
Different Host Plants Distinctly Influence the Feeding Ability of the Brown Citrus Aphid Toxoptera citricida

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Plants and Maintenance of Aphid Colonies
2.2. EPG Recording and Analysis
2.3. Population Development
2.4. Data Analysis
3. Results
3.1. Overall Characteristics of EPG Recordings
3.2. Pathway Activities
3.3. Phloem and Xylem Activities
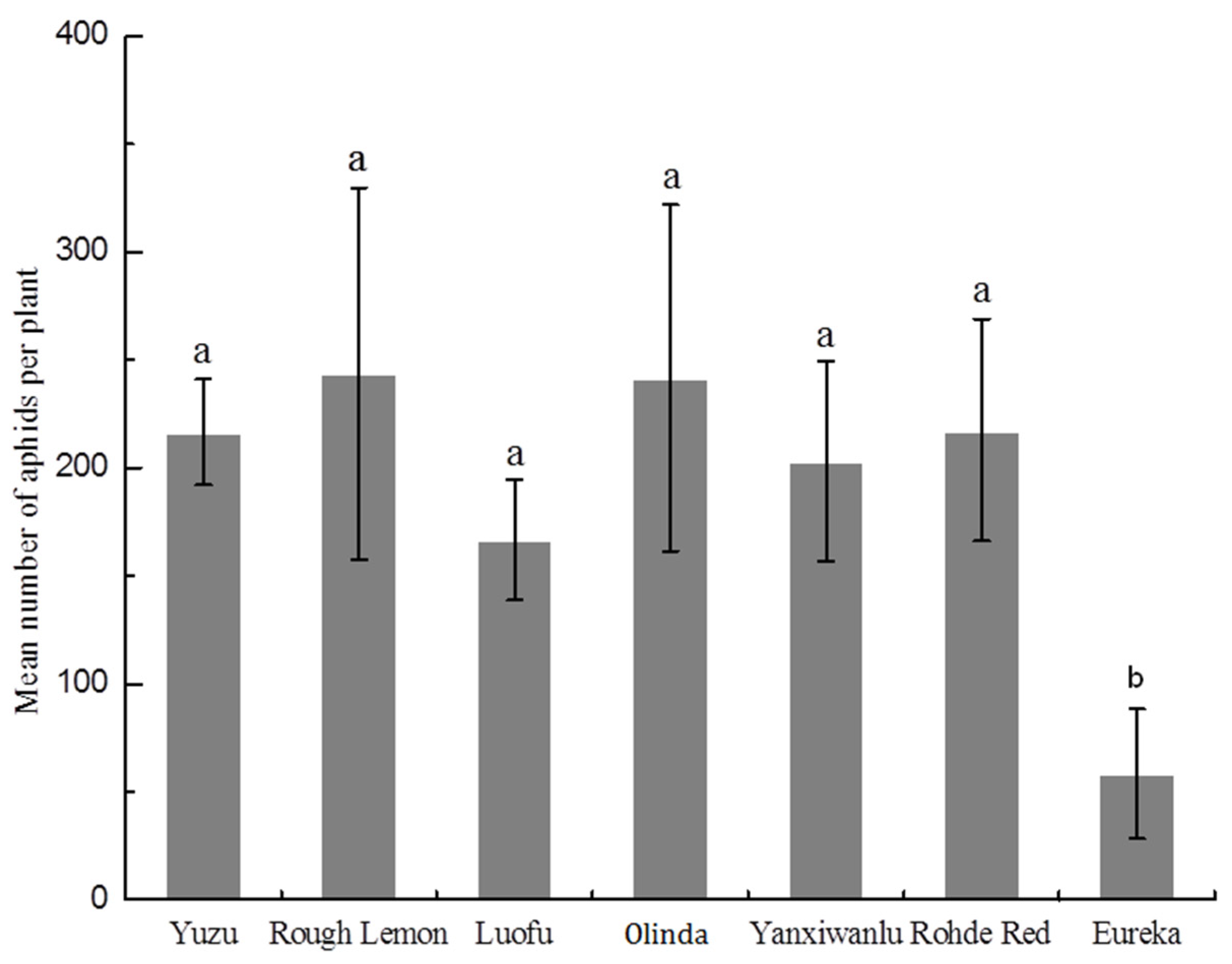
3.4. Population Development
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, Z.M. Current status of research on citrus pests in China. Chin. J. Appl. Entomol. 2000, 37, 110–116. [Google Scholar]
- Talon, M.; Gmitter, F.G. Citrus genomics. Int J. Plant. Genom. 2008, 2008, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Brlansky, R.H.; Damsteegt, V.D.; Howd, D.S.; Roy, A. Molecular analyses of Citrus tristeza virus subisolates separated by aphid transmission. Plant. Dis. 2003, 87, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, D.G.; Albrigo, L.G. Estimating the relative abundance of flush shoots in citrus with implications on monitoring insects associated with flush. HortScience 2007, 42, 364–368. [Google Scholar] [CrossRef] [Green Version]
- Yokomi, R.K.; Lastra, R.; Stoetzel, M.B.; Damsteegt, V.D.; Lee, R.F.; Garnsey, S.M.; Gottwald, T.R.; Rocha-Pena, M.A.; Niblett, C.L. Establishment of the brown citrus aphid (Homoptera: Aphididae) in central America and the Caribbean Basin and transmission of Citrus tristeza virus. J. Econ. Entomol. 1994, 87, 1078–1085. [Google Scholar] [CrossRef]
- Brlansky, R.H.; Roy, A.; Damsteegt, V.D. Stem-pitting Citrus tristeza virus predominantly transmitted by the brown citrus aphid from mixed infections containing non-stem-pitting and stem-pitting isolates. Plant. Dis. 2011, 95, 913–920. [Google Scholar] [CrossRef] [Green Version]
- Moreno, P.; Ambros, S.; Albiach-Marti, M.R.; Guerri, J.; Pena, L. Citrus tristeza virus: A pathogen that changed the course of the citrus industry. Mol. Plant. Pathol. 2008, 9, 251–268. [Google Scholar] [CrossRef]
- Tatineni, S.; Dawson, W.O. Enhancement or attenuation of disease by deletion of genes from Citrus tristeza virus. J. Virol. 2012, 86, 7850–7857. [Google Scholar] [CrossRef] [Green Version]
- Dawson, W.O.; Garnsey, S.M.; Tatineni, S.; Folimonova, S.Y.; Harper, S.J.; Gowda, S. Citrus tristeza virus-host interactions. Front. Microbiol. 2013, 4, 1–10. [Google Scholar] [CrossRef] [Green Version]
- He, Y.Q.; Chen, W.L.; Lu, Z.Y.; Zhou, C.Y.; Li, Z.A.; Wang, X.F.; Li, T.S. EPG analysis of the Toxopter acitricida on healthy and CTV -infected citrus. J. Mt. Agric. Biol. 2014, 33, 36–39. [Google Scholar]
- Tang, Y.Q.; Weathersbee, A.A.; Mayer, R.T. Effect of neem seed extract on the brown citrus aphid (Homoptera: Aphididae) and its parasitoid Lysiphlebus testaceipes (Hymenoptera: Aphidiidae). Environ. Entomol. 2002, 31, 172–176. [Google Scholar] [CrossRef] [Green Version]
- Smaili, M.C.; El Ghadraoui, L.; Gaboun, F.; Benkirane, R.; Blenzar, A. Impact of some alternative methods to chemical control in controlling aphids (Hemiptera: Sternorrhyncha) and their side effects on natural enemies on young Moroccan citrus groves. Phytoparasitica 2014, 42, 421–436. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Plant Health (PLH); Jeger, M.; Bragard, C.; Caffier, D.; Candresse, T.; Chatzivassiliou, E.; Dehnen-Schmutz, K.; Gilioli, G.; Gregoire, J.C.; Jaques Miret, J.A.; et al. Pest categorization of Toxoptera citricida. EFSA J. 2018, 16, 1–22. [Google Scholar]
- Zhou, H.; Du, L.; Wan, F.H.; Zhou, H. Comparative analysis of stylet penetration behaviors of Eriosoma lanigerum (Hemiptera: Aphididae) on main apple cultivars in China. J. Econ. Entomol. 2020, 113, 1761–1767. [Google Scholar]
- Pobozniak, M.; Gaborska, M.; Wojtowicz, T. Resistance and tolerance of ten carrot cultivars to the hawthorn-carrot aphid, Dysaphis crataegi Kalt., in Poland. PLoS ONE 2021, 16, e0247978. [Google Scholar] [CrossRef] [PubMed]
- Bhumannavar, B.S.; Singh, S.P.; Sulladmath, V.V. Field evaluation of citrus germplasm for resistance to the black aphid, Toxoptera aurantii (Boy.) under tropical humid South Indian conditions. Insect Sci. Applic. 1989, 10, 81–88. [Google Scholar] [CrossRef]
- Tsai, J.H. Development, survivorship, and reproduction of Toxoptera citricida (Kirkaldy) (Homoptera: Aphididae) on eight host plants. Environ. Entomol. 1998, 27, 1190–1195. [Google Scholar] [CrossRef]
- Lu, Z.Y.; Chen, W.L.; He, Y.Q.; Zhou, C.Y.; Wang, X.F.; Li, T.S. Difference in feeding behavior of brown citrus aphid Toxoptera citricida on five species of orange seedlings. J. Plant. Protect. 2014, 41, 637–638. [Google Scholar]
- Kimmins, F.M.; Tjallingii, W.F. Ultrastructure of sieve element penetration by aphid stylets during electrical recording. Exp. Appl. 1985, 39, 135–141. [Google Scholar] [CrossRef]
- Tjallingii, W.F.; Hogen Esch, T. Fine structure of aphid stylet routes in plant tissues in correlation with EPG signals. Physiol. Entomol. 1993, 18, 317–328. [Google Scholar] [CrossRef]
- Prado, E.; Tjallingii, W.F. Aphids activities during sieve element punctures. Exp. Appl. 1994, 72, 157–165. [Google Scholar] [CrossRef]
- Chen, J.Q.; Martín, B.; Rahbé, Y.; Fereres, A. Early intracellular punctures by two aphid species on near-isogenic melon lines with and without the virus aphid transmission (Vat) resistance gene. Eur. J. Plant Pathol. 1997, 103, 521–536. [Google Scholar] [CrossRef]
- Tjallingii, W.F.; Gabrys, B. Anomalous stylet punctures of phloem sieve elements by aphids. Exp. Appl. 1999, 91, 97–103. [Google Scholar] [CrossRef]
- Jimenez, J.; Garzo, E.; Alba-Tercedor, J.; Moreno, A.; Fereres, A.; Walker, G.P. The phloem pd: A distinctive brief sieve element stylet puncture prior to sieve element phase of aphid feeding behavior. Arthropod Plant Interact. 2019, 14, 67–78. [Google Scholar] [CrossRef]
- Van Helden, M.; Tjallingii, W.F. Tissue localization of lettuce resistance to the aphid Nasonovia ribisnigri using electrical penetration graphs. Exp. Appl. 1993, 68, 269–278. [Google Scholar] [CrossRef]
- Zhao, M.; Tian, T.W.; Li, W.Z.; Luo, M.H.; Guo, X.R.; Yan, F.M. Comparative analysis of Rhopalosiphum maidis feeding behaviors on eight maize hybrids (inbreds). Sci. Agr. Sinica 2015, 48, 1538–1547. [Google Scholar]
- Peng, H.C.; Walker, G.P. Sieve element occlusion provides resistance against Aphis gossypii in TGR-1551 melons. Insect Sci. 2018, 1, 33–48. [Google Scholar] [CrossRef]
- Garzo, E.; Soria, M.L.; Gómez-Guillamon, M.L.; Fereres, A. Feeding behaviour of Aphis gossypii on resistant accessions of different melon genotypes (Cucumis melo). Phytoparasitica 2002, 30, 129–140. [Google Scholar] [CrossRef]
- Liang, L.Y.; Liu, L.F.; Yu, X.P.; Han, B.Y. Evaluation of the resistance of different tea cultivars to tea aphids by EPG technique. J. Integr. Agr. 2012, 11, 2028–2034. [Google Scholar] [CrossRef]
- Alvarez, A.E.; Broglia, V.G.; Alberti D’Amato, A.M.; Wouters, D.; van der Vossen, E.; Garzo, E.; Tjallingii, W.F.; Dicke, M.; Vosman, B. Comparative analysis of Solanum stoloniferum responses to probing by the green peach aphid Myzus persicae and the potato aphid Macrosiphum euphorbiae. Insect Sci. 2012, 20, 207–227. [Google Scholar] [CrossRef]
- Sun, M.; Voorrps, R.E.; Steenhuis-Broers, G.; van’t Westende, W.; Vosman, B. Reduced phloem uptake of Myzus persicae on an aphid resistant pepper accession. BMC Plant Biol. 2018, 18, 138. [Google Scholar] [CrossRef]
- Guo, J.G.; Yuan, W.N.; Zhou, T.W.; Zhang, X.R.; He, C.G.; Zhao, G.Q.; Zhang, Z.W. Feeding behavior of biotype E of the greenbug, Schizaphis graminum (Hemiptera:Aphididae) on oats with different phenotypic resistance. Acta Entomol. Sinica 2017, 60, 1315–1323. [Google Scholar]
- Jiang, Y.X.; Chang, W.J.; Zhan, Y.D.; Liu, Z.; Liu, Y. Investigation on the resistance of wheat germplasm resources to aphid based on fuzzy recognition and electrical penetration graph (EPG) techniques. Chin. J. Appl. Ecol. 2020, 31, 3248–3254. [Google Scholar]
- Tjallingii, W.F. Electronic recording of penetration behaviour by aphids. Entomol. Exp. Appl. 1978, 24, 721–730. [Google Scholar] [CrossRef]
- Spiller, N.J.; Koenders, L.; Tjallingii, W.F. Xylem ingestion by aphids—A strategy for maintaining water balance. Entomol. Exp. Appl. 2011, 55, 101–104. [Google Scholar] [CrossRef]
- Garzo, E.; Moreno, A.; Plaza, M.; Fereres, A. Feeding behavior and virus-transmission ability of insect vectors exposed to systemic insecticides. Plants 2021, 9, 895. [Google Scholar] [CrossRef]
- Sarria, E.; Cid, M.; Garzo, E.; Fereres, A. Excel workbook for automatic parameter calculation of EPG data. Comput. Electron. Agr. 2009, 67, 35–42. [Google Scholar] [CrossRef]
- Schwarzkopf, A.; Rosenberger, D.; Niebergall, M.; Gershenzon, J.; Kunert, G. To feed or not to feed: Plant factors located in the epidermis, mesophyll, and sieve elements influence pea aphid’s ability to feed on Legume species. PLoS ONE 2013, 8, e75298. [Google Scholar]
- Zhao, R.N.; He, Y.Q.; Lu, Z.Y.; Chen, W.L.; Zhou, C.Y.; Wang, X.F.; Li, T.S. An analysis of the feeding behavior of three stages of Toxoptera citricida by DC electrical penetration graph waveforms. Entomol. Exp. Appl. 2019, 167, 370–376. [Google Scholar] [CrossRef] [Green Version]
- Powell, C.A.; Burton, M.S.; Pelosi, R.R.; Rundell, P.A.; Ritenour, M.A.; Bullock, R.C. Six-year evaluation of brown citrus and spirea aphid populations in a citrus grove and the effects of insecticides on these populations. HortScience 2006, 41, 688–690. [Google Scholar] [CrossRef]
- Zehnder, C.B.; Hunter, M.D. Effects of nitrogen deposition on the interaction between an aphid and its host plant. Ecol. Entomol. 2008, 33, 24–30. [Google Scholar] [CrossRef]
- Ford, E.S. Anatomy and histology of the eureka lemon. Bot. Gaz. 1942, 104, 288–305. [Google Scholar] [CrossRef]
- Wagner, G.J.; Wang, E.; Shepherd, R.W. New approaches for studying and exploiting an old protuberance, the plant trichome. Ann. Bot. 2004, 93, 3–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, A.T.; Gurr, G.M.; Mcgrath, D.; Nicol, H.I.; Martin, P.M. Trichomes of Lycopersicon spp. and their effect on Myzus persicae (Sulzer) (Hemiptera: Aphididae). Aust. J. Entomol. 2003, 42, 373–378. [Google Scholar] [CrossRef]
- Rakha, M.; Hanson, P.; Ramasamy, S. Identification of resistance to Bemisia tabaci Genn. in closely related wild relatives of cultivated tomato based on trichome type analysis and choice and no-choice assays. Genet. Resour. Crop. Evol. 2017, 64, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Lopez, M.J.; Garzo, E.; Bonani, J.P.; Fernandez-Munoz, R.; Moriones, E.; Fereres, A. Acylsucrose-producing tomato plants forces Bemisia tabaci to shift its preferred settling and feeding site. PLoS ONE 2012, 7, e33064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dardouri, T.; Gomez, L.; Ameline, A.; Costagliola, G.; Schoeny, A.; Gautier, H. Non-host volatiles disturb the feeding behavior and reduce the fecundity of the green peach aphid, Myzus persicae. Pest. Manag. Sci. 2021, 77, 1705–1713. [Google Scholar] [CrossRef]
- Walker, G.P. The role of leaf cuticle in leaf age preference by bayberry whitefly (Homoptera: Aleyrodidae) on lemon. Ann. Entomol. Soc. Am. 1988, 81, 365–369. [Google Scholar] [CrossRef]
- Prado, E.; Tjallingii, W.F. Effects of previous plant infestation on sieve element acceptance by two aphids. Entomol. Exp. Appl. 1997, 82, 189–200. [Google Scholar] [CrossRef]
- Nalam, V.J.; Han, J.L.; Pitt, W.J.; Acharya, S.R.; Nachappa, P. Location, location, location: Feeding site affects aphid performance by altering access and quality of nutrients. PLoS ONE 2021, 16, e0245380. [Google Scholar] [CrossRef] [PubMed]
- Shugart, H.; Ebert, T.; Gmitter, F.; Rogers, F. The power of electropenetrography in enhancing our understanding of host plant-vector interactions. Insects 2019, 10, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wensler, R.J.; Filshie, B.K. Gustatory sense organs in the food canal of aphids. J. Morphol. 1969, 129, 473–492. [Google Scholar] [CrossRef]
- Kordan, B.; Stec, K.; Slominski, P.; Giertych, M.J.; Wróblewska-Kurdyk, A.; Gabrys, B. Susceptibility of forage legumes to infestation by the pea aphid Acyrthosiphon pisum (Harris) (Hemiptera: Aphididae). Crop. Pasture Sci. 2018, 69, 775–784. [Google Scholar] [CrossRef]
- Medina-Ortega, K.J.; Walker, G.P. Does aphid salivation affect phloem sieve element occlusion in vivo? J. Exp. Bot. 2013, 64, 5525–5535. [Google Scholar] [CrossRef] [PubMed]
- Caillaud, C.M.; Niemeyer, H.M. Possible involvement of the phloem sealing system in the acceptance of a plant as host by an aphid. Experientia 1996, 52, 927–931. [Google Scholar] [CrossRef]
- Miao, J.; Han, B.Y.; Zhang, Q.H. Probing behavior of Empoasca vitis (Homoptera: Cicadellidae) on resistance and susceptible cultivars of tea plants. J. Insect Sci. 2014, 14, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, J.C.K.; Perry, K.L. Transmission of plant viruses by aphid vectors. Mol. Plant. Pathol. 2004, 5, 505–511. [Google Scholar] [CrossRef]
- Rodríguez-López, M.J.; Garzo, E.; Fereres, A.; Fernández-Muñoz, R.; Bonani, J.P.; Moriones, E. Whitefly resistance traits derived from the wild tomato Solanum pimpinellifolium affect the preference and feeding behavior of Bemisia tabaci and reduce the spread of Tomato yellow leaf curl virus. Phytopathology 2011, 101, 1191–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paprocka, M.; Gliszczyńska, A.; Dancewicz, K.; Gabryś, B. Novel hydroxy- and epoxy-cis-jasmone and dihydro jasmone derivatives affect the foraging activity of the peach potato aphid Myzus persicae (Sulzer) (Homoptera: Aphididae). Molecules 2018, 23, 2362. [Google Scholar] [CrossRef] [PubMed]
- Escudero-Martinez, C.; Leybourne, D.J.; Bos, J.I.B. Plant resistance in different cell layers affects aphid probing and feeding behaviour during non-host and poor-host interactions. B Entomol. Res. 2021, 111, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Lucini, T.; Panizzi, A.R.; Bueno, A.D.F. Evaluating resistance of the soybean block technology cultivars to the Neotropical brown stink bug, Euschistus heros (F.). J. Insect Physiol. 2021, 131, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.P.; Zhan, H.X.; Wang, Y.L.; Hou, S.M. How cabbage aphids Brevicoryne brassicae (L.) make a choice to feed on Brassica napus cultivars. Insects 2019, 10, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
| Variables/ per Insect | Host Plants | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Yuzu | Rough Lemon | Luofu | Olinda | Yanxiwanlu | Rohde Red | Eureka | H | P | |
| Probing duration | 343.40 (306.47–350.62)a | 319.32 (257.64–334.59)a | 285.24 (265.18–356.57)a | 305.42 (228.82–335.80)a | 310.31 (277.87–336.36)a | 306.37 (273.98–330.82)a | 281.08 (241.45–351.41)a | 13.04 | 0.042 |
| Number of probes | 5.00 (2.50–10.00)c | 9.00 (6.00–13.00)abc | 7.50 (2.75–11.25)bc | 15.00 (7.00–20.00)ab | 9.00 (4.00–13.50)abc | 15.00 (11.50–22.00)a | 12.00 (5.00–21.00)abc | 26.86 | 0.000 |
| Probing duration per probe | 68.81 (28.13–145.45)a | 35.48 (18.65–58.16)ab | 38.12 (24.63–133.75)ab | 16.35 (12.90–43.99)b | 33.40 (17.44–81.44)ab | 20.96 (13.01–28.72)b | 22.48 (12.95–70.28)ab | 23.39 | 0.001 |
| Variables | Host Plants | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Yuzu | Rough Lemon | Luofu | Olinda | Yanxiwanlu | Rohde Red | Eureka | H | P | |
| Time to 1st probe (s) | 0.00 (0.00–80.69)b | 51.99 (35.11–83.30)a | 0.00 (0.00–97.24)b | 20.76 (0.00–43.51)ab | 0.00 (0.00–25.10)b | 0.00 (0.00–4.94)b | 16.46 (0.00–59.07)ab | 29.36 | 0.000 |
| Number of short probes (C < 3 min) | 4.00 (1.00–5.50)b | 6.00 (3.00–8.00)ab | 2.50 (0.00–5.75)ab | 8.00 (4.00–11.00)ab | 4.00 (1.50–7.50)ab | 7.00 (3.50–13.50)a | 5.00 (2.00–11.00)ab | 17.89 | 0.007 |
| Number of probes to the 1st E1 | 5.00 (2.00–8.00)b | 9.00 (5.00–11.50)ab | 5.50 (1.75–11.25)ab | 11.00 (7.00–19.00)a | 7.00 (4.00–9.00)ab | 12.00 (9.00–17.50)a | 7.00 (5.00–19.00)ab | 26.74 | 0.000 |
| Time from 1st probe to 1st E1(min) | 65.69 (31.05–108.68)b | 118.47 (79.65–192.00)ab | 126.45 (47.80–242.36)ab | 144.90 (83.30–239.03)ab | 116.83 (89.44–191.86)ab | 108.69 (76.79–221.82)ab | 161.47 (124.66–319.61)a | 14.57 | 0.024 |
| Time from 1st probe to 1st E2(min) | 66.61 (32.32–111.02)b | 119.14 (80.56–207.55)b | 166.22 (49.45–295.14)ab | 146.04 (84.30–249.45)ab | 121.49 (90.80–216.76)ab | 109.42 (77.85–222.89)b | 357.55 (239.46–359.95)a | 27.93 | 0.000 |
| % E2>10min in E2 | 100.00 (100.00–100.00)a | 100.00 (100.00–100.00)a | 100.00 (83.33–100.00)a | 100.00 (100.00–100.00)a | 100.00 (100.00–100.00)a | 100.00 (100.00–100.00)a | 12.50 (0.00–50.00)b | 26.92 | 0.000 |
| Variables | Host Plants | ppw | NWI | P | WDI | P | WDEI | P |
|---|---|---|---|---|---|---|---|---|
| np | Yuzu | 25/25 | 5.00 (2.00–9.50)c | 0.00 | 16.60 (9.38–53.53)a | 0.04 | 2.99 (2.25–53.53)a | 0.36 |
| Rough Lemon | 21/21 | 9.00 (6.00–13.50)abc | 40.68 (25.41–102.36)a | 4.11 (2.33–8.04)a | ||||
| Luofu | 14/15 | 7.00 (1.75–11.00)bc | 74.76 (3.43–94.82)a | 5.72 (1.83–10.50)a | ||||
| Olinda | 24/24 | 15.00 (6.00–20.00)ab | 54.58 (24.20–131.18)a | 4.84 (1.77–8.40)a | ||||
| Yanxiwanlu | 25/25 | 8.00 (4.00–13.00)abc | 49.69 (23.64–82.14)a | 5.06 (3.76–8.50)a | ||||
| Rohde Red | 25/25 | 15.00 (11.00–21.00)a | 53.63 (29.18–86.02)a | 3.32 (2.40–5.08)a | ||||
| Eureka | 15/15 | 12.00 (4.00–21.00)abc | 78.92 (8.59–118.55)a | 4.19 (2.66–5.67)a | ||||
| C | Yuzu | 25/25 | 6.00 (2.50–10.50)c | 0.00 | 66.69 (42.91–109.78)b | 0.00 | 14.05 (8.75–19.15) ab | 0.00 |
| Rough Lemon | 21/21 | 10.00 (6.00–13.50)abc | 71.65 (58.54–154.86)b | 9. 85(7.09–11.61)b | ||||
| Luofu | 15/15 | 7.50 (3.75–12.00)bc | 109.8 8(47.97–207.63)ab | 18.33 (11.30–24.28)a | ||||
| Olinda | 24/24 | 18.00 (7.00–22.00)ab | 131.33 (64.03–162.00)ab | 9.54 (7.08–11.90)b | ||||
| Yanxiwanlu | 25/25 | 9.00 (4.50–14.00)abc | 114.03 (87.71–189.04)ab | 14.25 (9.00–19.38)ab | ||||
| Rohde Red | 25/25 | 17.00 (11.50–23.50)a | 158.43 (99.55–212.91)a | 9.32 (6.65–12.66)b | ||||
| Eureka | 15/15 | 13.00 (7.00–21.00)abc | 206.55 (172.41–270.09)a | 14.14 (10.82–19.59)ab | ||||
| pd | Yuzu | 25/25 | 62.00 (43.5–113.5)c | 0.00 | 482.39 (277.79–698.16)bc | 0.00 | 6.39 (6.00–7.30)ab | 0.00 |
| Rough Lemon | 21/21 | 67.00 (51.50–92.50)c | 357.91(287.64–514.06)c | 5.80 (5.52–6.02)c | ||||
| Luofu | 15/15 | 66.50 (42.25–154.00)bc | 472.08 (299.03–1014.46)a | 6.52 (6.25–7.16)a | ||||
| Olinda | 24/24 | 126.00 (69.00–155.00)abc | 676.63 (431.70–806.95)a | 5.83 (5.34–6.48)b | ||||
| Yanxiwanlu | 25/25 | 98.00 (79.50–146.00)abc | 684.48 (490.46–913.77)a | 6.14 (5.87–6.65)ab | ||||
| Rohde Red | 25/25 | 130.00 (92.50–186.00)ab | 827.57 (567.68–1086.85)ab | 6.18 (5.84–6.50)ab | ||||
| Eureka | 15/15 | 175.00 (145.00–248.00)a | 1109.70 (747.83–1222.02)a | 5.67 (5.12–6.42)c | ||||
| E1 | Yuzu | 24/25 | 1.00 (1.00–2.00)a | 0.51 | 1.62 (0.96–2.53)ab | 0.01 | 1.06 (0.90–1.75)ab | 0.00 |
| Rough Lemon | 18/21 | 1.00 (1.00–1.00)a | 0.97 (0.84–1.46)b | 0.90 (0.79–1.15)c | ||||
| Luofu | 15/15 | 1.00 (1.00–1.25)a | 2.55 (1.58–5.19)ab | 2.00 (1.45–4.38)a | ||||
| Olinda | 22/24 | 1.00 (1.00–2.00)a | 1.84 (1.03–3.12)ab | 1.16(1.00–2.66)ab | ||||
| Yanxiwanlu | 21/25 | 1.00 (1.00–3.00)a | 2.67 (0.79–6.30)ab | 1.33 (0.79–3.03)ab | ||||
| Rohde Red | 21/25 | 1.00 (1.00–3.00)a | 1.83 (0.87–5.32)ab | 1.15 (0.74–2.27)ab | ||||
| Eureka | 12/15 | 2.00 (1.00–3.00)a | 10.49 (1.76–50.69)a | 5.24 (1.76–10.53)a | ||||
| E2 | Yuzu | 24/25 | 1.00 (1.00–2.00)a | 0.14 | 264.60 (183.13–295.85)a | 0.00 | 241.95 (80.52–295.86)a | 0.00 |
| Rough Lemon | 18/21 | 1.00 (1.00–1.00)a | 220.73 (90.09–278.72)a | 211.38 (81.39–273.13)a | ||||
| Luofu | 14/15 | 1.00 (1.00–1.00)a | 151.31 (62.36–304.42)a | 128.64 (42.39–283.11)a | ||||
| Olinda | 22/24 | 1.00 (1.00–2.00)a | 169.26 (67.28–244.60)a | 119.29 (30.09–235.39)a | ||||
| Yanxiwanlu | 21/25 | 1.00 (1.00–2.00)a | 150.50 (39.96–237.83)ab | 92.64 (18.10–195.31)ab | ||||
| Rohde Red | 21/25 | 1.00 (1.00–2.00)a | 148.48 (68.64–215.021)ab | 89.21 (26.63–180.96)ab | ||||
| Eureka | 7/15 | 0.00 (0.00–2.00)a | 0.00 (0.00–29.48)b | 0.00 (0.00–13.17)b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, R.; Wu, C.; He, Y.; Yu, C.; Liu, J.; Li, T.; Zhou, C.; Chen, W. Different Host Plants Distinctly Influence the Feeding Ability of the Brown Citrus Aphid Toxoptera citricida. Insects 2021, 12, 864. https://doi.org/10.3390/insects12100864
Zhao R, Wu C, He Y, Yu C, Liu J, Li T, Zhou C, Chen W. Different Host Plants Distinctly Influence the Feeding Ability of the Brown Citrus Aphid Toxoptera citricida. Insects. 2021; 12(10):864. https://doi.org/10.3390/insects12100864
Chicago/Turabian StyleZhao, Runa, Chengxu Wu, Yingqin He, Chun Yu, Jianfeng Liu, Taisheng Li, Changyong Zhou, and Wenlong Chen. 2021. "Different Host Plants Distinctly Influence the Feeding Ability of the Brown Citrus Aphid Toxoptera citricida" Insects 12, no. 10: 864. https://doi.org/10.3390/insects12100864
APA StyleZhao, R., Wu, C., He, Y., Yu, C., Liu, J., Li, T., Zhou, C., & Chen, W. (2021). Different Host Plants Distinctly Influence the Feeding Ability of the Brown Citrus Aphid Toxoptera citricida. Insects, 12(10), 864. https://doi.org/10.3390/insects12100864