1. Introduction
Carrot flowers are unattractive to honey bees (
Apis mellifera Linnaeus), which often seek out other available forage [
1]. Hybrid or cytoplasmically male-sterile (CMS) cultivars are particularly unattractive, as they do not provide pollen [
2,
3,
4]. Such cultivars are also believed to be inferior in their morphology, scent, and nectar production to male fertile (MF) plants [
5].
Several studies have reported species-selective pollinator visitation to hybrid carrot seed crops including pollinators such as
A. mellifera [
2,
3,
6,
7,
8], flies [
9,
10],
Megachile bees [
11], and other native bees [
12], but there appear to be no reports on carrot pollinator assemblage level visitation to different CMS cultivars, despite strong indications of differences in pollinator attraction among cultivars [
13,
14,
15].
Generally, there are many visual and olfactory floral traits, including morphology, colour, nectar composition, and floral volatiles, which separately or collectively make one flower easily distinguishable from another to a pollinator [
16,
17,
18]. Furthermore, some pollinators such as honey bees are opportunistic foragers, whose tendencies for innate responses can be altered by learning, mainly via olfactory conditioning [
19,
20]. Manipulation of the chemistry in the colony has also been investigated, where the treatment of honey bee colonies with brood pheromone increased the foraging and yield of carrot seed in nearby fields [
21].
The nectar quality and availability are of almost universal importance to all insect foragers, as it is one reward that encourages insects to return to a flower [
22,
23]. Most honey bee-pollinated plants have high levels of sucrose in their nectar [
24,
25]. Percival analysed the nectar content of 889 species of angiosperm and concluded that plants pollinated by honey bees tend to have high disaccharide/monosaccharide (fructose and glucose) ratios [
26]. Nonetheless, crops well-visited by bees showed a relatively balanced distribution of sugars, and it was concluded that bees show no direct preference for sucrose-dominant nectars in the field. However, it is well known that honey bees display a genetic propensity for proboscis response to sucrose [
27,
28].
The colour spectrum experienced by various insects differs from that seen by humans, and the colour perception in many Hymenoptera extends into the ultraviolet range [
29]. Therefore, different levels of ultraviolet reflectance may affect the attraction of honey bees and other insects to flowers. Furthermore, carrots have flowers grouped together in large umbels, which provide a large, highly visible landing platform. As umbel size is known to vary between cultivars and individuals, with the primary or terminal umbel being the largest of the umbels on each plant, typically between 100 and 150 mm in diameter [
30], both umbel size and UV absorbance may be important for pollinator attraction.
To date, the few studies that have been conducted on the floral traits that attract insects to different carrot flowers have mainly focussed on floral volatiles from a limited number of cultivars [
7,
8]. In this study, insect visitation to 17 cultivars of flowering CMS carrots is recorded to determine whether some cultivars attract more insect visitors than others and whether there is variation in the visitation frequency by some of the more abundant visitor groups. The aims are to identify (1) how rates of insect visitation vary to different CMS carrot genotypes or cultivars, and (2) how visitation by honey bees and other insects is related to key floral traits other than floral volatiles, including ultraviolet reflectance, nectar production, and the size of inflorescences.
2. Materials and Methods
2.1. Insect Visitation to Seventeen CMS Cultivars
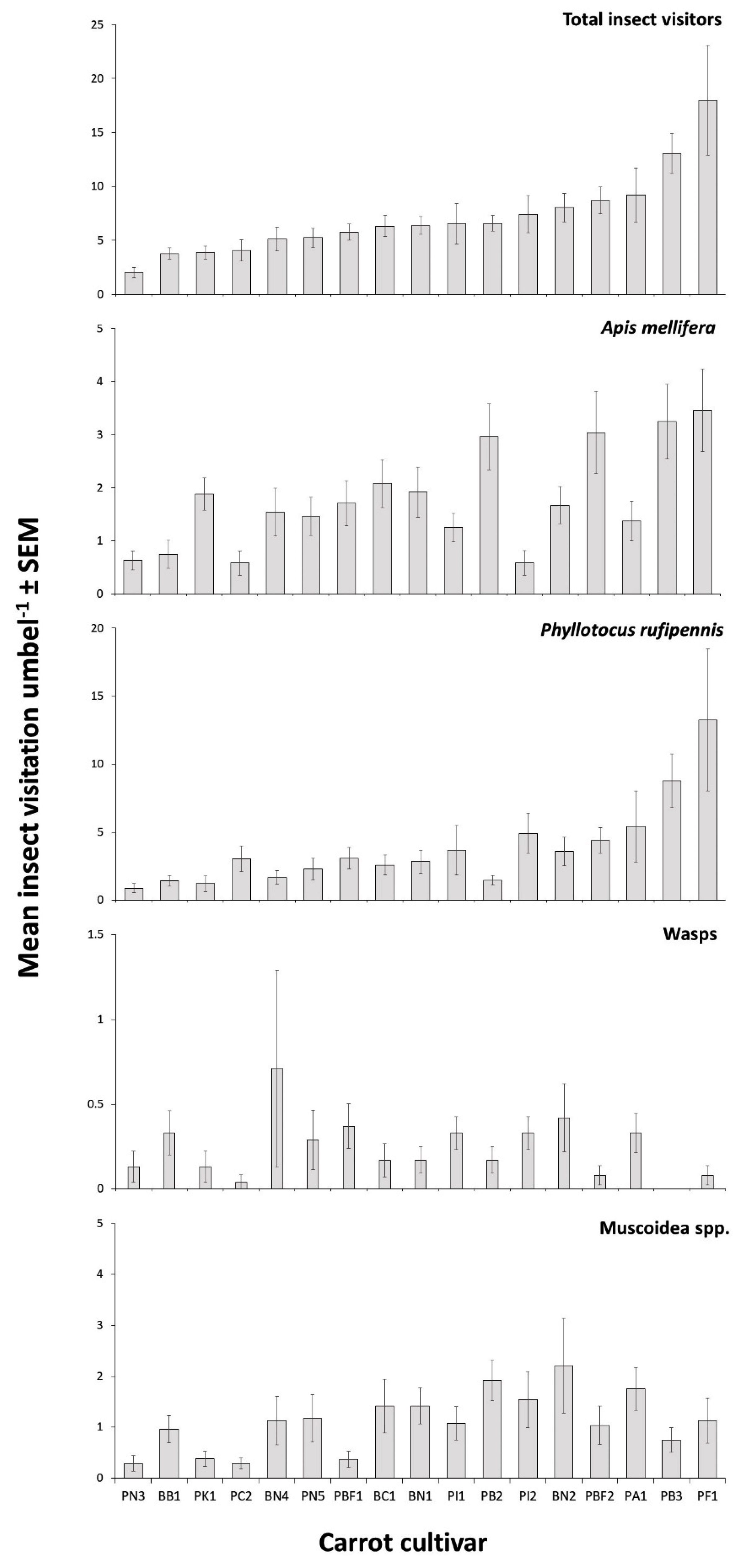
In season 1, a comparison of insect visitation to 17 cultivars (lines) of CMS carrots was conducted in field trial A at Bejo Seeds in Southern Tasmania (42.704° S, 147.445° E), on four days between 14 and 24 December 2002 (24 umbels per cultivar, 408 total umbels observed). The carrot cultivars were planted for a commercial grow-out trial in a completely randomised design (
Figure S1). Seventeen carrot lines were allocated to these plots. Of the additional seven plots, six contained duplicated carrot lines not used in this study, and one was empty. Each plot was a 5 m × 1.6 m bed containing three rows of carrots. The rows of carrots in each bed were 0.3 m distant from each other with a 60 cm space between each of the different carrot lines. No managed European honey bee hives were placed at the trial site to prevent positional biasing of the results.
The 17 carrot cultivars were of four different CMS types (
Table 1). The two main groups were: brown anther (having non-productive anthers, which are brown in colour) and petaloid (having petal-like structures instead of anthers) with petaloid colour (green, white, or purple) used to subdivide this group into a further three CMS types. There were five brown anther cultivars and three petaloid cultivars: purple (1 cultivar), white (7 cultivars), and green (9 cultivars). All root types were represented by one or more cultivar: Nantes (5 cultivars, n = 120 umbels), Berlicumer (3 cultivars, n = 72 umbels), Berlicumer/Flakee (2 cultivars, n = 48 umbels), Chantenay (2 cultivars, n = 48 umbels), Imperator (2 cultivars, n = 48 umbels), Amsterdam (1 cultivar, n = 24 umbels), Flakee (1 cultivar, n = 24 umbels), and ABK/N (1 cultivar, n = 24 umbels). Details of the cultivars used, and the experimental ID codes allocated to each can be found in
Table 1.
Receptive carrot umbels from the 17 cultivars were randomly selected with all selected umbels having > 40% receptive flowers. Each of the four days was divided into three 2-hour observation periods; being 09:00–11:00, 11:00–13:00 and 13:00–15:00. All four observation days were calm and sunny, air temperatures were above 15 °C at 09:00, and the maximum temperature varied between 20 and 27 °C. From each cultivar, two observers each watched two umbels, which were randomly selected and observed for five minutes within each of these time periods. Cultivars were observed in a pre-determined random order. Prior to the commencement of each 5-minute observation period, the number and type of insects that were already present on each of the selected umbels were recorded.
Each time an insect alighted on one of the umbels under observation, it was recorded according to its classification. No distinction was made between insects alighting for the first time on an umbel or returning to an umbel after departing. Each umbel of a pair was scored separately. To avoid affecting the behaviour of the insects under observation, and to ensure not to cast a shadow over the umbel, the observer remained at least 1 m away from the umbels under observation. The insect groupings recorded were European honey bee (A. mellifera), nectar scarabs (predominantly Phyllotocus rufipennis (Boisduval)), bee flies (Bombyliidae, predominantly Comptosia ocellata (Newman)), flies (Muscoidea predominantly Calliphoridae), native bees (Halictidae and Colletidae), soldier beetles (Cantharidae, specifically Chauliognathus lugubris (Fabricius)), wasps, and other, which comprised species that did not fall within the aforementioned groupings.
In field trial A, visitation data for cultivars were also grouped for analysis based on CMS type (brown anther or petaloid), root type, and flower colour to assess the effects of floral morphological differences on insect visitation. Comparisons were undertaken using non-parametric Kruskal–Wallis tests, Mann–Whitney U tests, and Friedman tests, depending on the numbers of cultivars being compared for all insects pooled together and for each insect grouping using SPSS® 17.0. Where Friedman’s and Kruskal–Wallis tests were undertaken, multiple Wilcoxon signed ranks tests were then used to determine which cultivars differed from one another. P values were adjusted using a sequential Bonferroni procedure to control for an increased chance of type-I-error resulting from multiple comparisons.
2.2. Floral traits and insect visitation
Ultraviolet reflectance, nectar sugar levels, and umbel diameter are all possible floral traits that may influence insect visitation to an umbel. To examine whether these traits varied among cultivars, field trial B was undertaken. In this trial, umbel insect visitation was recorded in the field, and then, the floral traits of those umbels were assessed. This was performed by selecting six petaloid cultivars that were both commercially available and deemed either the most attractive (PA1, PB3, PF1) or least attractive (PN3, PC2, PBF1) to insect visitors during field trial A.
Field site B was planted in a randomised complete block design comprising five blocks containing each of the six cultivars (
Figure S2). One experimental plot of cultivar PA1 failed to germinate, leaving only four blocks containing PA1 at flowering. Each block consisted of a single bed measuring 10 m × 0.8 m containing three rows of each cultivar. A single bed (3 rows) of one MF cultivar (MX1) was planted in between each of the experimental CMS cultivars to separate the plots and to stimulate pollinating insects, as is the practice in commercial crops. Field trial B was conducted from 20 January to 16 February 2004, at the University Farm (42.797° S, 147.426° E), Southern Tasmania. The methods for the collection and analysis of data for each floral trait are presented in the following sections.
2.2.1. Insect Activity and Diameter
As in trial A, insect visitation was recorded in trial B, but in this trial, the diameter was also measured for each umbel observed. Observations were done each day across all cultivars on seven separate days (n = 160–214 observations per cultivar). Insect activity on each umbel was observed for five minutes. Cultivars were observed in a pre-determined random order. Each time an insect alighted on one of the umbels under observation, it was recorded according to its classification as categorised in trial A. Following observations for insect visits, the diameter of each umbel was measured in the field, by holding the hand flat, underneath the umbel, with fingers on either side of the stem to fully extend the surface area. Then, a 30-cm ruler was used to measure the umbels to the nearest 0.5 cm. Overall, 1198 umbels were observed for insect visits and measured for umbel diameter. Both correlation and direct logistic regression were used to assess the impact that cultivar and umbel diameter had on insect visitation by each grouping, with visitation recorded as visited or not visited.
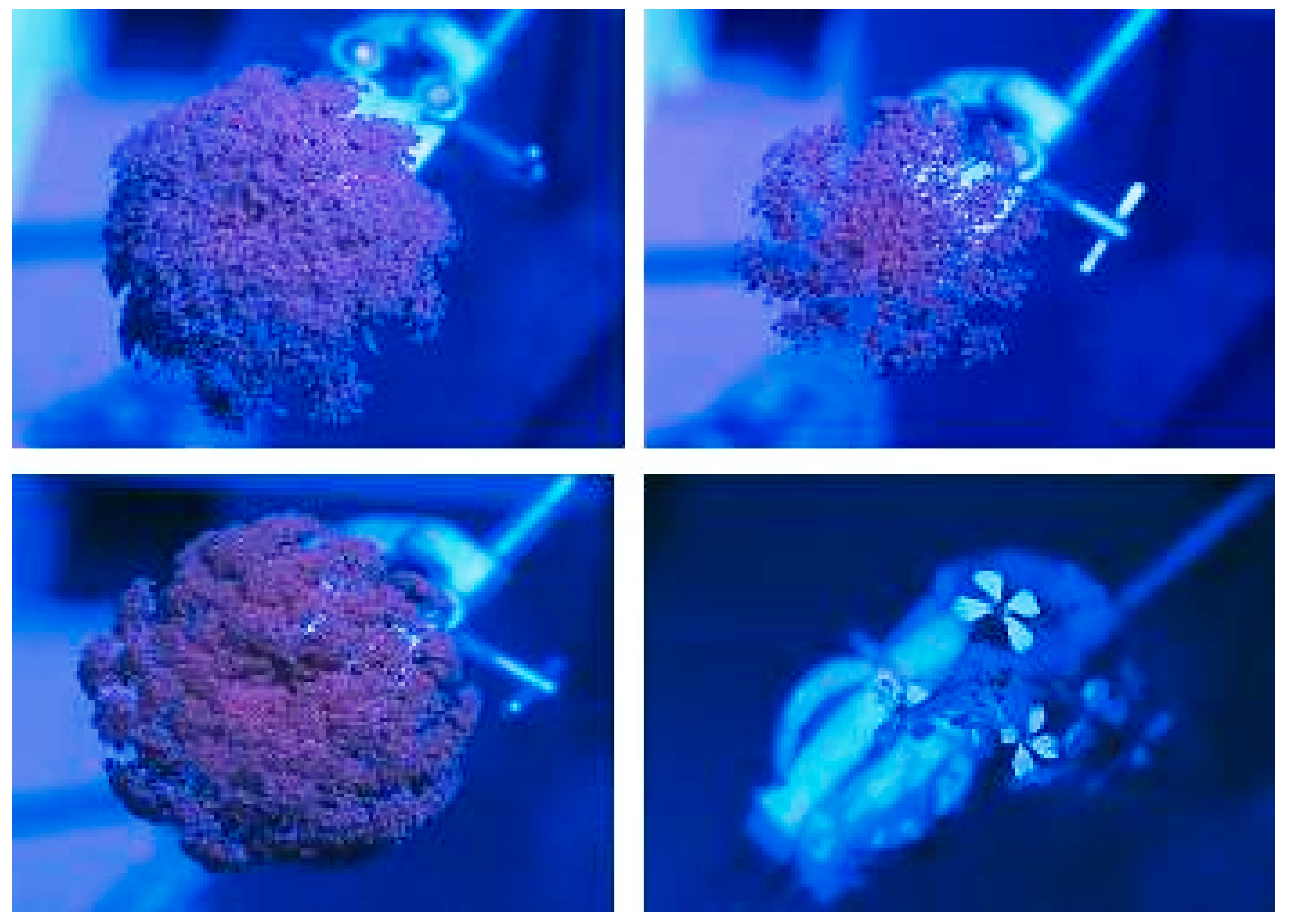
2.2.2. Ultraviolet Reflectance
Photographs of ultraviolet reflectance were taken at field site B. Seven umbels were subsampled per cultivar, removed, and placed in a box. The box sheltered the sample from any wind that may have caused movement of the umbels during photography. Then, the stalk of each umbel was placed directly into a small hole in a black card for photographing. Photographs were taken using a Pentax SP1000 SLR camera (F 5.6) (Tokyo, Japan) and 35 mm Fuji RTP II film (Fujifilm, Tokyo, Japan). This film has enhanced sensitivity to ultraviolet light. A Hoya U360 (ultraviolet bandpass) (Hoya, Tokyo, Japan) filter was used. Illumination was provided by natural sunlight and a Yuzo DC2814 ring flash (Fujifilm, Tokyo, Japan). Three flashes were found to provide optimum exposure to detect ultraviolet reflectance. The camera was mounted on a tripod and focussed prior to attaching the ultraviolet filter. The box and camera were placed at a fixed distance from each other so that refocussing was not required.
Follow-up confirmation of field photographic results was performed in the laboratory on the cut umbels using a Zeiss Tessovar macro lens system (Carl Zeiss, Oberkochen, Germany) linked to a Watec WAT202D digital colour camera (Watec, Yamagata, Japan). Images were viewed using Fly Video software on a PC fitted with a Fly Video ’98 video capture card (Animation Technologies, Hsin Tien City, Taiwan).
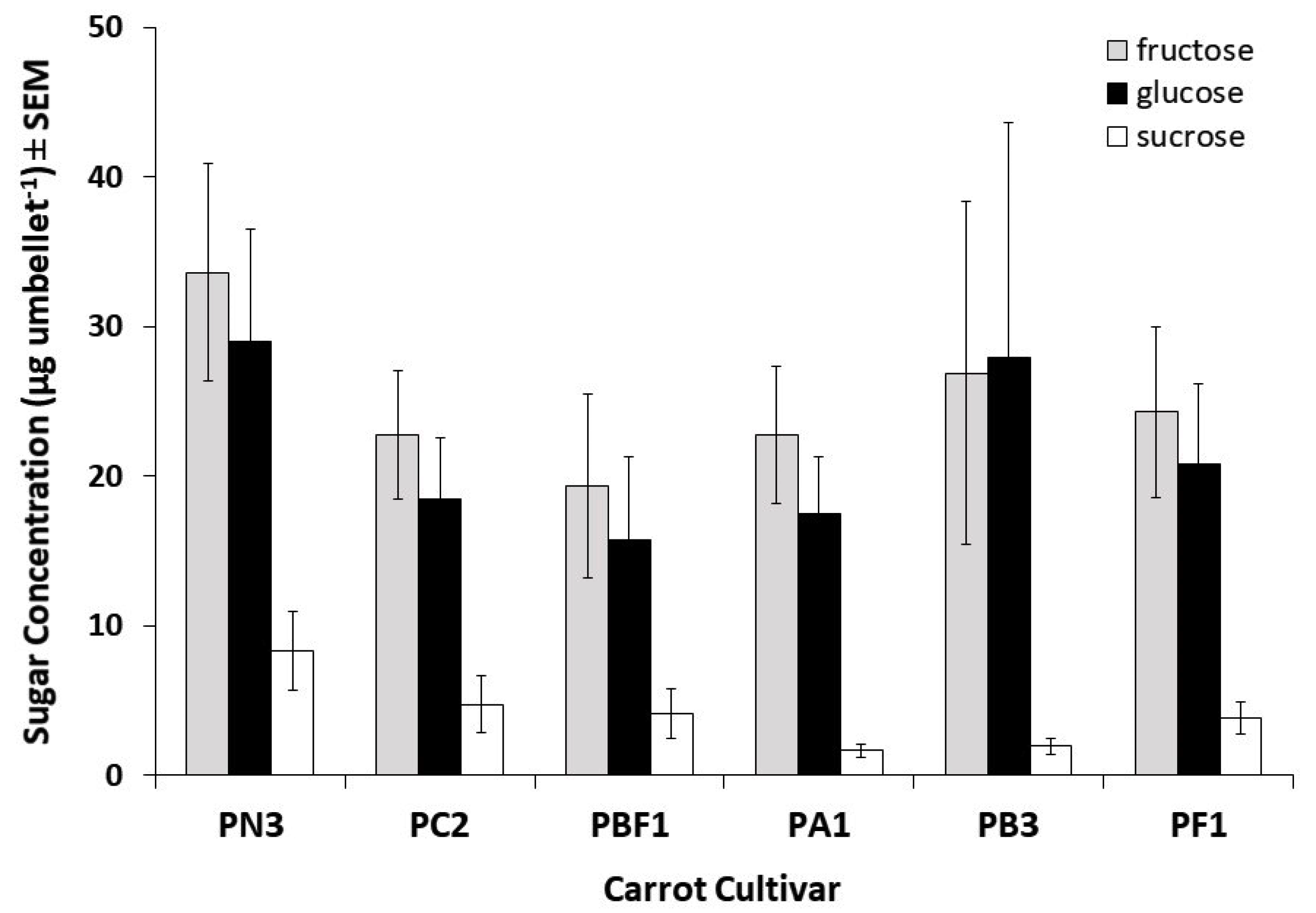
2.2.3. Soluble Sugar Analysis of Nectar
Soluble sugar analysis was undertaken in trial B immediately following photography for UV reflectance. An extraction technique was devised that could be used under windy field conditions by modifying a method by Manetas and Petropoulou [
31]:
A carrot umbel consists of four whorls of umbellets. It proved more practical to sample nectar from umbellets rather than entire umbels, with preliminary experiments revealing that dipping carrot umbellets in distilled water 20 times removed over 88% of the nectar. Forceps were used to remove two umbellets from opposite sides of the 3rd whorl of each umbel.
The umbellets were gently dipped 20 times each in bottles containing 10 mL of refrigerated, distilled water. The bottles were sealed and stored at 4 °C for subsequent laboratory analysis. For soluble sugar analysis, seven flowers per cultivar were sampled. Soluble fructose, glucose, and sucrose were identified using high performance liquid chromatography–mass spectrometry (HPLC–MS). The column was a Waters High Performance Carbohydrate Cartridge (Waters, Milford, ME, USA), 4.6 mm × 250 mm, fitted with a guard cartridge of the same material. The mobile phase was 75% methanol/25% water, isocratic at 1.2 mL per minute. Sugars were detected in negative mode by Atmospheric Pressure Chemical Ionisation (APCI) mass spectrometry on a Finnigan LCQ ion trap MS (Thermo Fisher Scientific, Waltham, MA, USA).
Sugars were detected as negative ion adducts formed by post column infusion of 20 µL/min of 5% formic acid in water. For the monosaccharides tandem MS was used, with the [M+ formate] anion at m/z = 225.3 being isolated, fragmented using 25% collision energy, and the subsequent daughter ion at m/z = 179 being further isolated and fragmented at 25% collision energy with the final products at m/z = 89, 119, 131, and 143 being used for quantitation. Sucrose quantitation was achieved by selected ion monitoring of the [M + formate] anion at m/z = 387.3.
Variation in total sugar concentration as well as the proportions of fructose, glucose, and sucrose between cultivars, were assessed using non-parametric Kruskal–Wallis tests with SPSS® 17.0.
4. Discussion
Insect-pollinated crops require reliable and effective insect visitation to maximise seed set and yield. In this study, significant differences in insect visitation rates were found among CMS carrot cultivars in field trials. In the initial trial involving 17 cultivars across eight root types and four CMS systems, a greater than threefold difference in overall visitation rate between the least and the most visited cultivars was found, confirming the results from previous trials that some cultivars of CMS carrots appear to be more attractive to pollinators, including honey bees, than others [
13,
14,
15,
31]. Visits of honey bees in our first trial varied among cultivars from 0.58 ± 0.23 to 3.46 ± 0.23 visits/umbel/5 min.
In the current study, more insects visited cultivars with the Berlicumer than the Nantes root types. While there are no reported comparable studies on the effect of carrot root type on pollinator attraction, we suggest that the observed difference between Berlicumer and Nantes demonstrates a genotypic influence on pollinator visitation. We found no significant difference in insect visitation between brown anther and petaloid phenotypes. In a previous study of three brown anther cultivars, one white petaloid cultivar, and one light green petaloid cultivar, brown anther cultivars were reported to be more attractive to honey bees than white petaloid types [
15]. Furthermore, Erickson
et al. found that honey bees show intra-phenotypic and genotypic foraging preferences between white and light green flowers in open-pollinated carrot cultivars [
14]. In our study, the lack of difference in visitation rate between CMS types indicates that differences in physical appearance associated with male sterility traits do not influence pollinator behaviour in this case. It is possible that root type and flower colour do not directly impact pollinator visitation, but instead, these traits could occasionally be genetically linked to other physiological traits, such as the production of nectar and volatiles, which would affect pollinator foraging behaviour and would explain the variation in results between studies.
Significant differences in floral diameters were evident among cultivars. However, when grouped into attractive and unattractive cultivars, although a significant difference in umbel diameter was observed, the unattractive cultivars were shown to produce the larger umbels. The influence of umbel diameter in this study varied according to the insect species, with more soldier beetles visiting larger umbels, and more ladybirds and native bees visiting smaller umbels. Other studies have also found that flower size affects the behaviour of pollinators. Abraham found that naïve bumblebees (
Bombus spp.) visited all of the flowers of 15 Althea (
Hibiscus syriacus L.) plants in equal numbers but visited the larger flowers first [
32]. It was suggested that this was an optimal foraging strategy, as larger flowers were likely to produce more nectar. Similarly, on
Achillea ptarmica (Asteraceae), it has been shown that the pollinating insects visited larger, many-headed inflorescences in preference to inflorescences with less floral heads [
33].
We found that honey bee visits decreased with increasing umbel diameter and/or if soldier beetles were initially present on the umbel. Such interference from other insects was also found by Danderson and Molano-Flores, who found that larger floral displays on
Eryngium yuccifolium (Apiaceae) attracted more visitors, but that herbivory on inflorescences negatively affected pollinator visitation [
34]. Furthermore, Kirk
et al. found that nectar-foraging honey bees were deterred from landing on umbels with real or simulated pollen beetles
Meligethes aeneus (Nitidulidae) present [
35]. It is plausible, given the negative relationship between the presence of
C. lugubris and
A. mellifera and the preference shown for smaller umbels by
A. mellifera, that it was the presence of
C. lugubris that deterred
A. mellifera from preferentially visiting larger umbels.
Although the structure of carrot inflorescences is in the form of large, highly visible, white or pale green umbels, which would provide a conspicuous visible cue for insects seeking nectar, there is no evidence that colour affects insect visitation. In addition, no significant levels of UV reflectance were observed; only small pinpricks of UV reflectance from nectar were seen in some of the carrot flowers.
In general, male sterile carrot flowers (not producing pollen) are the least visited [
3], warranting the analysis of nectar as the likely most important reward. However, our analysis indicated that nectar composition varies greatly between umbellets within cultivars but not significantly among cultivars. Furthermore, the nectar composition was dominated by monosaccharides, with only small amounts (2–12%) of sucrose. Such a mixture, rich in monosaccharides, can be predicted to be less than ideal for honey bee attraction, with species that are best pollinated by honey bees, generally found to be dominant in sucrose [
24,
25]. Our findings are not unexpected, as Apiaceae are known to have nectar with relatively low sucrose levels. We found that two of the least attractive cultivars assessed here contained both the highest and lowest total sugar concentrations (mean ± SEM: PN3: 70.9 ± 17.1 µg per umbellet; PBF1: 39.2 ± 12.9 µg per umbellet), indicating that sugar content within the nectars had little effect on pollinator visitation. In earlier studies, it has been found that
Carum carvi (annual caraway: Apiaceae), had just 13.6% sucrose in its nectar [
36], and
Eryngium campestre (field eryngo),
Scandix australis (southern shepherd’s needle),
Thapsia garganica, and
Tordylium apulum (Mediterranean hartwort) are also low in sucrose with an average of only 8.8% of the total sugar content [
37]. In the latter study, it was found that plants with high disaccharide/monosaccharide ratios were attractive to bees and wasps, whereas those with the opposite composition, similar to the carrot cultivars in our study, were more attractive to syrphids (hoverflies), anthomyids (root flies), and beetles.
The results from our investigation indicate that umbel colour and nectar sugar composition appear to be of little importance regarding pollinator attraction to hybrid CMS carrot umbels. Such a result strongly suggests that other factors, most likely of chemical origin, are at play. Floral volatiles have been proposed to be important for pollinator attraction in many systems [
38,
39], including hybrid carrot [
4,
7,
8]. Chemical components in nectar, apart from fructose, glucose, and sucrose, have also been highlighted as candidates for pollinator attractants [
22,
40]. For fertile lines, even pollen properties may be important in the decision making of pollinators [
41,
42]. In hybrid carrot pollination, no direct links among any variable chemical factors from the flower, nectar or pollen, whether volatiles, phenolics, sugars, or minerals, have been reported. We believe more detailed studies on the chemical ecology of hybrid carrot flowers are necessary to reveal which factors are driving the low pollinator attraction observed in hybrid carrot systems. Such an understanding would allow the development of practical breeding strategies to rectify the poor pollinator visitation and seed yields observed.






{kind=link}
{kind=link}
{kind=link}
{kind=link}







