Environmental Display Can Buffer the Effect of Pesticides on Solitary Bees



Abstract
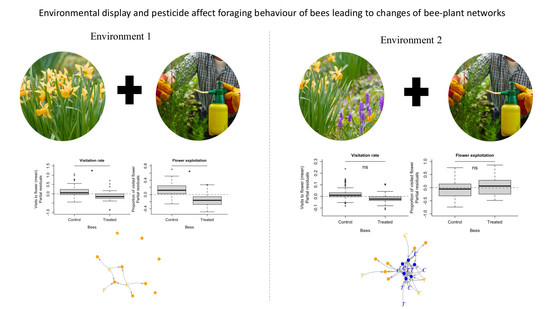
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. Bee Species
2.2. Stressor
2.3. Training Phase
2.4. Experimental Displays
2.4.1. Environment Type 1-Experiment with Flowers of a Single Colour Hanging on the Sides of the Arena
2.4.2. Environment Type 2- Experiment with Flowers of Different Colours Set at the Bottom of the Arena
2.5. Analysis of Foraging Behaviour
2.6. Analysis of Nectar Uptake
2.7. Effect of Training Flower on Environment Type 2
2.8. Network Analysis
3. Results
3.1. Environment Type 1
3.2. Environment Type 2
3.3. Nectar Uptake
3.4. Individual Specialization and Interaction Network Structures
4. Discussion
4.1. Environment Type Effect on Foraging Behaviour of Bees
4.2. Nectar Uptake and Effect of Training Flower
4.3. Individual Specialization and Structural Network Metrics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Seibold, S.; Gossner, M.M.; Simons, N.K.; Blüthgen, N.; Müller, J.; Ambarlı, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, J.C.; et al. Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 2019, 574, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Jucker, C.; Lupi, D. Exotic Insects in Italy: An Overview on their Environmental Impact, the Importance of Biological Interactions in the Study of Biodiversity; Pujol, J.L., Ed.; InTech: Rijeka, Croatia, 2011; pp. 51–74. ISBN 978-953-307-751-2. [Google Scholar] [CrossRef]
- MacDougall, A.S.; McCann, K.S.; Gellner, G.; Turkington, R. Diversity loss with persistent human disturbance increases vulnerability to ecosystem collapse. Nature 2013, 494, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Cordero-Rivera, A. Behavioral diversity (ethodiversity): A neglected level in the study of biodiversity. Front. Eco. Evol. 2017, 5, 1–7. [Google Scholar] [CrossRef]
- Grue, C.E.; Gardner, S.C.; Gibert, P. Behavioural Ecotoxicology; Dell’Omo, G., Ed.; John Wiley & Sons: New York, NY, USA, 2002. [Google Scholar]
- Valido, A.; Rodríguez-Rodríguez, M.C.; Jordano, P. Honeybees disrupt the structure and functionality of plant-pollinator networks. Sci. Rep. 2019, 9, 1. [Google Scholar] [CrossRef]
- Theodorou, P.; Albig, K.; Radzevičiūtė, R.; Settele, J.; Schweiger, O.; Murray, T.E.; Paxton, R.J. The structure of flower visitor networks in relation to pollination across an agricultural to urban gradient. Funct. Ecol. 2017, 31, 838–847. [Google Scholar] [CrossRef]
- Peterson, E.K.; Buchwalter, D.B.; Kerby, J.L.; LeFauve, M.K.; Varian-Ramos, C.W.; Swaddle, J.P. Integrative behavioral ecotoxicology: Bringing together fields to establish new insight to behavioral ecology, toxicology, and conservation. Cur. Zool. 2017, 63, 185–194. [Google Scholar] [CrossRef]
- Goldenberg, S.U.; Nagelkerken, I.; Marangon, E.; Bonnet, A.; Ferreira, C.M.; Connell, S.D. Ecological complexity buffers the impacts of future climate on marine consumers. Nat. Clim. Chang. 2018, 8, 229–233. [Google Scholar] [CrossRef]
- Shivik, J.A.; Palmer, G.L.; Gese, E.M.; Osthaus, B. Captive coyotes compared to their counterparts in the wild: Does environmental enrichment help? J. Appl. Anim. Welf. Sci. 2009, 12, 223–235. [Google Scholar] [CrossRef]
- Campderrich, I.; Nazar, F.N.; Wichman, A.; Marin, R.H.; Estevez, I.; Keeling, L.J. Environmental complexity: A buffer against stress in the domestic chick. PLoS ONE 2019, 14, e0210270. [Google Scholar] [CrossRef]
- DePasquale, C.; Neuberger, T.; Hirrlinger, A.M.; Braithwaite, V.A. The influence of complex and threatening environments in early life on brain size and behaviour. Proc. R. Soc. B 2016, 283, 20152564. [Google Scholar] [CrossRef]
- Cusser, S.; Neff, J.L.; Jha, S. Landscape context differentially drives diet breadth for two key pollinator species. Oecologia 2019, 30, 1–4. [Google Scholar] [CrossRef]
- Stanley, D.A.; Garratt, M.P.; Wickens, J.B.; Wickens, V.J.; Potts, S.G.; Raine, N.E. Neonicotinoid pesticide exposure impairs crop pollination services provided by bumblebees. Nature 2015, 528, 548. [Google Scholar] [CrossRef] [PubMed]
- Tschoeke, P.H.; Oliveira, E.E.; Dalcin, M.S.; Silveira-Tschoeke, M.C.; Sarmento, R.A.; Santos, G.R. Botanical and synthetic pesticides alter the flower visitation rates of pollinator bees in Neotropical melon fields. Environ. Pollut. 2019, 251, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Whitehorn, P.R.; Wallace, C.; Vallejo-Marin, M. Neonicotinoid pesticide limits improvement in buzz pollination by bumblebees. Sci. Rep. 2017, 7, 15562. [Google Scholar] [CrossRef] [PubMed]
- Boff, S.; Friedel, A.; Mussury, R.M.; Lenis, P.R.; Raizer, J. Changes in social behavior are induced by pesticide ingestion in a Neotropical stingless bee. Ecotox. Environ. Saf. 2018, 164, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.A.; Smith, K.E.; Raine, N.E. Bumblebee learning and memory is impaired by chronic exposure to a neonicotinoid pesticide. Sci. Rep. 2015, 5, 16508. [Google Scholar] [CrossRef]
- Muth, F.; Leonard, A.S. A neonicotinoid pesticide impairs foraging, but not learning, in free-flying bumblebees. Sci. Rep. 2019, 9, 1–3. [Google Scholar] [CrossRef]
- Sandrock, C.; Tanadini, L.G.; Pettis, J.S.; Biesmeijer, J.C.; Potts, S.G.; Neumann, P. Sublethal neonicotinoid insecticide exposure reduces solitary bee reproductive success. Agric. For. Entomol. 2014, 16, 119–128. [Google Scholar] [CrossRef]
- Jin, N.; Klein, S.; Leimig, F.; Bischoff, G.; Menzel, R. The neonicotinoid clothianidin interferes with navigation of the solitary bee Osmia cornuta in a laboratory test. J. Exp. Biol. 2015, 218, 2821–2825. [Google Scholar] [CrossRef]
- Biddinger, D.J.; Robertson, J.L.; Mullin, C.; Frazier, J.; Ashcraft, S.A.; Rajotte, E.G.; Joshi, N.K.; Vaughn, M. Comparative toxicities and synergism of apple orchard pesticides to Apis mellifera (L.) and Osmia cornifrons (Radoszkowski). PLoS ONE 2013, 8, e72587. [Google Scholar] [CrossRef]
- Hudewenz, A.; Klein, A.M. Red mason bees cannot compete with honey bees for floral resources in a cage experiment. Ecol. Evol. 2015, 5, 5049–5056. [Google Scholar] [CrossRef] [PubMed]
- Westrich, P. Die Wildbienen Baden-Wüttembergs; Eugen Ulmer: Stuttgart, Germany, 1989. [Google Scholar]
- Gruber, B.; Eckel, K.; Everaars, J.; Dormann, C.F. On managing the red mason bee (Osmia bicornis) in apple orchards. Apidologie 2011, 42, 564–576. [Google Scholar] [CrossRef]
- Gresty, C.E.; Clare, E.; Devey, D.S.; Cowan, R.S.; Csiba, L.; Malakasi, P.; Lewis, O.T.; Willis, K.J. Flower preferences and pollen transport networks for cavity-nesting solitary bees: Implications for the design of agri-environment schemes. Ecol. Evol. 2018, 8, 7574–7587. [Google Scholar] [CrossRef]
- Raw, A. The biology of the solitary bee Osmia rufa (L.) (Megachilidae). Trans. R. Entomol. Soc. Lond. 1972, 124, 213–229. [Google Scholar] [CrossRef]
- Seidelmann, K.; Bienasch, A.; Pröhl, F. The impact of nest tube dimensions on reproduction parameters in a cavity nesting solitary bee, Osmia bicornis (Hymenoptera: Megachilidae). Apidologie 2016, 47, 114–122. [Google Scholar] [CrossRef]
- Artz, D.R.; Pitts-Singer, T.L. Effects of fungicide and adjuvant sprays on nesting behavior in two managed solitary bees, Osmia lignaria and Megachile rotundata. PLoS ONE 2015, 10, e0135688. [Google Scholar] [CrossRef] [PubMed]
- Sgolastra, F.; Arnan, X.; Cabbri, R.; Isani, G.; Medrzycki, P.; Teper, D.; Bosch, J. Combined exposure to sublethal concentrations of an insecticide and a fungicide affect feeding, ovary development and longevity in a solitary bee. Proc. R. Soc. B 2018, 285, 20180887. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, B.D.; Lois, A.N.; Guédot, C. Effect of fungicide on pollen foraging by honeybees (Hymenoptera: Apidae) in cranberry differs by fungicide type. J. Econ. Entomol. 2018, 112, 499–503. [Google Scholar] [CrossRef]
- European Food Safety Authority. Conclusion on the peer review of the pesticide risk assessment of the active substance fenbuconazole. EFSA J. 2010, 8, 1558. [Google Scholar] [CrossRef]
- Gathmann, A.; Tscharntke, T. Foraging ranges of solitary bees. J. Anim. Ecol. 2002, 71, 757–764. [Google Scholar] [CrossRef]
- Akter, A.; Biella, P.; Klecka, J. Effects of small-scale clustering of flowers on pollinator foraging behaviour and flower visitation rate. PLoS ONE 2017, 12, e0187976. [Google Scholar] [CrossRef]
- Brito, V.L.; Weynans, K.; Sazima, M.; Lunau, K. Trees as huge flowers and flowers as oversized floral guides: The role of floral colour change and retention of old flowers in Tibouchina pulchra. Front. Plant Sci. 2015, 6, 362. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.S.; Masek, P. Multisensory integration of colours and scents: Insights from bees and flowers. J. Comp. Physiol. A 2014, 200, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.; Kemp, W.P. Effect of wintering duration and temperature on survival and emergence time in males of the orchard pollinator Osmia lignaria (Hymenoptera: Megachilidae). Environ. Entomol. 2003, 32, 711–716. [Google Scholar] [CrossRef]
- Bosch, J.; Sgolastra, F.; Kemp, W.P. Timing of eclosion affects diapause development, fat body consumption and longevity in Osmia lignaria, a univoltine, adult-wintering solitary bee. J. Insect Physiol. 2010, 56, 1949–1957. [Google Scholar] [CrossRef]
- Sgolastra, F.; Bosch, J.; Molowny-Horas, R.; Maini, S.; Kemp, W.P. Effect of temperature regime on diapause intensity in an adult-wintering Hymenopteran with obligate diapause. J. Insect Physiol. 2010, 56, 185–194. [Google Scholar] [CrossRef]
- Dmochowska, K.; Giejdasz, K.; Fliszkiewicz, M.; Żółtowska, K. Prolonged postdiapause: Influence on some indicators of carbohydrate and lipid metabolism of the red mason bee, Osmia rufa. J. Insect Sci. 2013, 13, 77. [Google Scholar] [CrossRef]
- Bertrand, C.; Eckerter, P.W.; Ammann, L.; Entling, M.H.; Gobet, E.; Herzog, F.; Mestre, L.; Tinner, W.; Albrecht, M. Seasonal shifts and complementary use of pollen sources by two bees, a lacewing and a ladybeetle species in European agricultural landscapes. J. Appl. Ecol. 2019, 56, 2431–2442. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. Int. J. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Dormann, C.F.; Fründ, J.; Blüthgen, N.; Gruber, B. Indices, graphs and null models: Analyzing bipartite ecological networks. Open Ecol. J. 2009, 2, 7–24. [Google Scholar] [CrossRef]
- Dehling, D.M. The Structure of Ecological Networks. In Ecological Networks in the Tropics; Springer: Cham, Switzerland, 2018; pp. 29–42. [Google Scholar]
- Jordano, P. Patterns of mutualistic interactions in pollination and seed dispersal: Connectance, dependence asymmetries, and coevolution. Am. Nat. 1987, 129, 657–677. [Google Scholar] [CrossRef]
- Guimarães, P.R., Jr.; Guimarães, P.R. Improving the analyses of nestedness for large sets of matrices. Environ. Model. Softw. 2006, 21, 1512–1513. [Google Scholar] [CrossRef]
- Dormann, C.F.; Gruber, B.; Fründ, J. Introducing the Bipartite Package: Analysing Ecological Networks. Interaction 2008, 1, 0.2413793. [Google Scholar]
- Blüthgen, N.; Menzel, F.; Blüthgen, N. Measuring specialization in species interaction networks. BMC Ecol. 2006, 6, 9. [Google Scholar] [CrossRef]
- Blüthgen, N.; Menzel, F.; Hovestadt, T.; Fiala, B.; Blüthgen, N. Specialization, constraints, and conflicting interests in mutualistic networks. Cur. Biol. 2007, 17, 341–346. [Google Scholar] [CrossRef]
- Hartig, F. Package “DHARMa”: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. An R Package. 2016. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 3 July 2020).
- Harvey, J.A.; Heinen, R.; Armbrecht, I.; Basset, Y.; Baxter-Gilbert, J.H.; Bezemer, T.M.; Böhm, M.; Bommarco, R.; Borges, P.A.; Cardoso, P.; et al. International scientists formulate a roadmap for insect conservation and recovery. Nat. Ecol. Evol. 2020, 6, 1–3. [Google Scholar] [CrossRef]
- Tosi, S.; Nieh, J.C. Lethal and sublethal synergistic effects of a new systemic pesticide, flupyradifurone (Sivanto®), on honeybees. Proc. R. Soc. B 2019, 286, 20190433. [Google Scholar] [CrossRef]
- Sattler, C.; Gianuca, A.T.; Schweiger, O.; Franzén, M.; Settele, J. Pesticides and land cover heterogeneity affect functional group and taxonomic diversity of arthropods in rice agroecosystems. Agric. Ecos. Environ. 2020, 297, 106927. [Google Scholar] [CrossRef]
- Dubuc-Messier, G.; Réale, D.; Perret, P.; Charmantier, A. Environmental heterogeneity and population differences in blue tits personality traits. Behav. Ecol. 2017, 28, 448–459. [Google Scholar] [CrossRef]
- Hatjina, F.; Papaefthimiou, C.; Charistos, L.; Dogaroglu, T.; Bouga, M.; Emmanouil, C.; Arnold, G. Sublethal doses of imidacloprid decreased size of hypopharyngeal glands and respiratory rhythm of honeybees in vivo. Apidologie 2013, 44, 467–480. [Google Scholar] [CrossRef]
- Karise, R.; Muljar, R.; Smagghe, G.; Kaart, T.; Kuusik, A.; Dreyersdorff, G.; Williams, I.H.; Mänd, M. Sublethal effects of kaolin and the biopesticides Prestop-Mix and BotaniGard on metabolic rate, water loss and longevity in bumble bees (Bombus terrestris). J. Pest Sci. 2016, 89, 171–178. [Google Scholar] [CrossRef]
- Muljar, R.; Karise, R.; Viik, E.; Kuusik, A.; Williams, I.; Metspalu, L.; Hiiesaar, K.; Must, A.; Luik, A.; Mänd, M. Effects of Fastac 50 EC on bumble bee Bombus terrestris L. respiration: DGE disappearance does not lead to increasing water loss. J. Insect Physiol. 2012, 58, 1469–1476. [Google Scholar] [CrossRef] [PubMed]
- Muchhala, N.; Serrano, D. The complexity of background clutter affects nectar bat use of flower odor and shape cues. PLoS ONE 2015, 10, e0136657. [Google Scholar] [CrossRef] [PubMed]
- Boff, S.; Melo-de-Pinna, G.F.; Pott, A.; Araujo, A.C. High visitation rate of oil bees may increase pollination efficiency of Couepia uiti in Pantanal wetland. Apidologie 2018, 49, 747–758. [Google Scholar] [CrossRef]
- Breed, M.F.; Ottewell, K.M.; Gardner, M.G.; Marklund, M.H.; Dormontt, E.E.; Lowe, A.J. Mating patterns and pollinator mobility are critical traits in forest fragmentation genetics. Heredity 2015, 115, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Halinski, R.; Garibaldi, L.A.; dos Santos, C.F.; Acosta, A.L.; Guidi, D.D.; Blochtein, B. Forest fragments and natural vegetation patches within crop fields contribute to higher oilseed rape yields in Brazil. Agric. Syst. 2020, 180, 102768. [Google Scholar] [CrossRef]
- Kaluza, B.F.; Wallace, H.M.; Heard, T.A.; Minden, V.; Klein, A.; Leonhardt, S.D. Social bees are fitter in more biodiverse environments. Sci. Rep. 2018, 8, 12353. [Google Scholar] [CrossRef]
- Vaudo, A.D.; Tooker, J.F.; Patch, H.M.; Biddinger, D.J.; Coccia, M.; Crone, M.K.; Fiely, M.; Francis, J.S.; Hines, H.M.; Hodges, M.; et al. Pollen protein: Lipid macronutrient ratios may guide broad patterns of bee species floral preferences. Insects 2020, 11, 132. [Google Scholar] [CrossRef]
- LoCascio, G.M.; Aguirre, L.; Irwin, R.E.; Adler, L.S. Pollen from multiple sunflower cultivars and species reduces a common bumblebee gut pathogen. R. Soc. Open Sci. 2019, 6, 190279. [Google Scholar] [CrossRef]
- Hesselbach, H.; Scheiner, R. Effects of the novel pesticide flupyradifurone (Sivanto) on honeybee taste and cognition. Sci. Rep. 2018, 8, 4954. [Google Scholar] [CrossRef]
- Goñalons, C.M.; Farina, W.M. Effects of sublethal doses of imidacloprid on young adult honeybee behaviour. PLoS ONE 2015, 10, e0140814. [Google Scholar] [CrossRef] [PubMed]
- Giurfa, M.; Nunez, J.; Chittka, L.; Menzel, R. Colour preferences of flower-naive honeybees. J. Comp. Physiol. 1995, 177, 247–259. [Google Scholar] [CrossRef]
- Martínez-Núñez, C.; Manzaneda, A.J.; Lendínez, S.; Pérez, A.J.; Ruiz-Valenzuela, L.; Rey, P.J. Interacting effects of landscape and management on plant–solitary bee networks in olive orchards. Funct. Ecol. 2019, 33, 2316–2326. [Google Scholar] [CrossRef]
- Blüthgen, N.; Klein, A.M. Functional complementarity and specialisation: The role of biodiversity in plant–pollinator interactions. Basic Appl. Ecol. 2011, 12, 282–291. [Google Scholar] [CrossRef]
- Rundlöf, M.; Lundin, O.; Bommarco, R. Annual flower strips support pollinators and potentially enhance red clover seed yield. Ecol. Evol. 2018, 8, 7974–7985. [Google Scholar] [CrossRef]
- Krishna, A.; Guimarães, P.R., Jr.; Jordano, P.; Bascompte, J. A neutral-niche theory of nestedness in mutualistic networks. Oikos 2008, 117, 1609–1618. [Google Scholar] [CrossRef]
- Pisula, W.; Modlinska, K.; Chrzanowska, A. Behavioural response to the environmental changes of various types in Lister-Hooded male rats. Sci. Rep. 2019, 9, 7111. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boff, S.; Raizer, J.; Lupi, D. Environmental Display Can Buffer the Effect of Pesticides on Solitary Bees. Insects 2020, 11, 417. https://doi.org/10.3390/insects11070417
Boff S, Raizer J, Lupi D. Environmental Display Can Buffer the Effect of Pesticides on Solitary Bees. Insects. 2020; 11(7):417. https://doi.org/10.3390/insects11070417
Chicago/Turabian StyleBoff, Samuel, Josué Raizer, and Daniela Lupi. 2020. "Environmental Display Can Buffer the Effect of Pesticides on Solitary Bees" Insects 11, no. 7: 417. https://doi.org/10.3390/insects11070417
APA StyleBoff, S., Raizer, J., & Lupi, D. (2020). Environmental Display Can Buffer the Effect of Pesticides on Solitary Bees. Insects, 11(7), 417. https://doi.org/10.3390/insects11070417