
Characterizing the Spatiotemporal Distribution of Three Native Stink Bugs (Hemiptera: Pentatomidae) across an Agricultural Landscape


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Collection
2.2. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pilkay, G.L.; Reay-Jones, F.P.; Toews, M.D.; Greene, J.K.; Bridges, W.C. Spatial and temporal dynamics of stink bugs in southeastern farmscapes. J. Insect Sci. 2015, 15, 23. [Google Scholar]
- Reeves, R.; Greene, J.; Reay-Jones, F.; Toews, M.; Gerard, P. Effects of adjacent habitat on populations of stink bugs (Heteroptera: Pentatomidae) in cotton as part of a variable agricultural landscape in South Carolina. Environ. Entomol. 2010, 39, 1420–1427. [Google Scholar] [PubMed] [Green Version]
- Reisig, D.D.; Roe, M.; Dhammi, A. Dispersal pattern and dispersion of adult and nymph stink bugs (Hemiptera: Pentatomidae) in wheat and corn. Environ. Entomol. 2013, 42, 1184–1192. [Google Scholar]
- Tillman, P.G.; Northfield, T.D.; Mizell, R.F.; Riddle, T.C. Spatiotemporal patterns and dispersal of stink bugs (Heteroptera: Pentatomidae) in peanut-cotton farmscapes. Environ. Entomol. 2009, 38, 1038–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tillman, P.G.; Cottrell, T.E.; Mizell, R.F.; Kramer, E. Effect of field edges on dispersal and distribution of colonizing stink bugs across farmscapes of the Southeast USA. Bull. Entomol. Res. 2014, 104, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Babu, A.; Reisig, D.D.; Walgenbach, J.F.; Heiniger, R.W.; Eyerman, W. Influence of weed manipulation in field borders on brown stink bug (Hemiptera: Pentatomidae) densities and damage in field corn. Environ. Entomol. 2019, 48, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.A., Jr.; Sullivan, M. Overwintering habitats, spring emergence patterns, and winter mortality of some South Carolina Hemiptera. Environ. Entomol. 1981, 10, 409–414. [Google Scholar]
- Jones, W.A.; Sullivan, M. Role of host plants in population dynamics of stink bug pests of soybean in South Carolina. Environ. Entomol. 1982, 11, 867–875. [Google Scholar]
- Lee, D.-H.; Nielsen, A.L.; Leskey, T.C. Dispersal capacity and behavior of nymphal stages of Halyomorpha halys (Hemiptera: Pentatomidae) evaluated under laboratory and field conditions. J. Insect Behav. 2014, 27, 639–651. [Google Scholar]
- Bernon, G. Biology of Halyomorpha Halys, The Brown Marmorated Stink Bug (BMSB): Final Report; APHIS CPHST project T3P01; US Department of Agriculture: Washington, DC, USA, 2004.
- Funayama, K. Oviposition and development of Halyomorpha halys (Stäl) and Homalogonia obtusa (Walker)(Heteroptera: Pentatomidae) on apple [Molus pumila] trees. Jpn. J. Appl. Entomol. Zool. 2002, 46, 1–6. [Google Scholar]
- Nielsen, A.L.; Hamilton, G.C. Life history of the invasive species Halyomorpha halys (Hemiptera: Pentatomidae) in northeastern United States. Ann. Entomol. Soc. Am. 2009, 102, 608–616. [Google Scholar]
- Tillman, P. Likelihood of stink bugs colonizing crops: A case study in southeastern farmscapes. Environ. Entomol. 2013, 42, 438–444. [Google Scholar] [PubMed]
- Herbert, J.; Toews, M. Seasonal abundance and population structure of brown stink bug (Hemiptera: Pentatomidae) in farmscapes containing corn, cotton, peanut, and soybean. Ann. Entomol. Soc. Am. 2011, 104, 909–918. [Google Scholar]
- Olson, D.; Ruberson, J.; Andow, D. Effects on stink bugs of field edges adjacent to woodland. Agric. Ecosyst. Environ. 2012, 156, 94–98. [Google Scholar]
- Tillman, P.G.; Cottrell, T.E. Density and egg parasitism of stink bugs (Hemiptera: Pentatomidae) in elderberry and dispersal into crops. J. Insect Sci. 2016, 16, 10. [Google Scholar] [CrossRef] [Green Version]
- Tillman, P.G. Density and egg parasitism of stink bugs (Hemiptera: Pentatomidae) in mimosa. Fla. Entomol. 2019, 102, 227–230. [Google Scholar]
- Aldrich, J.R.; Hoffmann, M.P.; Kochansky, J.P.; Lusby, W.R.; Eger, J.E.; Payne, J.A. Identification and attractiveness of a major pheromone component for Nearctic Euschistus spp. stink bugs (Heteroptera: Pentatomidae). Environ. Entomol. 1991, 20, 477–483. [Google Scholar]
- Cottrell, T.E.; Yonce, C.E.; Wood, B.W. Seasonal occurrence and vertical distribution of Euschistus servus (Say) and Euschistus tristigmus (Say)(Hemiptera: Pentatomidae) in pecan orchards. J. Entomol. Sci. 2000, 35, 421–431. [Google Scholar]
- Johnson, D.; Lewis, B.; Mizell, R., III. Trapping brown stink bugs in peach. Horticultural studies 2001. Arkans. Agric. Exp. Stn. Res. Ser. 2002, 494, 19–23. [Google Scholar]
- Leskey, T.; Hogmire, H. Monitoring stink bugs (Hemiptera: Pentatomidae) in mid-Atlantic apple and peach orchards. J. Econ. Entomol. 2005, 98, 143–153. [Google Scholar]
- Mizell, R.; Tedders, W. A New Monitoring Method for Detection of the Stinkbug Complex in Pecan Orchards. Proc. Southeast. Pecan Grow. Assoc. 1995, 88, 36–40. [Google Scholar]
- Tillman, P.G.; Cottrell, T.E. Attraction of stink bug (Hemiptera: Pentatomidae) nymphs to Euschistus aggregation pheromone in the field. Fla. Entomol. 2016, 99, 678–682. [Google Scholar]
- Tillman, P.; Aldrich, J.; Khrimian, A.; Cottrell, T. Pheromone attraction and cross-attraction of Nezara, Acrosternum, and Euschistus spp. stink bugs (Heteroptera: Pentatomidae) in the field. Environ. Entomol. 2010, 39, 610–617. [Google Scholar] [PubMed] [Green Version]
- Khrimian, A.; Zhang, A.; Weber, D.C.; Ho, H.-Y.; Aldrich, J.R.; Vermillion, K.E.; Siegler, M.A.; Shirali, S.; Guzman, F.; Leskey, T.C. Discovery of the aggregation pheromone of the brown marmorated stink bug (Halyomorpha halys) through the creation of stereoisomeric libraries of 1-bisabolen-3-ols. J. Nat. Prod. 2014, 77, 1708–1717. [Google Scholar]
- Weber, D.C.; Walsh, G.C.; DiMeglio, A.S.; Athanas, M.M.; Leskey, T.C.; Khrimian, A. Attractiveness of harlequin bug, Murgantia histrionica, aggregation pheromone: Field response to isomers, ratios, and dose. J. Chem. Ecol. 2014, 40, 1251–1259. [Google Scholar] [PubMed] [Green Version]
- Cottrell, T.E.; Horton, D.L. Trap capture of brown and dusky stink bugs (Hemiptera: Pentatomidae) as affected by pheromone dosage in dispensers and dispenser source. J. Entomol. Sci. 2011, 46, 135–147. [Google Scholar]
- Cottrell, T.E. Improved trap capture of Euschistus servus and Euschistus tristigmus (Hemiptera: Pentatomidae) in pecan orchards. Fla. Entomol. 2001, 84, 731–732. [Google Scholar]
- McPherson, J.E.; McPherson, R. Stink Bugs of Economic Importance in America North of Mexico; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Borges, M.; Zhang, A.; Camp, M.J.; Aldrich, J.R. Adult diapause morph of the brown stink bug, Euschistus servus (Say)(Heteroptera: Pentatomidae). Neotrop. Entomol. 2001, 30, 179–182. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.N.; Winder, L.; Holland, J.M.; Alston, R.D. Red-blue plots for detecting clusters in count data. Ecol. Lett. 1999, 2, 106–113. [Google Scholar] [CrossRef]
- Gigot, C. Analyzing Plant Disease Epidemics with R Package Epiphy. 2018. Available online: https://cran.r-project.org/web/packages/epiphy/vignettes/epiphy.html (accessed on 21 September 2021).
- Winder, L.; Alexander, C.; Griffiths, G.; Holland, J.; Woolley, C.; Perry, J. Twenty years and counting with SADIE: Spatial Analysis by Distance Indices software and review of its adoption and use. Rethink. Ecol. 2019, 4, 1. [Google Scholar]
- He, F.; Jeske, D.R.; Grafton-Cardwell, E. Identifying high-density regions of pests within an orchard. Appl. Stoch. Models Bus. Ind. 2020, 36, 417–431. [Google Scholar]
- Reay-Jones, F. Spatial and temporal patterns of stink bugs (Hemiptera: Pentatomidae) in wheat. Environ. Entomol. 2014, 39, 944–955. [Google Scholar]
- Grabarczyk, E.E.; Olson, D.M.; Tillman, P.G.; Hodges, A.C.; Hodges, G.; Horton, D.L.; Cottrell, T.E. Spatiotemporal distribution of stink bugs (Hemiptera: Pentatomidae) in peach orchards and surrounding habitat. Fla. Entomol. 2021, 104, 27–35. [Google Scholar]
- Tillman, P.G. Composition and abundance of stink bugs (Heteroptera: Pentatomidae) in corn. Environ. Entomol. 2010, 39, 1765–1774. [Google Scholar] [PubMed]
- Olson, D.M.; Prescott, K.K.; Zeilinger, A.R.; Hou, S.; Coffin, A.W.; Smith, C.M.; Ruberson, J.R.; Andow, D.A. Landscape effects on reproduction of Euschistus servus (Hemiptera: Pentatomidae), a mobile, polyphagous, multivoltine arthropod herbivore. Environ. Entomol. 2018, 47, 660–668. [Google Scholar]
- Mizell, R., III; Tedders, W.; Yonce, C.; Aldrich, J. Stink bug monitoring—An update. Proc. Southeast. Pecan Grow. Assoc. 1997, 90, 50–52. [Google Scholar]
- Yonce, C.; Mizell, R., III. Stink bug trapping with a pheromone. Proc. Southeast. Pecan Grow. Assoc. 1997, 90, 54–56. [Google Scholar]
- Venugopal, P.D.; Coffey, P.L.; Dively, G.P.; Lamp, W.O. Adjacent habitat influence on stink bug (Hemiptera: Pentatomidae) densities and the associated damage at field corn and soybean edges. PLoS ONE 2014, 9, e109917. [Google Scholar] [CrossRef]
- Bergmann, E.J.; Venugopal, P.D.; Martinson, H.M.; Raupp, M.J.; Shrewsbury, P.M. Host plant use by the invasive Halyomorpha halys (Stål) on woody ornamental trees and shrubs. PLoS ONE 2016, 11, e0149975. [Google Scholar]
- Holthouse, M.C.; Spears, L.R.; Alston, D.G. Urban host plant utilisation by the invasive Halyomorpha halys (Stål)(Hemiptera, Pentatomidae) in northern Utah. NeoBiota 2021, 64, 87. [Google Scholar]
- Blackshaw, R.P.; Vernon, R.S. Spatiotemporal stability of two beetle populations in non-farmed habitats in an agricultural landscape. J. Appl. Ecol. 2006, 43, 680–689. [Google Scholar]
- Amaral, D.S.; Venzon, M.; dos Santos, H.H.; Sujii, E.R.; Schmidt, J.M.; Harwood, J.D. Non-crop plant communities conserve spider populations in chili pepper agroecosystems. Biol. Control. 2016, 103, 69–77. [Google Scholar]
- Pearce, S.; Zalucki, M.P. Do predators aggregate in response to pest density in agroecosystems? Assessing within-field spatial patterns. J. Appl. Ecol. 2006, 43, 128–140. [Google Scholar]
- Shrestha, G.; Rijal, J.P.; Reddy, G.V. Characterization of the spatial distribution of alfalfa weevil, Hypera postica, and its natural enemies, using geospatial models. Pest Manag. Sci. 2021, 77, 906–918. [Google Scholar]
- Winder, L.; Alexander, C.J.; Holland, J.M.; Woolley, C.; Perry, J.N. Modelling the dynamic spatio-temporal response of predators to transient prey patches in the field. Ecol. Lett. 2001, 4, 568–576. [Google Scholar] [CrossRef]
- Klick, J.; Yang, W.Q.; Walton, V.M.; Dalton, D.; Hagler, J.; Dreves, A.; Lee, J.; Bruck, D. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. J. Appl. Entomol. 2016, 140, 37–46. [Google Scholar]
- Conti, E.; Avila, G.; Barratt, B.; Cingolani, F.; Colazza, S.; Guarino, S.; Hoelmer, K.; Laumann, R.A.; Maistrello, L.; Martel, G. Biological control of invasive stink bugs: Review of global state and future prospects. Entomol. Exp. Appl. 2021, 169, 28–51. [Google Scholar]
- Tillman, P.; Khirmian, A.; Cottrell, T.; Lou, X.; Mizell, R., III; Johnson, C. Trap cropping systems and a physical barrier for suppression of stink bugs (Hemiptera: Pentatomidae) in cotton. J. Econ. Entomol. 2015, 108, 2324–2334. [Google Scholar] [CrossRef]
- Gordon, T.; Haseeb, M.; Kanga, L.; Legaspi, J. Potential of three trap crops in managing Nezara viridula (Hemiptera: Pentatomidae) on tomatoes in Florida. J. Econ. Entomol. 2017, 110, 2478–2482. [Google Scholar] [CrossRef] [PubMed]
- Mizell III, R.F.; Riddle, T.C.; Blount, A.S. Trap cropping system to suppress stink bugs in the southern coastal plain. Proc. Fla. State Hortic. Soc. 2008, 121, 377–382. [Google Scholar]
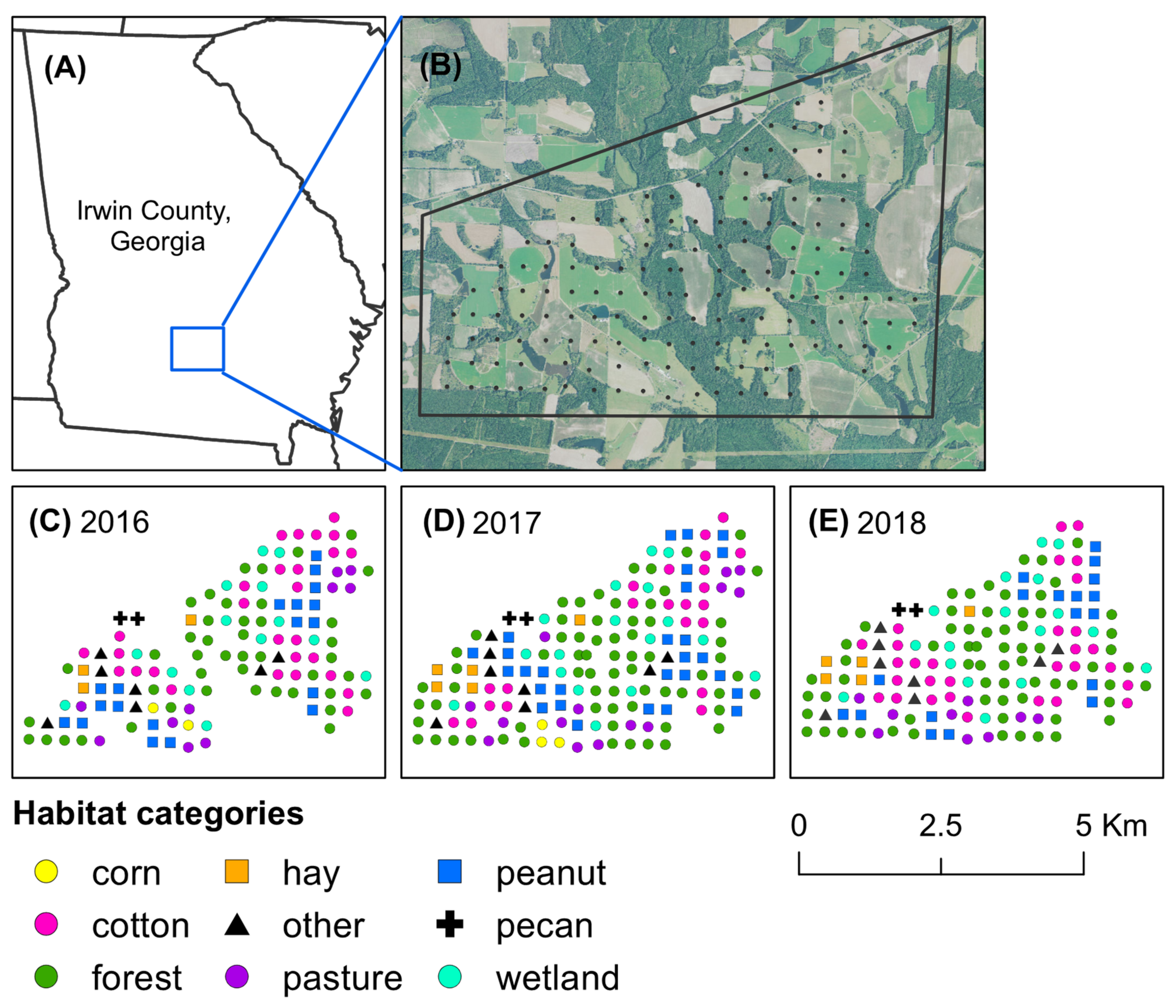




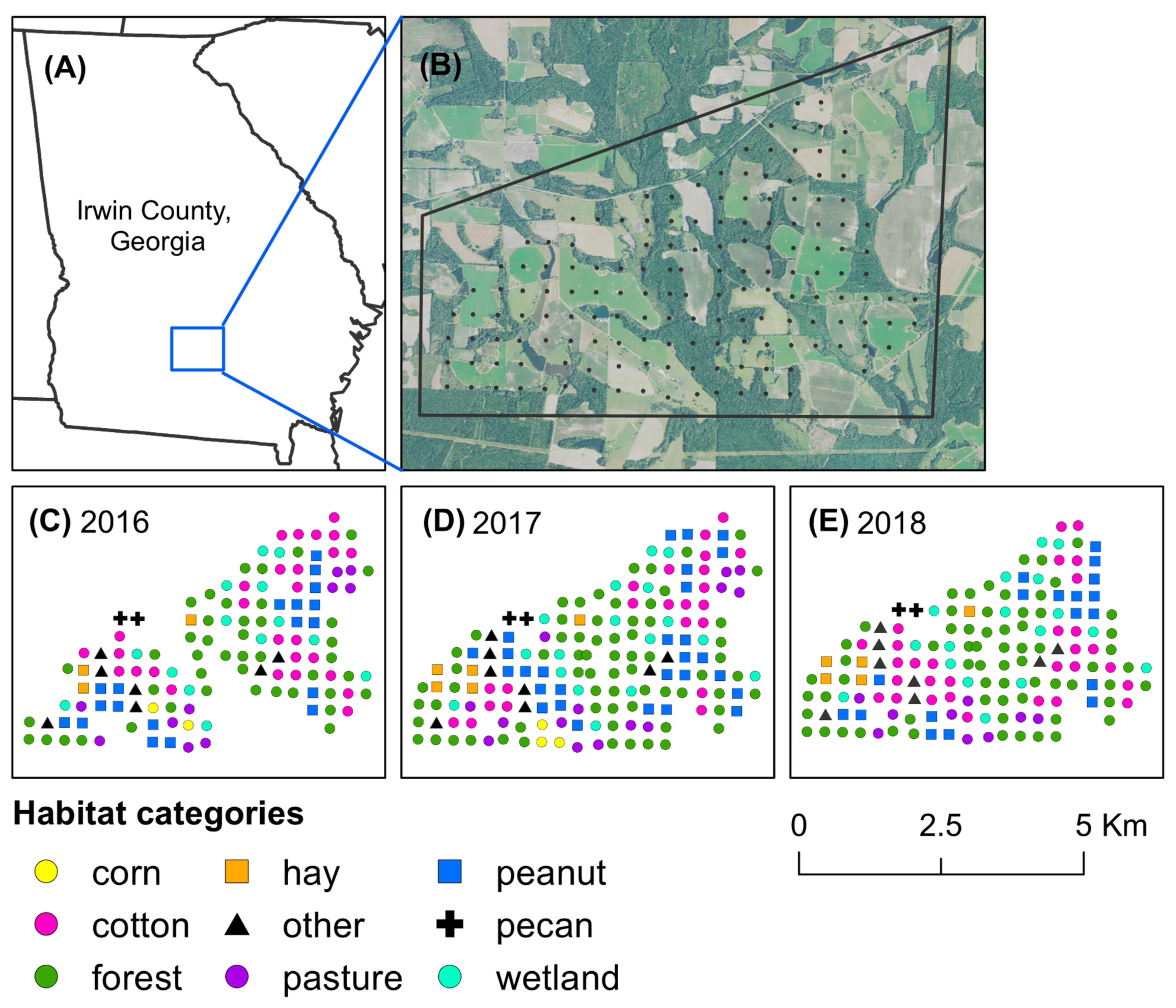{kind=link}
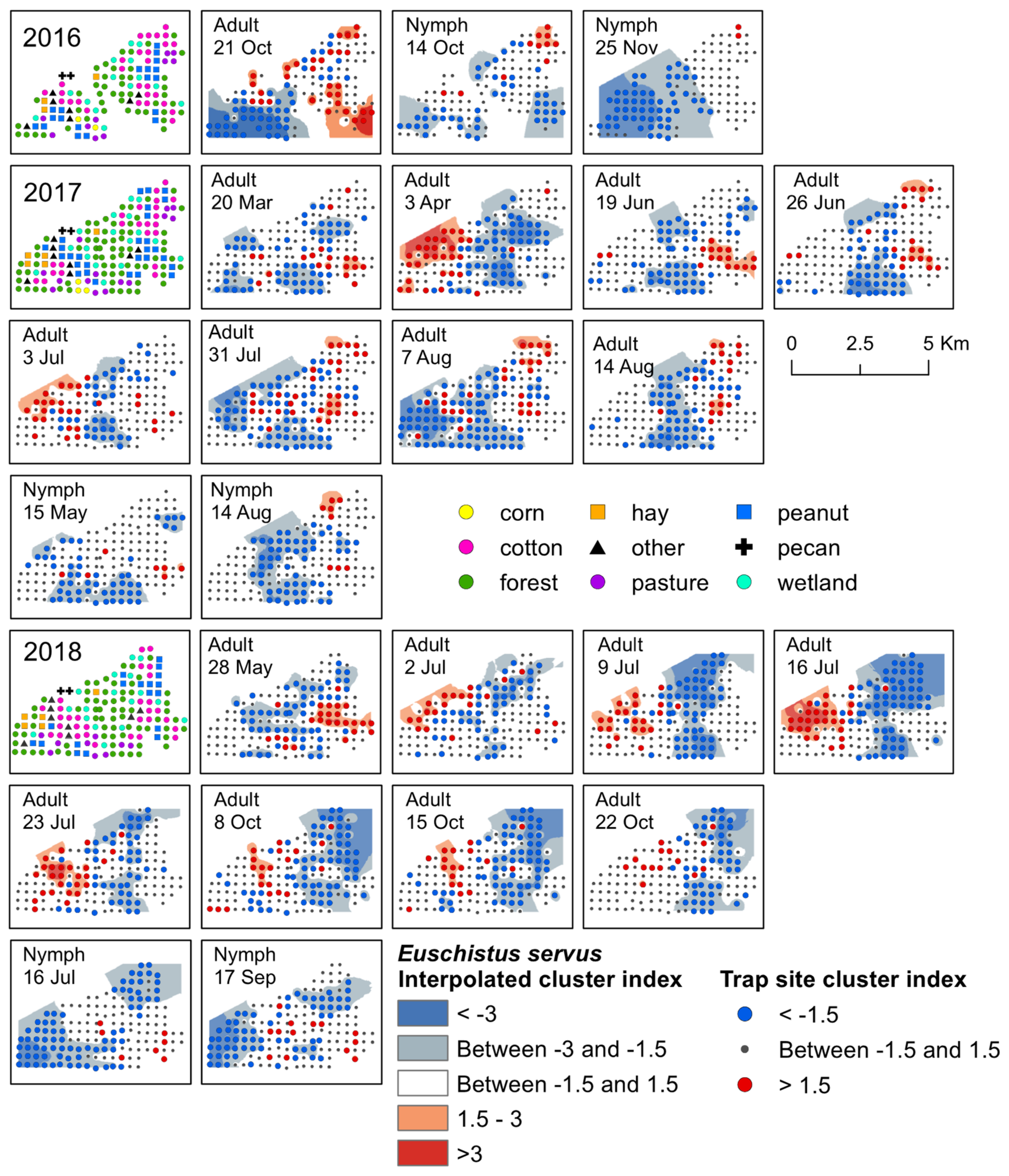{kind=link}
{kind=link}
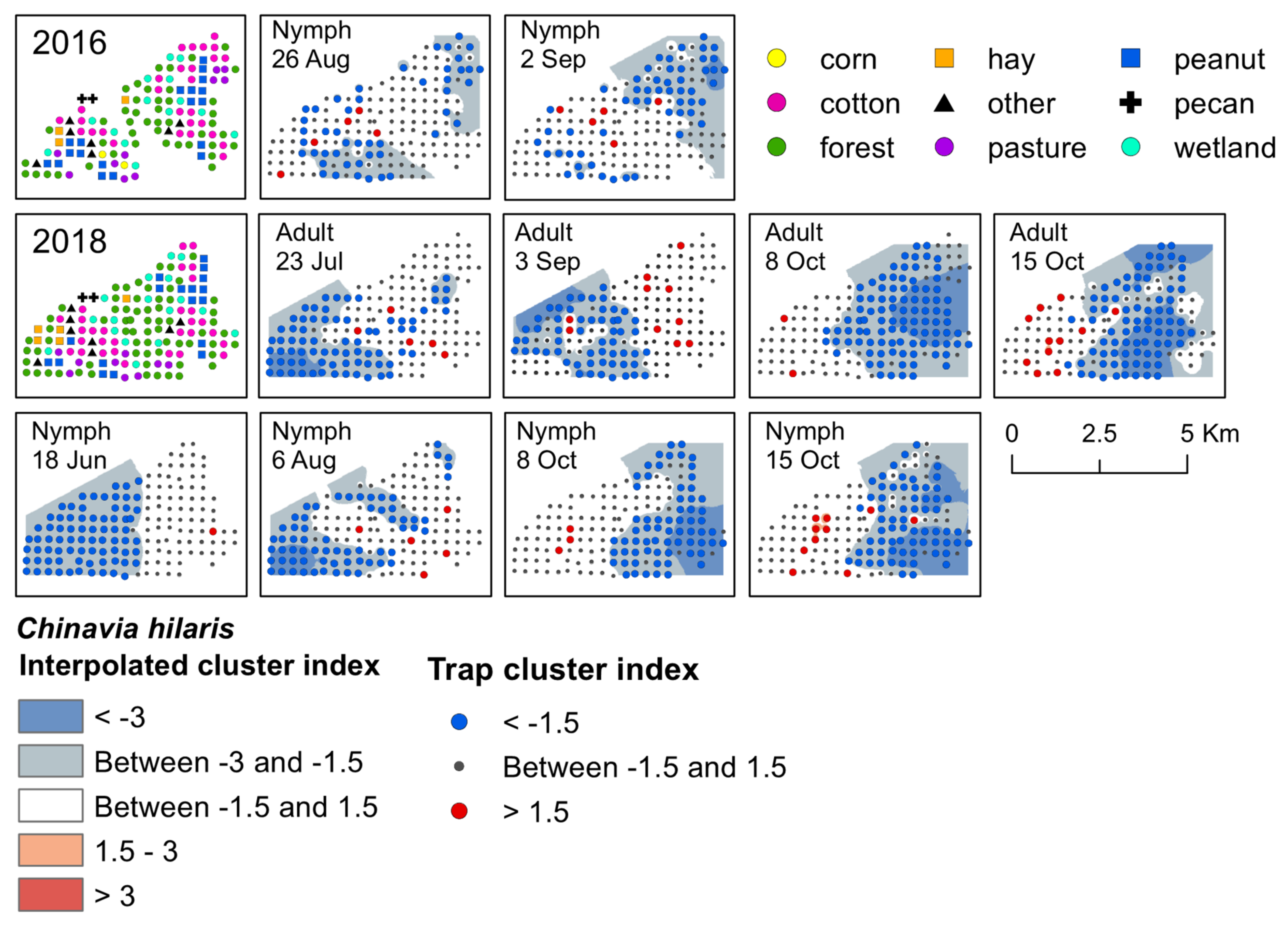{kind=link}
| Row | Forest + Wetland | Other | |
|---|---|---|---|
| Species | Mean ± SE | Mean ± SE | Mean ± SE |
| E. servus adult | 7.61 ± 0.24 a | 2.74 ± 0.07 c | 3.27 ± 0.14 b |
| E. servus nymph | 0.15 ± 0.001 a | 0.16 ± 0.01 a | 0.12 ± 0.01 a |
| E. tristigmus adult | 0.62 ± 0.03 b | 2.22 ± 0.05 a | 0.35 ± 0.04 c |
| E. tristigmus nymph | 0.09 ± 0.01 b | 0.39 ± 0.02 a | 0.07 ± 0.01 c |
| C. hilaris adult | 0.07 ± 0.01 a | 0.04 ± 0.004 a | 0.01 ± 0.002 a |
| C. hilaris nymph | 0.09 ± 0.01 ab | 0.16 ± 0.02 a | 0.01 ± 0.004 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grabarczyk, E.E.; Cottrell, T.E.; Tillman, G. Characterizing the Spatiotemporal Distribution of Three Native Stink Bugs (Hemiptera: Pentatomidae) across an Agricultural Landscape. Insects 2021, 12, 854. https://doi.org/10.3390/insects12100854
Grabarczyk EE, Cottrell TE, Tillman G. Characterizing the Spatiotemporal Distribution of Three Native Stink Bugs (Hemiptera: Pentatomidae) across an Agricultural Landscape. Insects. 2021; 12(10):854. https://doi.org/10.3390/insects12100854
Chicago/Turabian StyleGrabarczyk, Erin E., Ted E. Cottrell, and Glynn Tillman. 2021. "Characterizing the Spatiotemporal Distribution of Three Native Stink Bugs (Hemiptera: Pentatomidae) across an Agricultural Landscape" Insects 12, no. 10: 854. https://doi.org/10.3390/insects12100854
APA StyleGrabarczyk, E. E., Cottrell, T. E., & Tillman, G. (2021). Characterizing the Spatiotemporal Distribution of Three Native Stink Bugs (Hemiptera: Pentatomidae) across an Agricultural Landscape. Insects, 12(10), 854. https://doi.org/10.3390/insects12100854