Host Suitability for Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) Differed Significantly among Crapemyrtle Species




Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Test Plants
2.2. CMBS and Host Range Test
2.3. Statistical Analysis
3. Results
3.1. Host Range Confirmation
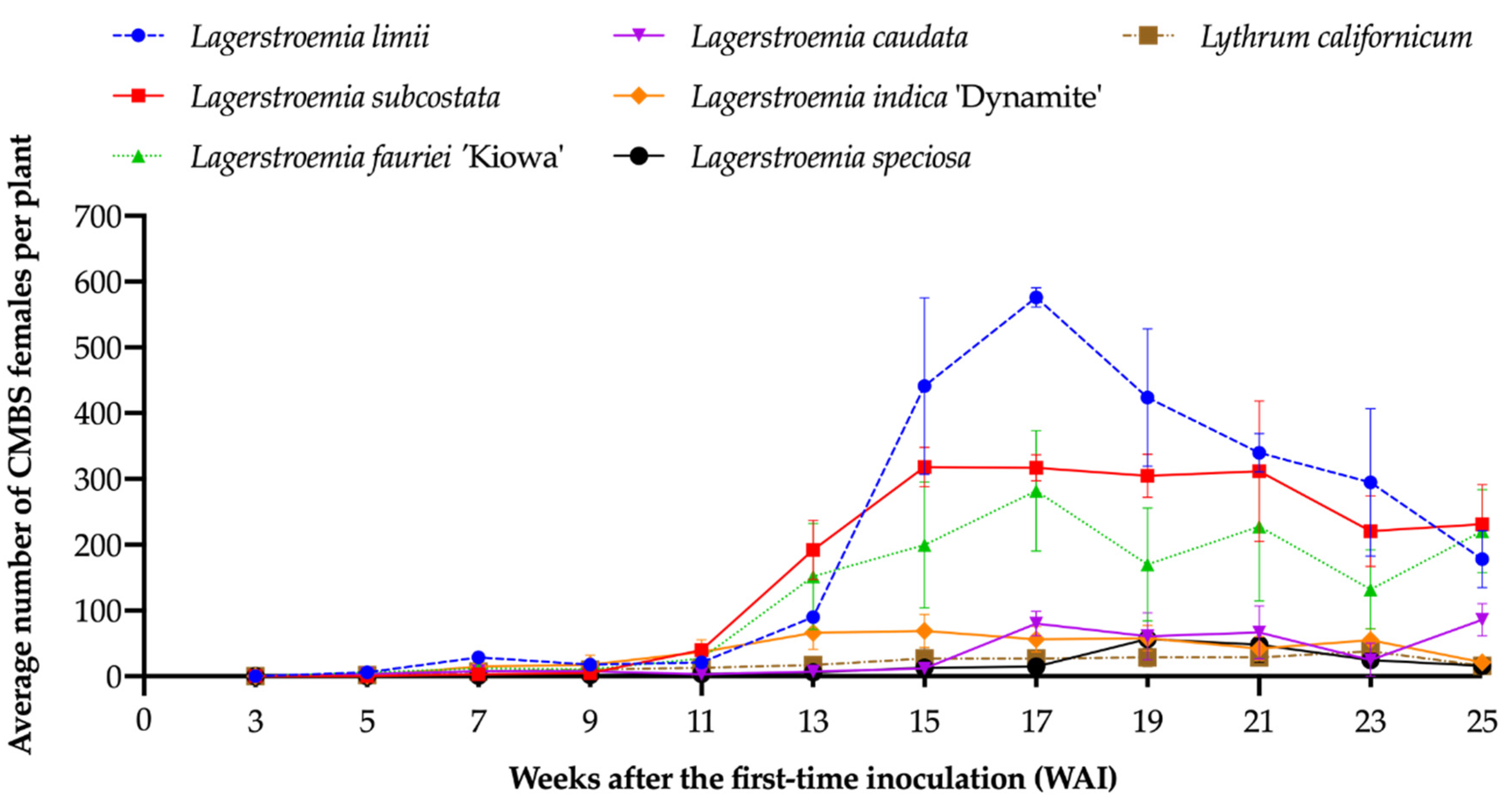
3.2. The Suitability for CMBS Differed Significantly among the Lagerstroemia Species
3.3. The Effect of the Species–Time Interaction on the Weeks after Inoculation (WAI) when the Number of CMBS Increased Significantly Compared to the Previous Week on Different Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Egolf, D. ‘Muskogee’ and ’Natchez’ Lagerstroemia [New cultivars, crapemyrtle]. HortScience 1981, 16, 576–577. [Google Scholar]
- Egolf, D.R. Choctaw’ Lugerstroemia. HortScience 1990, 25, 992–993. [Google Scholar] [CrossRef]
- Egolf, D.R. Caddo’ and Tonto’ Lugerstroemia. HortScience 1990, 25, 585–587. [Google Scholar] [CrossRef]
- Pooler, M.R. Arapaho’andCheyenne’Lagerstroemia. HortScience 2006, 41, 855–856. [Google Scholar] [CrossRef]
- Herbert, J.J.; Mizell, R., III; McAuslane, H. Host preference of the crapemyrtle aphid (Hemiptera: Aphididae) and host suitability of crapemyrtle cultivars. Environ. Entomol. 2009, 38, 1155–1160. [Google Scholar] [CrossRef] [PubMed]
- Mizell, R.F., III; Knox, G.W. Susceptibility of crape myrtle, Lagerstroemia indica L., to the crapemyrtle aphid (Homoptera: Aphididae) in north Florida. J. Entomol. Sci. 1993, 28, 1–7. [Google Scholar] [CrossRef]
- Cabrera, R.I.; Reinert, J.A.; McKenney, C.B. Differential resistance among crape myrtle (Lagerstroemia) species, hybrids, and cultivars to foliar feeding by adult flea beetles (Altica litigata). HortScience 2008, 43, 403–407. [Google Scholar] [CrossRef]
- Pettis, G.V.; Boyd, D.W., Jr.; Braman, S.K.; Pounders, C. Potential resistance of crape myrtle cultivars to flea beetle (Coleoptera: Chrysomelidae) and Japanese beetle (Coleoptera: Scarabaeidae) damage. J. Econ. Entomol. 2004, 97, 981–992. [Google Scholar] [CrossRef]
- Kanda, S. Studies on Coccidae from Corea. Insect World 1941, 45, 356–361. [Google Scholar]
- Kwon, T.; Park, S.; Choi, B.; Kwon, Y. Seasonal occurrence and chemical control effects of Eriococcus largerstroemiae Kuwana on persimmon trees. Korean J. Appl. Entomol. 1995, 34, 295–299. [Google Scholar]
- Suh, S.-J. Notes on some parasitoids (Hymenoptera: Chalcidoidea) associated with Acanthococcus lagerstroemiae (Kuwana) (Hemiptera: Eriococcidae) in the Republic of Korea. Insecta Mundi. 2019, 0690, 1–5. [Google Scholar]
- Suh, S.-J.; Yu, H.M.; Hong, K.-J. List of intercepted scale insects at Korean ports of entry and potential invasive species of scale insects to Korea (Hemiptera: Coccoidea). Korean J. Appl. Entomol. 2013, 52, 141–160. [Google Scholar] [CrossRef]
- Park, J.; Kim, Y.; Kim, S.; Park, I.; Kim, K. Seasonal occurrence, host preference and hatching behavior of Eriococcus lagerstroemiae. Korean J. Appl. Entomol 1993, 32, 83–89. [Google Scholar]
- Waku, Y.; Manabe, Y. Fine structure of the wax gland in a scale insect, Eriococcus lagerstraemiae KUWANA (Homoptera: Ericoccidae). Appl. Entomol. Zool. 1981, 16, 94–102. [Google Scholar] [CrossRef]
- Green, E. Observations on British Coccidae in 1914, with descriptions of new species. Entomol. Mon. Mag. 1915, 51, 175–185. [Google Scholar] [CrossRef]
- Zhang, Z.; Shi, Y. Studies on the morphology and biology of Eriococcus lagerstroemiae kuwana. J. Shandong Agric. Univ. 1986, 2, 61–66. [Google Scholar]
- Wang, Z.; Chen, Y.; Gu, M.; Vafaie, E.; Merchant, M.; Diaz, R. Crapemyrtle bark scale: A new threat for crapemyrtles, a popular landscape plant in the US. Insects 2016, 7, 78. [Google Scholar] [CrossRef]
- Work, T.T.; McCullough, D.G.; Cavey, J.F.; Komsa, R. Arrival rate of nonindigenous insect species into the United States through foreign trade. Biol. Invasions 2005, 7, 323. [Google Scholar] [CrossRef]
- Hendrix, P.F.; Bohlen, P.J. Exotic earthworm invasions in North America: Ecological and policy implications: Expanding global commerce may be increasing the likelihood of exotic earthworm invasions, which could have negative implications for soil processes, other animal and plant species, and importation of certain pathogens. Bioscience 2002, 52, 801–811. [Google Scholar]
- EDDMapS. Early Detection & Distribution Mapping System. The University of Georgia—Center for Invasive Species and Ecosystem Health. 2020. Available online: http://www.eddmaps.org/ (accessed on 6 January 2020).
- Louda, S.M.; Kendall, D.; Connor, J.; Simberloff, D. Ecological effects of an insect introduced for the biological control of weeds. Science 1997, 277, 1088–1090. [Google Scholar] [CrossRef]
- Worner, S.; Gevrey, M. Modelling global insect pest species assemblages to determine risk of invasion. J. Appl. Ecol. 2006, 43, 858–867. [Google Scholar] [CrossRef]
- Venette, R.C.; Kriticos, D.J.; Magarey, R.D.; Koch, F.H.; Baker, R.H.; Worner, S.P.; Gómez Raboteaux, N.N.; McKenney, D.W.; Dobesberger, E.J.; Yemshanov, D. Pest risk maps for invasive alien species: A roadmap for improvement. BioScience 2010, 60, 349–362. [Google Scholar] [CrossRef]
- Miller, D.R.; Miller, G.L. Systematic analysis of Acanthococcus (Homoptera: Coccoidea: Eriococcidae) in the western United States. Trans. Am. Entomol. Soc. 1992, 118, 1–106. [Google Scholar]
- Miller, D.R.; Miller, G.L. Eriococcidae of the eastern United States. Contrib. Am. Entomol. Inst. 1993, 27, 91. [Google Scholar]
- Merchant, M.E.; Gu, M.; Robbins, J.; Vafaie, E.; Barr, N.; Tripodi, A.D.; Szalanski, A.L. Discovery and Spread of Eriococcus lagerstroemiae Kuwana (Hemiptera: Eriococcidae), a New Invasive Pest of Crape Myrtle, Lagerstroemia spp. 2014. Available online: https://www.researchgate.net/profile/Allen_Szalanski/publication/267521252_Discovery_and_spread_of_Eriococcus_lagerstroemiae_Hemiptera_Eriococcidae_a_new_invasive_pest_of_crape_myrtle/links/56c1e9bf08ae44da37ff46af.pdf (accessed on 20 May 2019).
- Jiang, N.; Xu, H. Observertion on Eriococcus lagerostroemiae Kuwana. J. Anhui Agric. Coll. 1998, 25, 142–144. [Google Scholar]
- Miller, D.R. Selected scale insect groups (Hemiptera: Coccoidea) in the southern region of the United States. Fla. Entomol. 2005, 88, 482–501. [Google Scholar]
- Ahmed, M.Z.; Ray, C.H.; Moore, M.R.; Miller, D.R. The Matsucoccus Cockerell, 1909 of Florida (Hemiptera: Coccomorpha: Matsucoccidae): Potential pests of Florida pines. Insecta Mundi. 2020, 0810, 1–31. [Google Scholar]
- Wang, Z.; Chen, Y.; Diaz, R. Temperature-dependent development and host range of crapemyrtle bark scale, Acanthococcus lagerstroemiae (Kuwana)(Hemiptera: Eriococcidae). Fla. Entomol. 2019, 102, 181–186. [Google Scholar]
- Gu, M.; Merchant, M.; Robbins, J.; Hopkins, J. Crape Myrtle Bark Scale: A New Exotic Pest. Texas A&M AgriLife Ext. 2014. Available online: https://agrilifecdn.tamu.edu/citybugstest/files/2010/05/EHT-049-Crape-myrtle-bark-scale.pdf (accessed on 15 May 2018).
- Planthardiness. USDA Plant Hardiness Zone Map. Agricultural Research Service, U.S. Department of Agriculture. 2012. Available online: https://planthardiness.ars.usda.gov/ (accessed on 4 February 2019).
- Wang, Z.; Chen, Y.; Diaz, R. Thermal tolerance and prediction of northern distribution of the Crapemyrtle Bark Scale (Hemiptera: Eriococcidae). Environ. Entomol. 2019, 48, 641–648. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, Y.; Diaz, R.; Laine, R.A. Physiology of crapemyrtle bark scale, Acanthococcus lagerstroemiae (Kuwana), associated with seasonally altered cold tolerance. J. Insect Physiol. 2019, 112, 1–8. [Google Scholar] [CrossRef]
- Insausti, P.; Ploschuk, E.L.; Izaguirre, M.M.; Podworny, M. The effect of sunlight interception by sooty mold on chlorophyll content and photosynthesis in orange leaves (Citrus sinensis L.). Eur. J. Plant. Pathol 2015, 143, 559–565. [Google Scholar] [CrossRef]
- Vafaie, E.K. Bark and Systemic Insecticidal Control of Acanthococcus (=Eriococcus) lagerstroemiae (Hemiptera: Eriococcidae) on Potted Crapemyrtles, 2017. Arthropod Manag. Tests 2019, 44, tsy109. [Google Scholar] [CrossRef]
- Merchant, M.; Garcia, J.; Gu, M. Longevity of imidacloprid as a soil treatment for crapemyrtle bark scale on crapemyrtle, 2016–2017. Arthropod Manag. Tests 2018, 43, tsy012. [Google Scholar] [CrossRef]
- Matos Franco, G. Impacts of Commercial Biopesticides on Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) and Beneficial Insects. Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2020. [Google Scholar]
- Bolques, A.; Knox, G. Growth and flowering phenology of six crape myrtle cultivars in north Florida. In Proceedings of the Southern Nursery Association Research Conference, Atlanta, GA, USA, 6–8 August 1997; Volume 42, pp. 365–368. [Google Scholar]
- Pounders, C.T.; Blythe, E.K.; Fare, D.C.; Knox, G.W.; Sibley, J.L. Crapemyrtle genotype× environment interactions, and trait stability for plant height, leaf-out, and flowering. HortScience 2010, 45, 198–207. [Google Scholar] [CrossRef]
- Braman, S.K.; Quick, J.C. Differential bee attraction among crape myrtle cultivars (Lagerstroemia spp.: Myrtales: Lythraceae). Environ. Entomol. 2018, 47, 1203–1208. [Google Scholar] [CrossRef]
- Riddle, T.C.; Mizell III, R.F. Use of crape myrtle, Lagerstroemia (Myrtales: Lythraceae), cultivars as a pollen source by native and non-native bees (Hymenoptera: Apidae) in Quincy, Florida. Fla. Entomol. 2016, 99, 38–46. [Google Scholar] [CrossRef]
- Kremen, C.; Williams, N.M.; Bugg, R.L.; Fay, J.P.; Thorp, R.W. The area requirements of an ecosystem service: Crop pollination by native bee communities in California. Ecol. Lett. 2004, 7, 1109–1119. [Google Scholar] [CrossRef]
- Lau, P.; Bryant, V.; Ellis, J.D.; Huang, Z.Y.; Sullivan, J.; Schmehl, D.R.; Cabrera, A.R.; Rangel, J. Seasonal variation of pollen collected by honey bees (Apis mellifera) in developed areas across four regions in the United States. PLoS ONE 2019, 14, e0217294. [Google Scholar] [CrossRef]
- Van der Sluijs, J.P.; Simon-Delso, N.; Goulson, D.; Maxim, L.; Bonmatin, J.-M.; Belzunces, L.P. Neonicotinoids, bee disorders and the sustainability of pollinator services. Curr. Opin. Environ. Sustain. 2013, 5, 293–305. [Google Scholar] [CrossRef]
- Gill, R.J.; Ramos-Rodriguez, O.; Raine, N.E. Combined pesticide exposure severely affects individual-and colony-level traits in bees. Nature 2012, 491, 105–108. [Google Scholar] [CrossRef]
- Vanbergen, A.J.; Initiative, T.I.P. Threats to an ecosystem service: Pressures on pollinators. Front. Ecol. Environ. 2013, 11, 251–259. [Google Scholar] [CrossRef]
- Bernays, E.A.; Chapman, R.F. Host-Plant Selection by Phytophagous Insects; Chapman & Hall: New York, NY, USA, 1994; Volume 2, pp. 95–165. [Google Scholar]
- Schaffner, U. Host Range Testing of Insects for Biological Weed Control: How Can It Be Better Interpreted? Data on the host range of biocontrol candidates are particularly relevant in assessing potential detrimental effects to nontarget organisms. BioScience 2001, 51, 951–959. [Google Scholar]
- Ma, J.-H. Occurrence and biological characteristics of Eriococcus lagerostroemiae Kuwana in Panxi district. South China Fruits 2011, 5, 12–14. [Google Scholar]
- Li, S. The Occurrence and Control Techniques of Scale Insect in Weifang Region’s Landscape Plant. Weifang High. Vocat. Educ. 2009, 5, 68–72. [Google Scholar]
- Schultz, P.B.; Szalanski, A.L. Hypericum kalmianum (St. Johnswort) confirmed as a new host of the crapemyrtle bark scale in Virginia, USA. J. Agric. Urban. Entomol. 2019, 35, 12–14. [Google Scholar] [CrossRef]
- Kozár, F.; Kaydan, M.B.; Konczné Benedicty, Z.; Szita, É. Acanthococcidae and Related Families of the Palaearctic Region; Plant Protection Institute, Centre for Agricultural Research, Hungarian: Budapest, Hungary, 2013. [Google Scholar]
- Xie, R.; Wu, B.; Dou, H.; Liu, C.; Knox, G.W.; Qin, H.; Gu, M. Feeding Preference of Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) on Different Species. Insects 2020, 11, 399. [Google Scholar] [CrossRef]
- Gu, M. Alternative Hosts of Crapemyrtle Bark Scale. Texas A&M AgriLife Ext. 2018. Available online: https://cdn-ext.agnet.tamu.edu/wp-content/uploads/2018/10/EHT-103-alternative-hosts-of-crapemyrtle-bark-scale.pdf (accessed on 15 May 2019).
- efloras.org. Lagerstroemia Caudata. Available online: http://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id=200014638 (accessed on 15 March 2019).
- Brickell, C.D. International Code of Nomenclature for Cultivated Plants-1980; Bohn, Scheltema & Holkema: Utrecht, The Netherlands, 1980. [Google Scholar]
- Pooler, M.R.; Dix, R.L. Chickasaw’,Kiowa’, and Pocomoke’Lagerstroemia. HortScience 1999, 34, 361–363. [Google Scholar] [CrossRef]
- PatFT. Crape Myrtle Shrub Named ‘White II’. USPTO Patent Full-Text and Image Database. Available online: http://patft.uspto.gov/netacgi/nph-Parser?Sect1=PTO2&Sect2=HITOFF&u=%2Fnetahtml%2FPTO%2Fsearch-adv.htm&r=2&f=G&l=50&d=PTXT&p=1&S1=PP10296&OS=PP10296&RS=PP10296 (accessed on 10 July 2020).
- MONROVIA. Dynamite Crape Myrtle, Lagerstroemia Indica ‘White II’. Available online: https://www.monrovia.com/plant-catalog/plants/251/dynamite-crape-myrtle/ (accessed on 15 July 2019).
- Bayton, R.; Grimshaw, J. 2019. Available online: https://treesandshrubsonline.org/articles/lagerstroemia/lagerstroemia-limii/ (accessed on 15 July 2019).
- CABI. Lagerstroemia speciosa (Pride of India). Available online: https://www.cabi.org/ISC/datasheet/29704 (accessed on 7 October 2020).
- Klein, G.; Kim, J.; Himmeldirk, K.; Cao, Y.; Chen, X. Antidiabetes and anti-obesity activity of Lagerstroemia speciosa. Evid. Based Complement. Altern. Med. 2007, 4, 547546. [Google Scholar] [CrossRef]
- efloras.org. Lagerstroemia subcostata. Available online: http://www.efloras.org/florataxon.aspx?flora_id=2&taxon_id=200014650 (accessed on 15 March 2019).
- Calflora. Lythrum californicum. Available online: https://www.calflora.org//cgi-bin/species_query.cgi?where-taxon=Lythrum+californicum (accessed on 7 October 2020).
- Craig, T.P.; Price, P.W.; Itami, J.K. Facultative sex ratio shifts by a herbivorous insect in response to variation in host plant quality. Oecologia 1992, 92, 153–161. [Google Scholar] [CrossRef]
- Mopper, S.; Whitham, T.G. The plant stress paradox: Effects on pinyon sawfly sex ratios and fecundity. Ecology 1992, 73, 515–525. [Google Scholar] [CrossRef]
- Fujita, A.; Mitsuhashi, J. Effects of dietary amino acids on the production of sexual morph by the green peach aphid, Myzus persicae. Arch. Insect Biochem. Physiol. 1995, 29, 259–268. [Google Scholar] [CrossRef]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Charnov, E.L. The Theory of Sex. Allocation. (MPB-18), Volume 18; Princeton University Press: Princetion, NJ, USA, 2020; Volume 113. [Google Scholar]
- Lewis, S.; South, A.; Burns, R.; Al-Wathiqui, N. Nuptial gifts. Curr. Biol. 2011, 21, R644–R645. [Google Scholar] [CrossRef] [PubMed]
- Boggs, C.L. Male nuptial gifts: Phenotypic consequences and evolutionary implications. In Insect Reproduction; CRC Press: Boca Raton, FL, USA, 2018; pp. 215–242. [Google Scholar]
- Schoonhoven, L.M.; Van Loon, B.; van Loon, J.J.; Dicke, M. Insect-Plant Biology; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Smith, B. Effect of the plant alkaloid sparteine on the distribution of the aphid Acyrthosiphon spartii (Koch.). Nature 1966, 212, 213–214. [Google Scholar] [CrossRef]
- Montgomery, M.; Arn, H. Feeding response of Aphis pomi, Myzus persicae, and Amphorophora agathonica to phlorizin. J. Insect Physiol. 1974, 20, 413–421. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, B.W.; Schroeder, F.C.; Jander, G. Identification of indole glucosinolate breakdown products with antifeedant effects on Myzus persicae (green peach aphid). Plant J. 2008, 54, 1015–1026. [Google Scholar] [CrossRef]
- Basu, S.; Kundu, P.; Sinhababu, A. Characterization of fatty acid and sterol composition of seed lipid of Lagerstroemia speciosa Pers. Res. Chem. Intermed. 2015, 41, 6511–6522. [Google Scholar] [CrossRef]
- Basu, S. Chemical Investigation on the Plant Species Lagerstroemia speciosa Pers. Ph.D. Thesis, The University of Burdwan, Bardhaman, West Bengal, India, 2016. [Google Scholar]
- Jehan, C.M.; Daulatabad, D.; Mirajkar, A.M. A keto fatty acid from Lagerstroemia speciosa seed oil. Phytochemistry 1990, 29, 2323–2324. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Recommended U.S. Department of Agriculture Plant Hardiness Zone | Native Origin | Mature Height (m) * |
|---|---|---|---|
| Lagerstroemia caudata | 9–10 | China (Guangdong, Guangxi, Jiangxi) | 18.0–30.0 [56] |
| L. fauriei ‘Kiowa’ | 6–9 | Japan | 3.0–4.6 [57,58] |
| L. indica ‘Dynamite’ | 6–10 | The USA (Oklahoma) | 4.6–6.0 [59,60] |
| L. limii | 8–10 | China (Fujian, Hubei, and Zhejiang) | 4.0–7.0 [61] |
| L. speciosa | 9–13 | China (Fujian, Guangdong, Guangxi, Hainan, Yunnan), India, Malaysia, Philippines, Sri Lanka, Vietnam | 20.0–40.0 [62,63] |
| L. subcostata | 4–11 | Japan (Ryukyu Islands), China (Anhui, Guangdong, Guangxi, Hubei, Hunan, Jiangxi, Jiangsu, Qinghai Sichuan, Taiwan, Zhejiang) | 6.1 to 9.1 [64] |
| Lythrum californicum | 4–9 | Northern Mexico, central USA (Arizona, California, Kansas, New Mexico, Nevada, Oklahoma, Texas) | Up to 1.5 [65] |
| Plant Species | No. Males (95% CI z) | No. Females (95% CI) | Sex Ratio |
|---|---|---|---|
| L. subcostata | 139 (47–405) a y | 49 (18–131) ab | 2.8:1 |
| L. limii | 119 (40–348) ab | 63 (24–167) a | 1.9:1 |
| L. fauriei ‘Kiowa’ | 48 (16–140) ab | 30 (11–80) ab | 1.6:1 |
| L. indica ‘Dynamite’ | 40 (13–118) ab | 16 (6–43) ab | 2.5:1 |
| L. caudata | 19 (6–56) ab | 7 (2–20) ab | 2.7:1 |
| L. speciosa | 11 (3–33) b | 7 (2–19) b | 1.6:1 |
| Plant Species | The WAI When CMBS Significantly Increased | |
|---|---|---|
| CMBS Males | CMBS Females | |
| Lagerstroemia subcostata | 11 | 11 |
| Lagerstroemia limii | 11 | 11 |
| Lagerstroemia fauriei ‘Kiowa’ | 13 | 11 |
| Lagerstroemia indica ‘Dynamite’ | 11 | 11 |
| Lagerstroemia caudata | NA | 17 |
| Lagerstroemia speciosa | NA | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, B.; Xie, R.; Knox, G.W.; Qin, H.; Gu, M. Host Suitability for Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) Differed Significantly among Crapemyrtle Species. Insects 2021, 12, 6. https://doi.org/10.3390/insects12010006
Wu B, Xie R, Knox GW, Qin H, Gu M. Host Suitability for Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) Differed Significantly among Crapemyrtle Species. Insects. 2021; 12(1):6. https://doi.org/10.3390/insects12010006
Chicago/Turabian StyleWu, Bin, Runshi Xie, Gary W. Knox, Hongmin Qin, and Mengmeng Gu. 2021. "Host Suitability for Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) Differed Significantly among Crapemyrtle Species" Insects 12, no. 1: 6. https://doi.org/10.3390/insects12010006
APA StyleWu, B., Xie, R., Knox, G. W., Qin, H., & Gu, M. (2021). Host Suitability for Crapemyrtle Bark Scale (Acanthococcus lagerstroemiae) Differed Significantly among Crapemyrtle Species. Insects, 12(1), 6. https://doi.org/10.3390/insects12010006