Drosophila melanogaster Y Chromosome Genes Affect Male Sensitivity to Microbial Infections



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drosophila Rearing
2.2. Drosophila Oral Feeding Survival Assay
3. Results
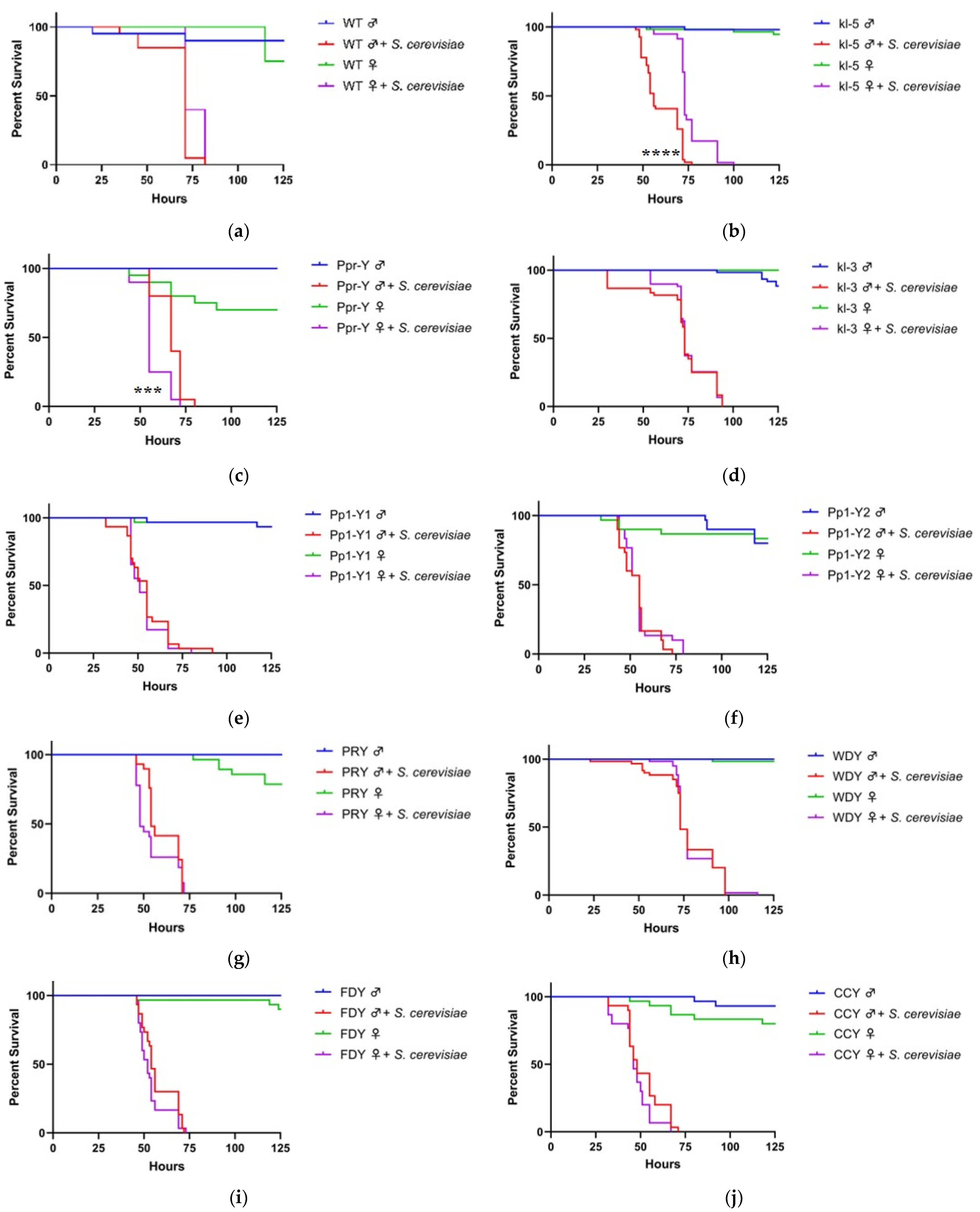
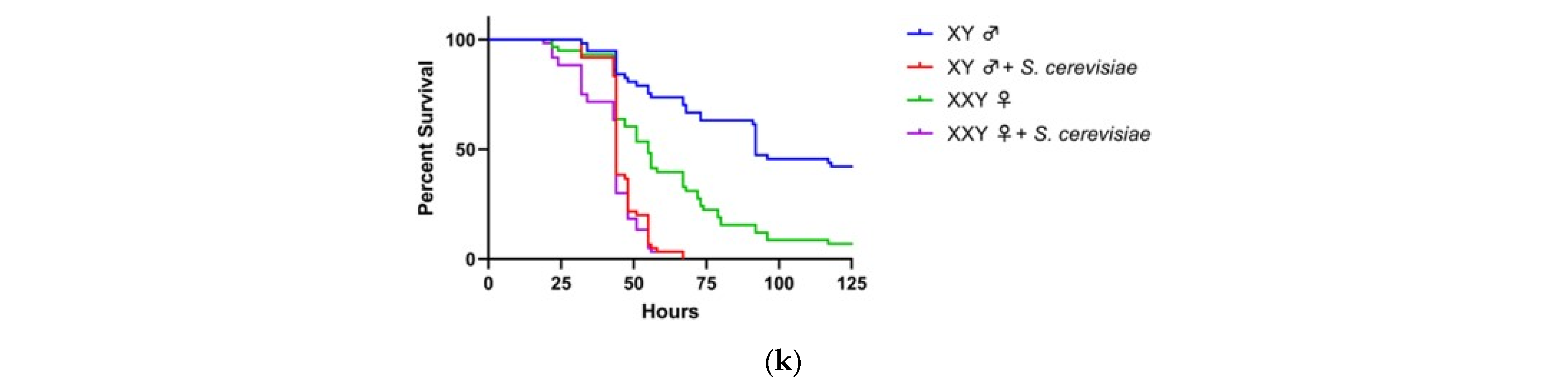
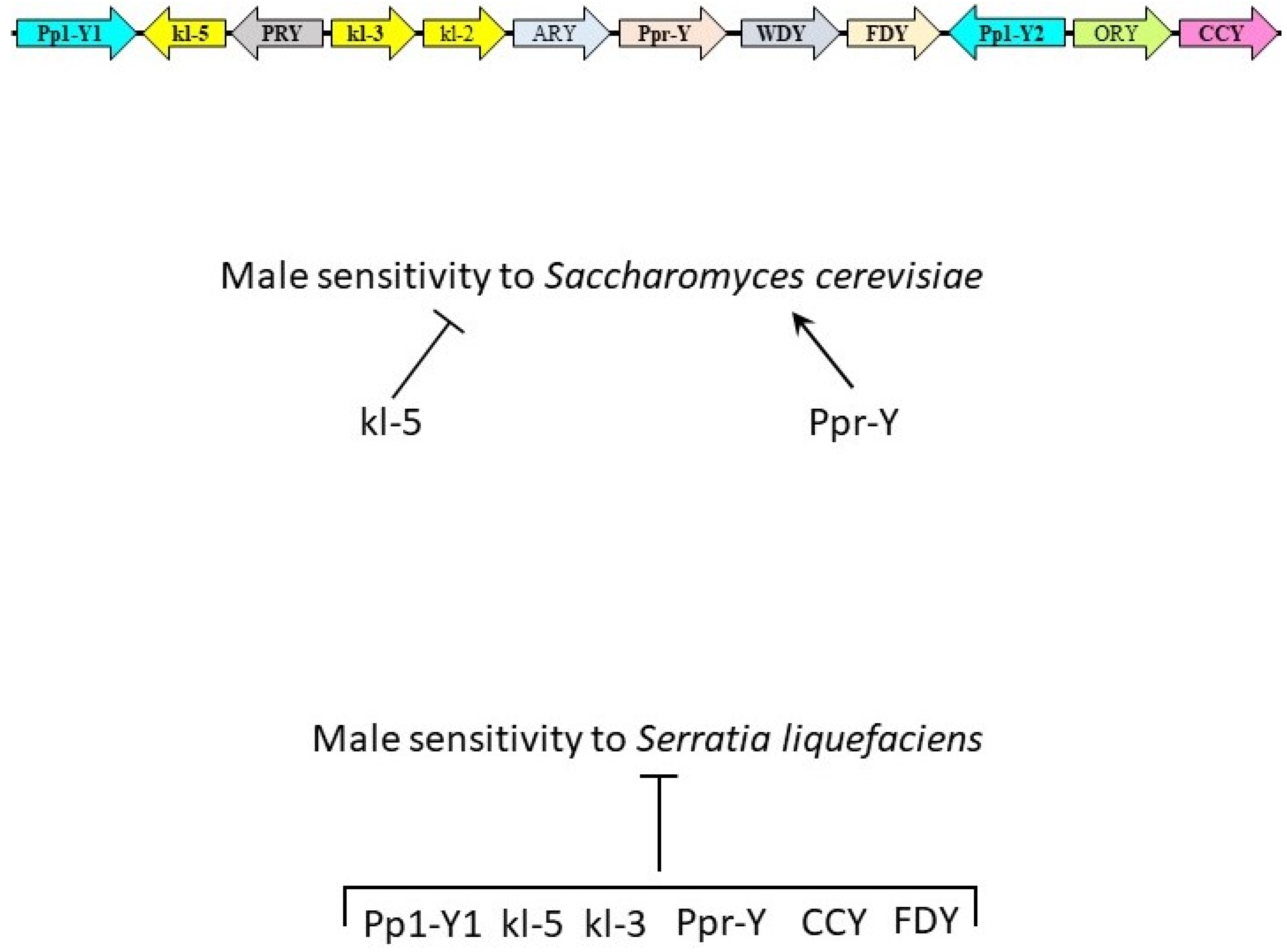
3.1. Y Genes Repress or Contribute to the Sensitivity of Fly Males to Yeast Infection
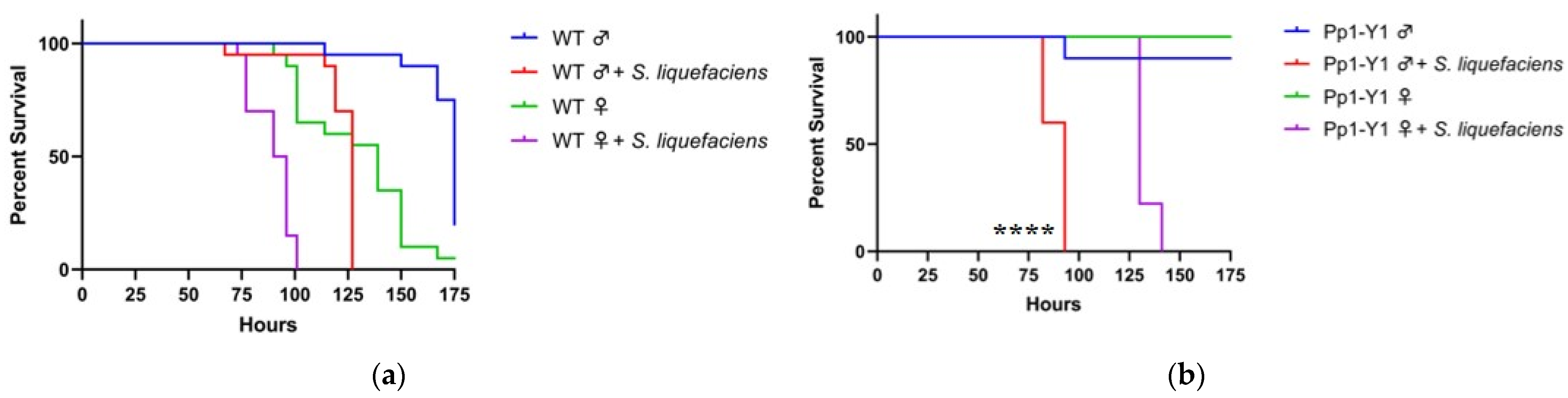
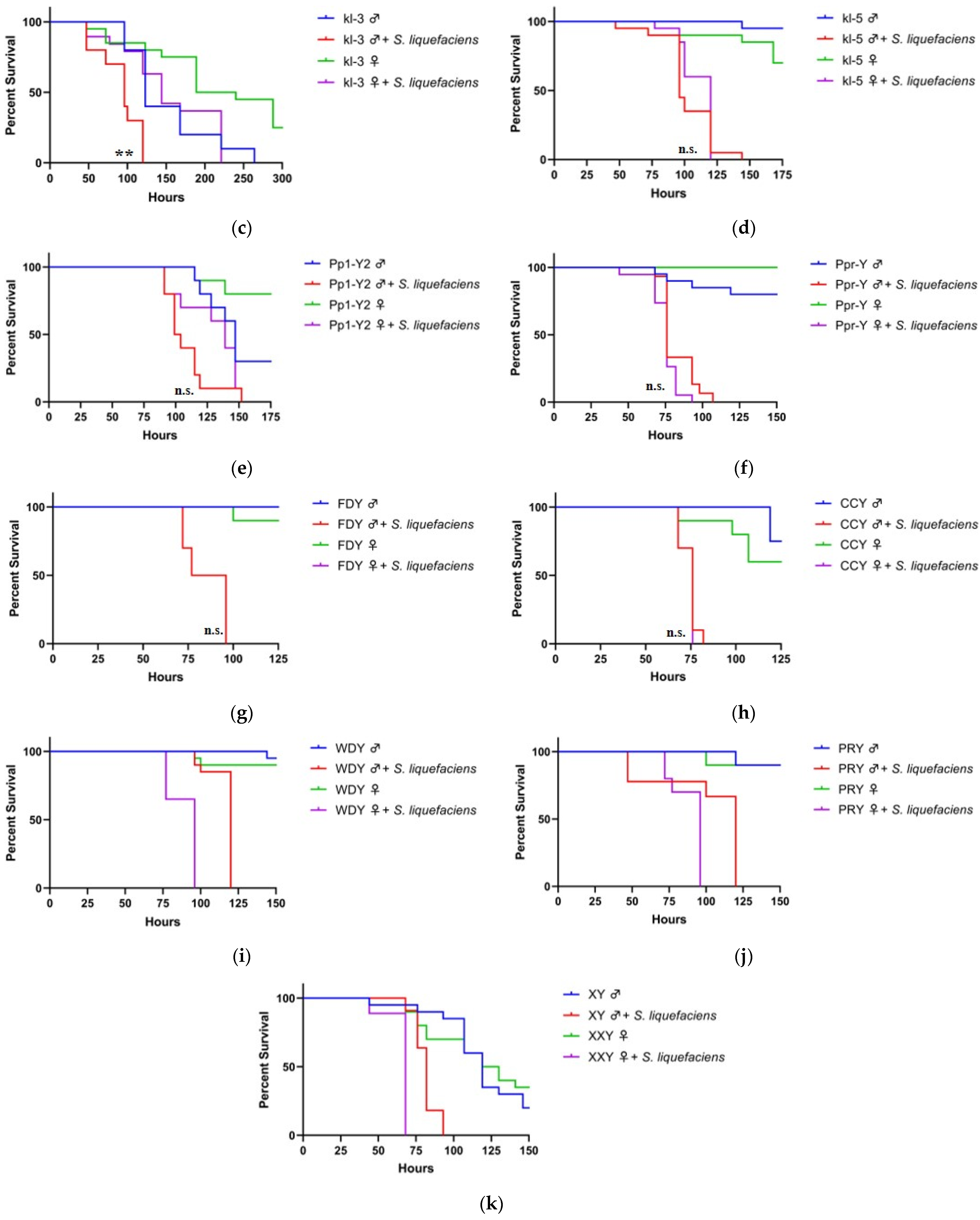
3.2. Y Genes Repress the Sensitivity of Fly Males to Bacterial Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Belmonte, R.L.; Corbally, M.K.; Duneau, D.F.; Regan, J.C. Sexual Dimorphisms in Innate Immunity and Responses to Infection in Drosophila melanogaster. Front. Immunol. 2019, 10, 3075. [Google Scholar] [CrossRef]
- Hanson, M.A.; Dostalova, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaitre, B. Correction: Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach. Elife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Tryselius, Y.; Samakovlis, C.; Kimbrell, D.A.; Hultmark, D. CecC, a cecropin gene expressed during metamorphosis in Drosophila pupae. Eur. J. Biochem. 1992, 204, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Buchon, N.; Silverman, N.; Cherry, S. Immunity in Drosophila melanogaster--from microbial recognition to whole-organism physiology. Nat. Rev. Immunol. 2014, 14, 796–810. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Z.; Lestradet, M.; Socha, C.; Schirmeier, S.; Schmitz, A.; Spenle, C.; Lefebvre, O.; Keime, C.; Yamba, W.M.; Bou Aoun, R.; et al. Enterocyte Purge and Rapid Recovery Is a Resilience Reaction of the Gut Epithelium to Pore-Forming Toxin Attack. Cell Host Microbe 2016, 20, 716–730. [Google Scholar] [CrossRef]
- Hill-Burns, E.M.; Clark, A.G. X-linked variation in immune response in Drosophila melanogaster. Genetics 2009, 183, 1477–1491. [Google Scholar] [CrossRef]
- Menon, D.U.; Meller, V.H. Identification of the Drosophila X chromosome: The long and short of it. RNA Biol. 2015, 12, 1088–1093. [Google Scholar] [CrossRef]
- Philip, P.; Stenberg, P. Male X-linked genes in Drosophila melanogaster are compensated independently of the Male-Specific Lethal complex. Epigenet. Chromatin 2013, 6, 35. [Google Scholar] [CrossRef]
- Koerich, L.B.; Wang, X.; Clark, A.G.; Carvalho, A.B. Low conservation of gene content in the Drosophila Y chromosome. Nature 2008, 456, 949–951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Luo, J.; Chen, J.; Dai, J.; Montell, C. The Role of Y Chromosome Genes in Male Fertility in Drosophila melanogaster. Genetics 2020, 215, 623–633. [Google Scholar] [CrossRef]
- Malone, J.H.; Oliver, B. The sex chromosome that refused to die. Bioessays 2008, 30, 409–411. [Google Scholar] [CrossRef]
- Salz, H.K.; Erickson, J.W. Sex determination in Drosophila: The view from the top. Fly 2010, 4, 60–70. [Google Scholar] [CrossRef]
- Krsticevic, F.J.; Schrago, C.G.; Carvalho, A.B. Long-Read Single Molecule Sequencing to Resolve Tandem Gene Copies: The Mst77Y Region on the Drosophila melanogaster Y Chromosome. G3 (Bethesda) 2015, 5, 1145–1150. [Google Scholar] [CrossRef]
- Hafezi, Y.; Sruba, S.R.; Tarrash, S.R.; Wolfner, M.F.; Clark, A.G. Dissecting Fertility Functions of Drosophila Y Chromosome Genes with CRISPR. Genetics 2020, 214, 977–990. [Google Scholar] [CrossRef] [PubMed]
- Vibranovski, M.D.; Koerich, L.B.; Carvalho, A.B. Two new Y-linked genes in Drosophila melanogaster. Genetics 2008, 179, 2325–2327. [Google Scholar] [CrossRef] [PubMed]
- Lemos, B.; Branco, A.T.; Hartl, D.L. Epigenetic effects of polymorphic Y chromosomes modulate chromatin components, immune response, and sexual conflict. Proc. Natl. Acad. Sci. USA 2010, 107, 15826–15831. [Google Scholar] [CrossRef] [PubMed]
- Kutch, I.C.; Fedorka, K.M. Y-linked variation for autosomal immune gene regulation has the potential to shape sexually dimorphic immunity. Proc. Biol. Sci. 2015, 282, 20151301. [Google Scholar] [CrossRef]
- Bi, E.; Pringle, J.R. ZDS1 and ZDS2, genes whose products may regulate Cdc42p in Saccharomyces cerevisiae. Mol. Cell Biol. 1996, 16, 5264–5275. [Google Scholar] [CrossRef]
- Nehme, N.T.; Liegeois, S.; Kele, B.; Giammarinaro, P.; Pradel, E.; Hoffmann, J.A.; Ewbank, J.J.; Ferrandon, D. A model of bacterial intestinal infections in Drosophila melanogaster. PLoS Pathog. 2007, 3, e173. [Google Scholar] [CrossRef]
- Groves, J.D.; Falson, P.; le Maire, M.; Tanner, M.J. Functional cell surface expression of the anion transport domain of human red cell band 3 (AE1) in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1996, 93, 12245–12250. [Google Scholar] [CrossRef]
- McFarland, J. The nephelometer: An instrument for estimating the number of bacteria in suspensions used for calculating the opsonic index and for vaccines. J. Am. Med Assoc. 1907, XLIX, 1176–1178. [Google Scholar] [CrossRef]
- Chowdhury, M.; Li, C.F.; He, Z.; Lu, Y.; Liu, X.S.; Wang, Y.F.; Ip, Y.T.; Strand, M.R.; Yu, X.Q. Toll family members bind multiple Spatzle proteins and activate antimicrobial peptide gene expression in Drosophila. J. Biol. Chem. 2019, 294, 10172–10181. [Google Scholar] [CrossRef] [PubMed]
- Marsano, R.M.; Giordano, E.; Messina, G.; Dimitri, P. A New Portrait of Constitutive Heterochromatin: Lessons from Drosophila melanogaster. Trends Genet. 2019, 35, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R.R. The ecology of insect-yeast relationships and its relevance to human industry. Proc. Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Tomkiel, J.; Pimpinelli, S.; Sandler, L. Rescue from the abnormal oocyte maternal-effect lethality by ABO heterochromatin in Drosophila melanogaster. Genetics 1991, 128, 583–594. [Google Scholar]
- Miskei, M.; Adam, C.; Kovacs, L.; Karanyi, Z.; Dombradi, V. Molecular evolution of phosphoprotein phosphatases in Drosophila. PLoS ONE 2011, 6, e22218. [Google Scholar] [CrossRef]
- Carvalho, A.B.; Vicoso, B.; Russo, C.A.; Swenor, B.; Clark, A.G. Birth of a new gene on the Y chromosome of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2015, 112, 12450–12455. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards Suite: From Gene Data Mining to Disease Genome Sequence Analyses. Curr. Protoc. Bioinform. 2016, 54, 1–30. [Google Scholar] [CrossRef]
- Carvalho, A.B.; Dobo, B.A.; Vibranovski, M.D.; Clark, A.G. Identification of five new genes on the Y chromosome of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2001, 98, 13225–13230. [Google Scholar] [CrossRef]
- Adám, C.; Henn, L.; Miskei, M.; Erdélyi, M.; Friedrich, P.; Dombrádi, V. Conservation of male-specific expression of novel phosphoprotein phosphatases in Drosoph. Dev. Genes Evol. 2010, 220, 123–128. [Google Scholar] [CrossRef]
- Bartolo, G.; Gonzalez, L.O.; Alameh, S.; Valencia, C.A.; Martchenko Shilman, M. Identification of glucocorticoid receptor in Drosophila melanogaster. BMC Microbiol. 2020, 20, 161. [Google Scholar] [CrossRef] [PubMed]
- Freimoser, F.M.; Rueda-Mejia, M.P.; Tilocca, B.; Migheli, Q. Biocontrol yeasts: Mechanisms and applications. World J. Microbiol. Biotechnol. 2019, 35, 154. [Google Scholar] [CrossRef] [PubMed]
- Grimont, P.A.D.; Grimont, F.; Starr, M.P. Serratia species isolated from plants. Curr. Microbiol. 1981, 5, 317–322. [Google Scholar] [CrossRef]
- Nascimento, F.; Vicente, C.; Cock, P.; Tavares, M.; Rossi, M.; Hasegawa, K.; Mota, M. From plants to nematodes: Serratia grimesii BXF1 genome reveals an adaptation to the modulation of multi-species interactions. Microb. Genom. 2018, 4. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| D. melanogaster Y-Genes | D. melanogaster Paralog(s) | H. sapiens Ortholog(s) | ||
|---|---|---|---|---|
| Gene Name(s) (Identity %/Similarity %) | Gene Function(s) | Gene Name(s) (Identity %/Similarity %) | Gene Function(s) | |
| Pp1-Y1: Protein phosphatase 1, Y-linked 1 | Pp1-Y2 [63/75] flapwing [60/76] Pp1α-96A [60/75] Pp1-13C [59/76] Pp1-87B [59/75] | Protein serine/threonine phosphatase activity | Protein Phosphatase 1 Catalytic Subunits α, β, γ PPP1CA [57/74] PPP1CB [59/74] PPP1CC [59/75] | Serine/threonine protein phosphatase; glycogen metabolism, muscle contractibility, protein synthesis |
| kl-5: male fertility factor kl5 | Dynein heavy chain Dhc93AB [61/76] | Ciliary and flagellar motor proteins | Dynein Axonemal Heavy Chain 9 DNAH9 [58/74] | Force generating protein of respiratory cilia |
| CG3339 [55/71] | Unknown | Dynein Axonemal Heavy Chain 17 DNAH17 [59/75] | Force generating protein in sperm flagellum | |
| PRY: polycystine-related-Y | CG30048 [26/44] | Unknown | Polycystin 1 Like 3, Transient Receptor Potential Channel Interacting PKD1L3 [20/36] | Calcium channel component |
| CG42685 [24/42] | Predicted calcium channel activity | |||
| kl-3: male fertility factor kl3 | CG9492 [52/69] | Unknown | Dynein Axonemal Heavy Chain 5 DNAH5 [51/69] | Force generating protein of respiratory cilia |
| Dynein heavy chain Dhc93AB [32/52] Dhc36C [32/51] | Ciliary and flagellar motor proteins | Dynein Axonemal Heavy Chain 8 DNAH8 [50/68] | Force generating protein in sperm flagellum | |
| kl-2: male fertility factor kl2 | Dynein heavy chain Dhc36C [35/54] Dhc16F [35/55] | Ciliary and flagellar motor proteins | Dynein Axonemal Heavy Chain 2 DNAH5 [33/53] | Force generating protein of respiratory cilia and in sperm flagellum |
| ARY: Aldehyde reductase Y | CG10638 [49/67] | Unknown | Aldo-Keto Reductase Family 1 Member B10 AKR1B10 [40/61] | Catalyzes Nicotinamide adenine dinucleotide phosphate NADPH-dependent reduction |
| Akr1B [40/59] | Aldo-keto reductase 1B | |||
| PPr-Y | TbCMF46 [61/80] | Unknown | Dynein Regulatory Complex Subunit 3 DRC3 [31/51] | Key regulator of ciliary and flagellar motility |
| WDY WD40 Y | CG34164 [60/73] | Unknown | EF-Hand Calcium Binding Domain 8 EFCAB8 [23/39] | Calcium ion binding |
| FDY: flagrante delicto | vig2 [93/94] | heterochromatin organization, histone H3-K9 methylation and chromatin silencing regulation | Serpin Family E Member 1 SERBP1 [44/62] | inhibitor of fibrinolysis |
| Hyaluronan Binding Protein 4 HABP4 [33/50] | Regulates transcription, pre-mRNA splicing and mRNA translation | |||
| Pp1-Y2: Protein phosphatase 1, Y-linked 2 | Pp1α-96A [75/88] Flw [76/89] Pp1-87B [74/87] Pp1-13C [73/86] Pp1-Y1 [63/75] | Protein serine/threonine phosphatase activity | Protein Phosphatase 1 Catalytic Subunits α, β, γ PPP1CA [57/74] PPP1CB [59/74] PPP1CC [59/75] | Serine/threonine protein phosphatase; glycogen metabolism, muscle contractility, protein synthesis |
| ORY: Occludin-Related Y | CG6059 [34/58] CG5882 [26/51] | Unknown | Cilia and Flagella Associated Protein 58 CFAP58 [29/54] | Associated with melanoma |
| CCY: Coiled-Coils Y | CG31161 [52/71] | Unknown | Cortactin-binding protein 2 isoform X7 CTTNBP2 [24/48] | Interacts with a central regulator of the actin cytoskeleton |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartolo, G.; Gonzalez, L.O.; Levitin, A.; Martchenko Shilman, M. Drosophila melanogaster Y Chromosome Genes Affect Male Sensitivity to Microbial Infections. Insects 2021, 12, 30. https://doi.org/10.3390/insects12010030
Bartolo G, Gonzalez LO, Levitin A, Martchenko Shilman M. Drosophila melanogaster Y Chromosome Genes Affect Male Sensitivity to Microbial Infections. Insects. 2021; 12(1):30. https://doi.org/10.3390/insects12010030
Chicago/Turabian StyleBartolo, Gloria, Leandra O. Gonzalez, Anastasia Levitin, and Mikhail Martchenko Shilman. 2021. "Drosophila melanogaster Y Chromosome Genes Affect Male Sensitivity to Microbial Infections" Insects 12, no. 1: 30. https://doi.org/10.3390/insects12010030
APA StyleBartolo, G., Gonzalez, L. O., Levitin, A., & Martchenko Shilman, M. (2021). Drosophila melanogaster Y Chromosome Genes Affect Male Sensitivity to Microbial Infections. Insects, 12(1), 30. https://doi.org/10.3390/insects12010030