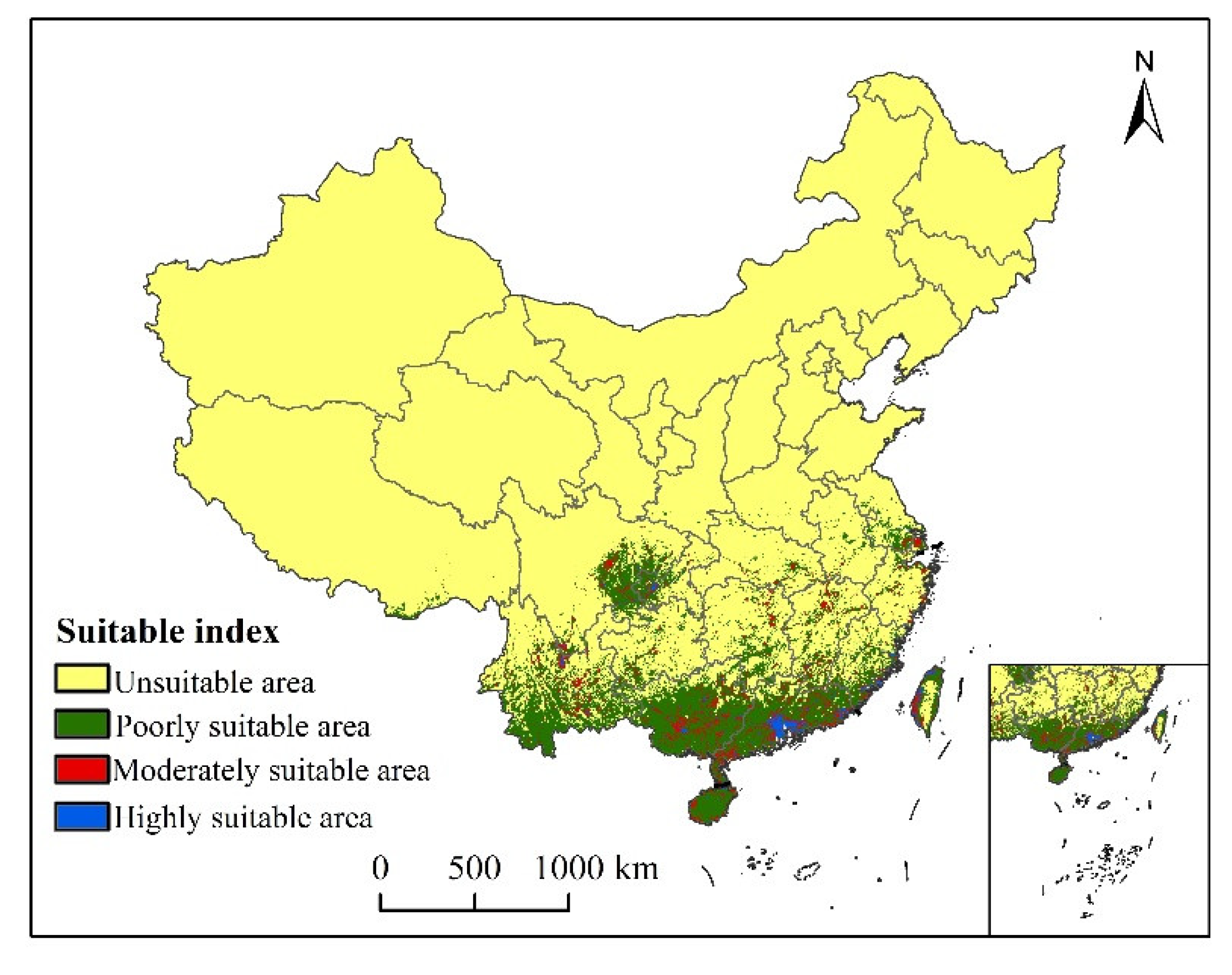
3.3. Suitable Areas under Current Conditions
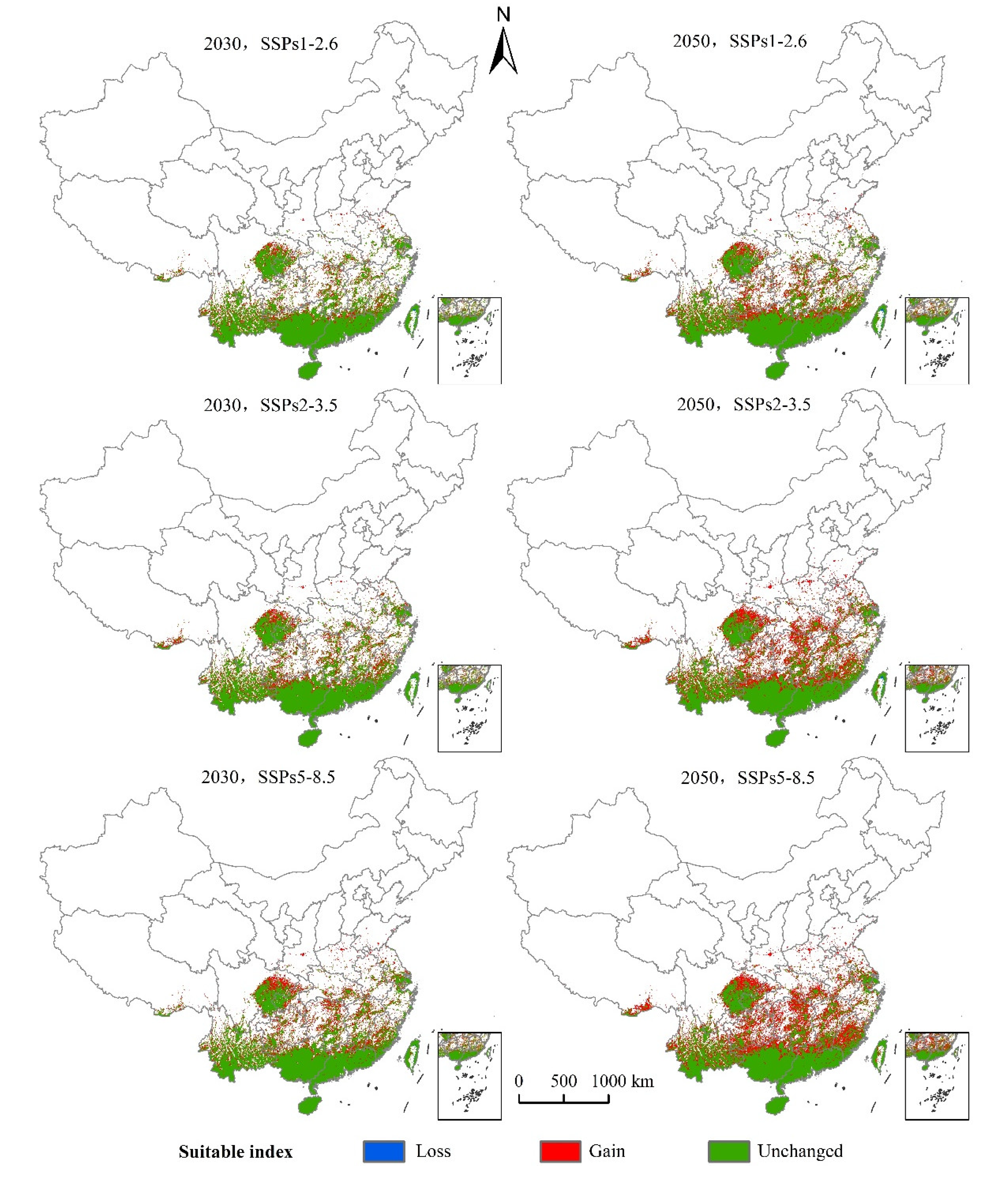
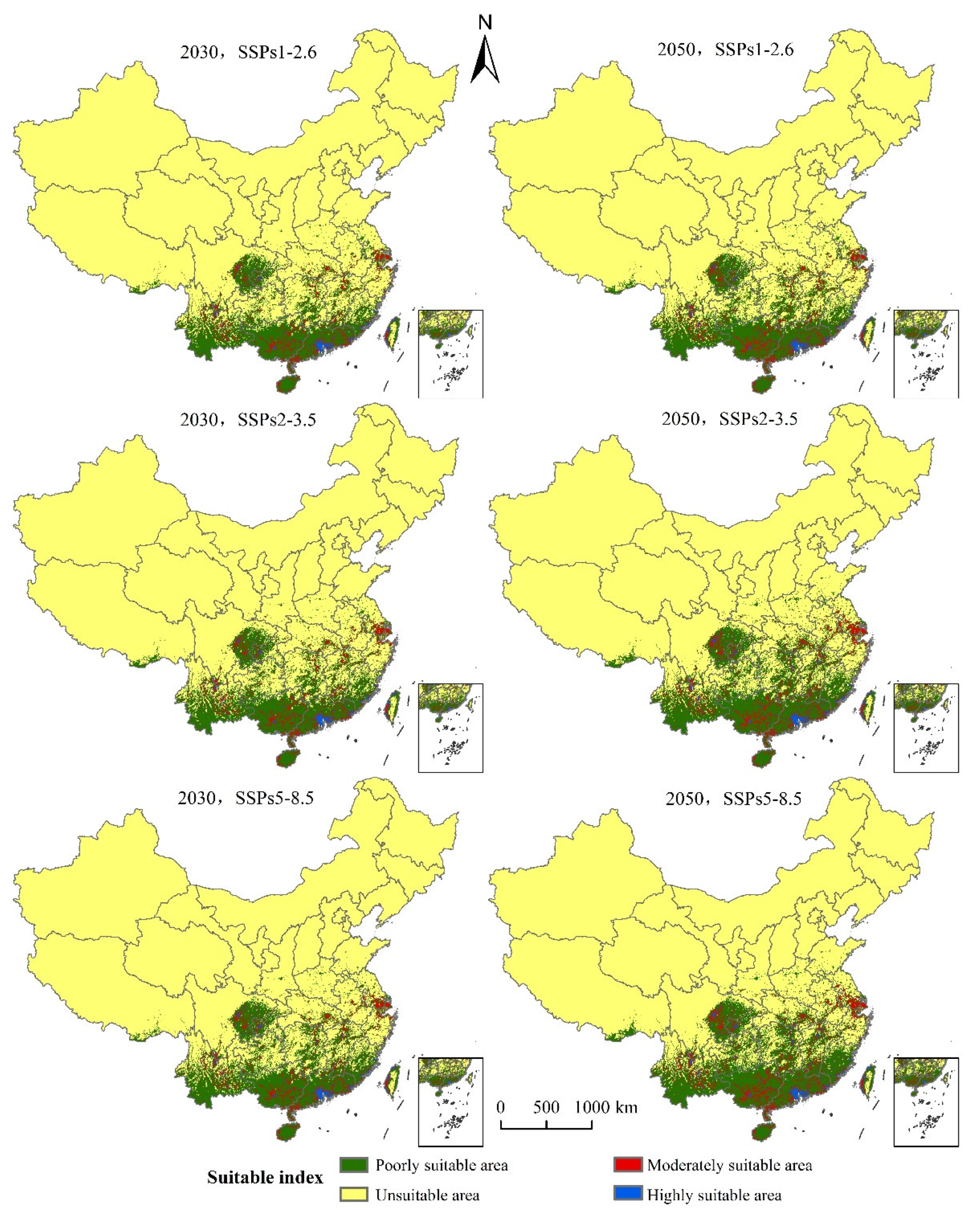
3.4. The Suitable Areas under Future Climate Change Scenarios
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ju, R.T.; Li, H.; Shi, Z.R.; Li, B. Progress of biological invasions research in China over the last decade. Biodivers. Sci. 2012, 20, 581–611. [Google Scholar]
- Peng, L.; Wan, F.H.; Hou, Y.P. Research progress on prevention and control of invasive insects in China. Chin. J. Appl. Entomol. 2020, 57, 244–258. [Google Scholar]
- Xu, H.G.; Qiang, S.; Genovesi, P.; Ding, H.; Wu, J.; Meng, L.; Han, Z.M.; Miao, J.L.; Hu, B.S.; Guo, J.Y.; et al. An inventory of invasive alien species in China. NeoBiota 2020, 15, 1–26. [Google Scholar] [CrossRef]
- Ying, Y.; Xian, X.; Jiang, M. Biological Invasion and Its Research in China: An Overview. Biol. Invasions Manag. China 2017, 56, 3–19. [Google Scholar]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Fang, F.; Zhang, C.X.; Huang, H.J.; Li, Y.; Chen, J.C.; Yang, L.; Wei, S.H. Potential distribution of Tausch’s goatgrass(Aegilops tauschii) in both China and the rest of the word as predicted by MaxEnt. Acta Pratacult. Sin. 2013, 22, 62–70. [Google Scholar]
- Li, M.Y.; Ju, Y.W.; Wu, W.H.; He, Y.J.; Xu, G.C. Dynamic analysis of potential habitat of alien forest invasive species under climate change scenarios: A case study of Dendroctonus frontalis. J. Beijing For. Univ. 2009, 31, 64–69. [Google Scholar]
- Jiang, Z.H.; Zhang, X.; Wang, J. Projection of climate change in China in the 21st century by IPCC-AR4 Models. Geogr. Res. 2008, 787–799. [Google Scholar]
- Wan, F.H.; Guo, J.Y.; Wang, D.H. Alien invasive species in China: Their damages and management strategies. Biodivers. Sci. 2002, 10, 119–125. [Google Scholar]
- Bullock, J.M.; Chapman, D.; Schafer, S.; Roy, D.; Brough, C. Assessing and Controlling the Spread and the Effects of Common Ragweed in Europe; Natural Environment Research Council: Swindon, UK, 2012; pp. 85–104. [Google Scholar]
- Zhou, W.C.; Cai, K.Z. An introduction to the alien invasive species of Solenopsis invicta in Taiwan. Taiwan Agric. Res. 2005, 39–41. [Google Scholar]
- Zeng, L.; Lu, Y.Y.; He, X.F.; Zhang, W.Q.; Liang, G.W. Identification of red imported fire ant Solenopsis invicta to invade mainland China and infestation in Wuchuan, Guangdong. Chin. J. Appl. Entomol. 2005, 42, 144–148. [Google Scholar]
- Sun, Z.J. Red imported fire ant: A dangerous alien species. Ecol. Econ. 2021, 37, 5–8. [Google Scholar]
- Fu, Q.Y.; Song, Z.D.; Zhao, Y.; Li, S.L.; Xu, Y.J. Analysis on the control cost of Solenopsis invicta in mainland China. J. Environ. Entomol. 2021. Available online: https://kns.cnki.net/kcms/detail/44.1640.q.20210223.0815.002.html. (accessed on 27 September 2021).
- Korzukhin, M.; Porter, S.D. Spatial Model of Territorial Competition and Population Dynamics in the Fire Ant Solenopsis invicta (Hymenoptera: Formicidae). Environ. Entomol. 1994, 23, 912–922. [Google Scholar] [CrossRef]
- Korzukhin, M.D.; Porter, S.D.; Thompson, L.C.; Wiley, S. Modeling temperature-dependent range limits for the fire ant Solenopsis invicta (Hymenoptera: Formicidae) in the United States. Environ. Entomol. 2001, 30, 645–655. [Google Scholar] [CrossRef] [Green Version]
- Morrison, L.W.; Porter, S.D.; Daniels, E.; Korzukhin, M. Potential Global Range Expansion of the Invasive Fire Ant, Solenopsis invicta. Biol. Invasions 2004, 6, 183–191. [Google Scholar] [CrossRef]
- Sutherst, R.W. A Climate Model of the Red Imported Fire Ant, Solenopsis invicta Buren (Hymenoptera: Formicidae): Implications for Invasion of New Regions, Particularly Oceania. Environ. Entomol. 2005, 34, 317–335. [Google Scholar] [CrossRef]
- Yaeger, W. Frequency of fire ant stinging in Lowndes County, Georgia. J. Med. Assoc. Ga. 1978, 67, 101–102. [Google Scholar]
- Hoffman, D.R. Reactions to less common species of fire ants. J. Allergy Clin. Immunol. 1997, 100, 679–683. [Google Scholar] [CrossRef]
- Allen, C.R.; Epperson, D.M.; Garmestani, A.S. Red imported fire ant impacts on wildlife: A decade of research. Am. Midl. Nat. 2014, 152, 88–103. [Google Scholar] [CrossRef] [Green Version]
- Ward, D.F. The potential distribution of the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae), in New Zealand. N. Z. Entomol. 2009, 32, 67–75. [Google Scholar] [CrossRef]
- Chen, N.Z.; Shi, Z.W.; Ma, X.G. Identification of red imported fire ant and important related species. Chin. Bull. Entomol. 2005, 341–345. [Google Scholar]
- Vinson, S.B. Invasion of the red imported fire ant (Hymenoptera: Formicidae): Spread, biology, and impact. Am. Entomol. 1997, 43, 23–39. [Google Scholar] [CrossRef]
- Song, Y.S. China’s Forestry Pest Survey Results in 2014–2017; China Forestry Publishing House: Beijing, China, 2019. [Google Scholar]
- Wang, R.L.; Li, Q.; Fen, C.H.; Shi, C.P. Predicting potential ecological distribution of Locusta migratoria tibetensis in China using MaxEnt ecological niche modeling. Acta Ecol. Sin. 2017, 37, 8556–8566. [Google Scholar]
- Xin, S.G.; Wu, T.W.; Zhang, J.; Zhang, F.; Li, W.P.; Zhang, Y.W.; Lu, Y.X.; Fang, Y.J.; Jie, W.H.; Zhang, L.; et al. Introduction of BCC models and its participation in CMIP6. Clim. Chang. Res. 2019, 15, 533–539. [Google Scholar]
- Weng, Y.W.; Cai, W.J.; Wang, C. The application and future directions of the Shared Socioeconomic Pathways (SSPs). Clim. Chang. Res. 2020, 16, 215–222. [Google Scholar]
- Wang, J.F.; Xu, C.D. Geodetector: Principle and prospective. Acta Geogr. Sin. 2017, 72, 116–134. [Google Scholar]
- Zhang, Q.; Zhang, D.F.; Wu, M.L.; Guo, J.Y.; Sun, C.Z.; Xie, C.X. Predicting the global areas for potential distribution of Gastrodia elata based on ecological niche models. Chin. J. Plant Ecol. 2017, 41, 770–778. [Google Scholar]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L.J.E. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2010, 29, 773–785. [Google Scholar] [CrossRef]
- Mamun, M.; Kim, S.; An, K.G. Distribution pattern prediction of an invasive alien species largemouth bass using a maximum entropy model (MaxEnt) in the Korean peninsula. J. Asia-Pac. Biodivers. 2018, 11, 516–524. [Google Scholar] [CrossRef]
- Qin, A.; Jin, K.; Batsaikhan, M.-E.; Nyamjav, J.; Li, G.; Li, J.; Xue, Y.; Sun, G.; Wu, L.; Indree, T. Predicting the current and future suitable habitats of the main dietary plants of the Gobi Bear using MaxEnt modeling. Glob. Ecol. Conserv. 2020, 22, e01032. [Google Scholar] [CrossRef]
- West, A.M.; Kumar, S.; Brown, C.S.; Stohlgren, T.J.; Bromberg, J. Field validation of an invasive species Maxent model. Ecol. Inform. 2016, 36, 126–134. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Zhu, G.P.; Liu, Q.; Gao, Y.B. Improving ecological niche model transferability to predict the potential distribution of invasive exotic species. Biodivers. Sci. 2014, 22, 223–230. [Google Scholar]
- Zhang, H.; Song, J.; Zhao, H.; Li, M.; Han, W. Predicting the Distribution of the Invasive Species Leptocybe invasa: Combining MaxEnt and Geodetector Models. Insects 2021, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- Swets, J. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Gong, W.R.; Hu, B.S.; Zhou, G.L.; Bao, Y.X.; Liu, F.Q. Potential establishment areas of Solenopsis invicta in China: A prediction based on GIS. Chin. J. Appl. Ecol. 2006, 17, 2093–2097. [Google Scholar]
- Shen, W.J.; Wang, Y.N.; Wan, F.H. Application of Analogy Deviation to Predict Potential Distribution of Solenopsis invicta Buren (Hymenoptera: Formicidae) and Its Invasion Probability in China. Sci. Agric. Sin. 2008, 41, 1673–1683. [Google Scholar]
- Zhu, G.P.; Qiao, H.J. Effect of the Maxent model’s complexity on the prediction of species potential distributions. Biodivers. Sci. 2016, 24, 1189–1196. [Google Scholar] [CrossRef] [Green Version]
- Lei, Y.; Jaleel, W.; Khan, M.A.; Shahzad, M.F.; Khan, K.A. Effect of constant and fluctuating temperature on the circadian foraging rhythm of the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae). Saudi J. Biol. Sci. 2020, 28, 64–72. [Google Scholar] [CrossRef]
- Morrill, W.L. Dispersal of red imported fire ants by water. Fla. Entomol. 1974, 57, 39–42. [Google Scholar] [CrossRef]
- Markin, G.P.; Dillier, J.H.; Collins, H.L. Growth and Development of Colonies of the Red Imported Fire Ant, Solenopsis invicta. Ann. Entomol. Soc. Am. 1973, 66, 803–808. [Google Scholar] [CrossRef]
- Boulangeat, I.; Gravel, D.; Thuiller, W. Accounting for dispersal and biotic interactions to disentangle the drivers of species distributions and their abundances. Ecol. Lett. 2012, 15, 584–593. [Google Scholar] [CrossRef]
- Qiao, H.J.; Hu, J.H.; Huang, J.H. Theoretical basis, future directions, and challenges for ecological niche models. Sci. Sin. 2013, 43, 915–927. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Description | Whether to Use for Modeling |
|---|---|---|
| Bio1 | Annual Mean Temperature (°C) | Yes |
| Bio2 | Mean Diurnal Range (Mean of monthly (max temp–min temp)) (°C) | No |
| Bio3 | Isothermality (BIO2/BIO7) (×100) | No |
| Bio4 | Temperature Seasonality (standard deviation ×100) | No |
| Bio5 | Max Temperature of Warmest Month (°C) | No |
| Bio6 | Min Temperature of Coldest Month (°C) | Yes |
| Bio7 | Temperature Annual Range (BIO5-BIO6) (°C) | Yes |
| Bio8 | Mean Temperature of Wettest Quarter (°C) | No |
| Bio9 | Mean Temperature of Driest Quarter (°C) | Yes |
| Bio10 | Mean Temperature of Warmest Quarter (°C) | No |
| Bio11 | Mean Temperature of Coldest Quarter (°C) | No |
| Bio12 | Annual precipitation (mm) | Yes |
| Bio13 | Precipitation of Wettest Month (mm) | No |
| Bio14 | Coefficient of Variation of Precipitation Seasonality | Yes |
| Bio15 | Precipitation Seasonality (Coefficient of Variation) | No |
| Bio16 | Precipitation of Wettest Quarter (mm) | No |
| Bio17 | Precipitation of Driest Quarter (mm) | No |
| Bio18 | Precipitation of Warmest Quarter (mm) | No |
| Bio19 | Precipitation of Coldest Quarter (mm) | No |
| NDVI | Normalized Vegetation Index | Yes |
| Altitude | Altitude (m) | Yes |
| Slope | Slope (°) | No |
| Aspect | Aspect | No |
| LUCC | Land Use and Land Cover Change | Yes |
| Period | Highly Suitable | Moderately Suitable | Poorly Suitable | Total Suitable |
|---|---|---|---|---|
| Current | 3.24 | 10.66 | 67.47 | 81.37 |
| 2030s, SSPs1-2.6 | 3.87 | 14.44 | 84.54 | 98.98 |
| 2030s, SSPs2-4.5 | 4.00 | 15.40 | 88.78 | 108.18 |
| 2030s, SSPs5-8.5 | 4.26 | 16.05 | 92.83 | 113.14 |
| 2050s, SSPs1-2.6 | 4.29 | 16.31 | 92.62 | 113.22 |
| 2050s, SSPs2-4.5 | 5.02 | 19.42 | 104.68 | 129.12 |
| 2050s, SSPs5-8.5 | 5.46 | 20.94 | 110.31 | 136.71 |
| Period | Loss | Gain | Unchanged |
|---|---|---|---|
| 2030s, SSPs1-2.6 | 0.08 | 19.32 | 72.96 |
| 2030s, SSPs2-3.5 | 0.04 | 23.89 | 73.00 |
| 2030s, SSPs5-8.5 | 0.05 | 28.60 | 72.99 |
| 2050s, SSPs1-2.6 | 0.14 | 28.42 | 72.89 |
| 2050s, SSPs2-3.5 | 0.11 | 42.91 | 72.93 |
| 2050s, SSPs5-8.5 | 0.01 | 49.44 | 73.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Zhang, H.; Li, M.; Han, W.; Yin, Y.; Lei, J. Prediction of Spatiotemporal Invasive Risk of the Red Import Fire Ant, Solenopsis invicta (Hymenoptera: Formicidae), in China. Insects 2021, 12, 874. https://doi.org/10.3390/insects12100874
Song J, Zhang H, Li M, Han W, Yin Y, Lei J. Prediction of Spatiotemporal Invasive Risk of the Red Import Fire Ant, Solenopsis invicta (Hymenoptera: Formicidae), in China. Insects. 2021; 12(10):874. https://doi.org/10.3390/insects12100874
Chicago/Turabian StyleSong, Jinyue, Hua Zhang, Ming Li, Wuhong Han, Yuxin Yin, and Jinping Lei. 2021. "Prediction of Spatiotemporal Invasive Risk of the Red Import Fire Ant, Solenopsis invicta (Hymenoptera: Formicidae), in China" Insects 12, no. 10: 874. https://doi.org/10.3390/insects12100874
APA StyleSong, J., Zhang, H., Li, M., Han, W., Yin, Y., & Lei, J. (2021). Prediction of Spatiotemporal Invasive Risk of the Red Import Fire Ant, Solenopsis invicta (Hymenoptera: Formicidae), in China. Insects, 12(10), 874. https://doi.org/10.3390/insects12100874