Effects of Enzymatic Hydrolysis on the Functional Properties, Antioxidant Activity and Protein Structure of Black Soldier Fly (Hermetia illucens) Protein


 , ,
, ,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. One-Step Chemical Extraction of Protein
2.2. Enzymatic Hydrolysis of BSF Meal
2.3. Degree of Hydrolysis
2.4. Analysis of Functional Properties
2.5. Antioxidant Activity
2.6. Amino Acid Composition
2.7. Raman Spectroscopy
2.8. Data Analysis
3. Results and Discussion
3.1. Degree of Hydrolysis and Yield
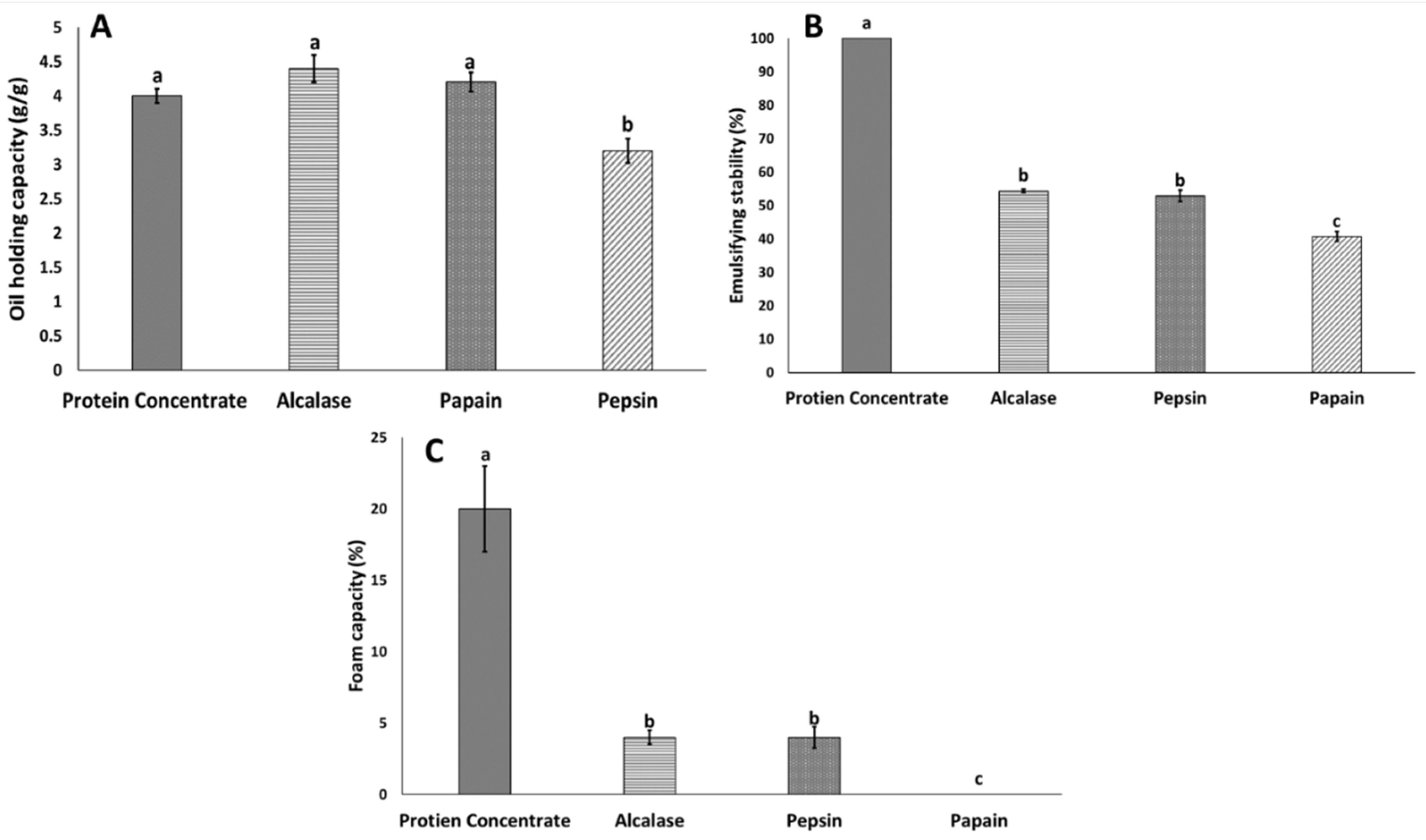
3.2. Functional Properties
3.3. Amino Acid Composition
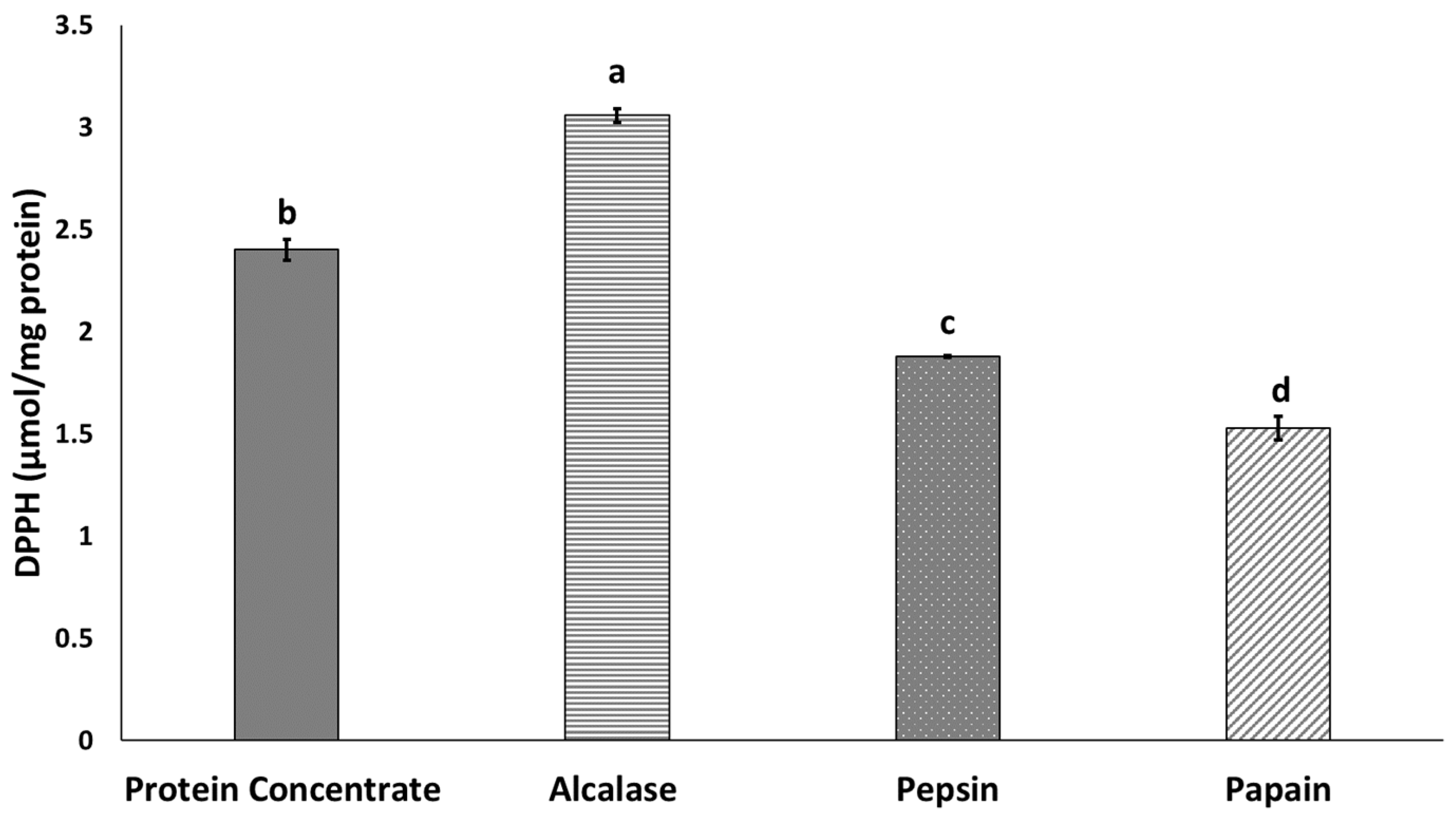
3.4. Antioxidant Activity
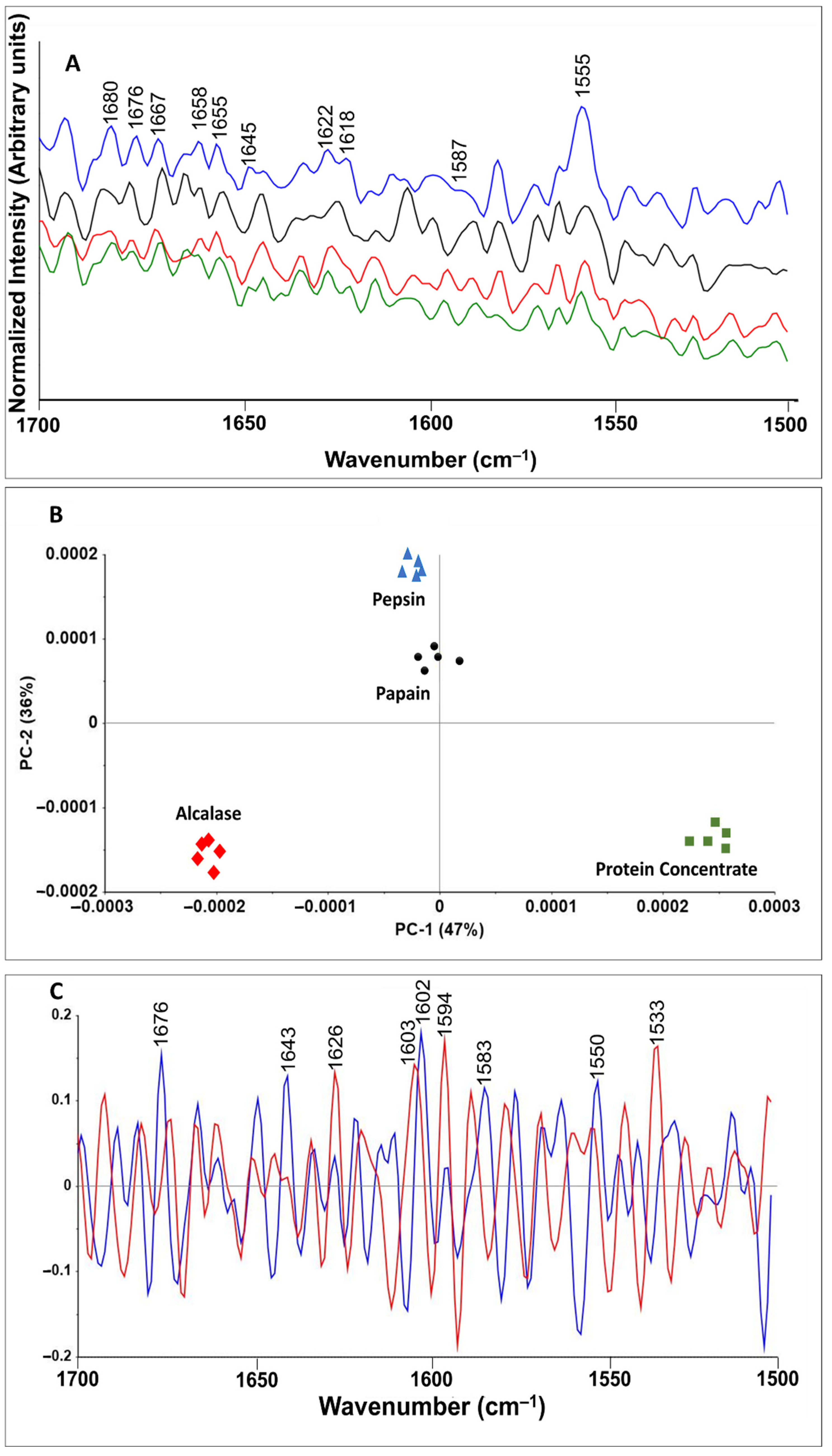
3.5. Raman Spectroscopy and Chemometrics
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2020; FAO: Rome, Italy, 2020; ISBN 978-92-5-132692-3. [Google Scholar]
- Herrero, M.; Wirsenius, S.; Henderson, B.; Rigolot, C.; Thornton, P.; Havlík, P.; de Boer, I.; Gerber, P.J. Livestock and the environment: What have we learned in the past decade? Annu. Rev. Environ. Resour. 2015, 40, 177–202. [Google Scholar] [CrossRef]
- Herrero, M.; Henderson, B.; Havlík, P.; Thornton, P.K.; Conant, R.T.; Smith, P.; Wirsenius, S.; Hristov, A.N.; Gerber, P.; Gill, M. Greenhouse gas mitigation potentials in the livestock sector. Nat. Clim. Chang. 2016, 6, 452–461. [Google Scholar] [CrossRef]
- Kanianska, R. Agriculture and its impact on land-use, environment, and ecosystem services. Landscape Ecology—The Influences of Land Use and Anthropogenic Impacts of Landscape Creation. Kanianska, R., Ed.; Slovakia. 2016, pp. 1–26. Available online: https://www.intechopen.com/books/landscape-ecology-the-influences-of-land-use-and-anthropogenic-impacts-of-landscape-creation/agriculture-and-its-impact-on-land-use-environment-and-ecosystem-services (accessed on 17 November 2020).
- Ziolkowska, J. Economic and environmental costs of agricultural food losses and waste in the US. ETP Int. J. Food Eng. 2017. [Google Scholar] [CrossRef]
- Liceaga, A.M. Approaches for utilizing insect protein for human consumption: Effect of enzymatic hydrolysis on protein quality and functionality. Ann. Entomol. Soc. Am. 2019, 112, 529–532. [Google Scholar] [CrossRef]
- Anaya, J.-M.; Shoenfeld, Y.; Rojas-Villarraga, A.; Levy, R.A.; Cervera, R. Autoimmunity: From Bench to Bedside; El Rosario University Press: Bogota, Colombia, 2013; ISBN 9587383664. [Google Scholar]
- Omotoso, O.T. Nutritional quality, functional properties and anti-nutrient compositions of the larva of Cirina forda (Westwood) (Lepidoptera: Saturniidae). J. Zhejiang Univ. Sci. B 2006, 7, 51–55. [Google Scholar] [CrossRef]
- Van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Borremans, A.; Bußler, S.; Sagu, S.T.; Rawel, H.; Schlüter, O.K.; Leen, V.C. Effect of Blanching Plus Fermentation on Selected Functional Properties of Mealworm (Tenebrio molitor) Powders. Foods 2020, 9, 917. [Google Scholar] [CrossRef]
- Ovissipour, M.; Rasco, B.; Shiroodi, S.G.; Modanlow, M.; Gholami, S.; Nemati, M. Antioxidant activity of protein hydrolysates from whole anchovy sprat (Clupeonella engrauliformis) prepared using endogenous enzymes and commercial proteases. J. Sci. Food Agric. 2013, 93, 1718–1726. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef]
- Purschke, B.; Meinlschmidt, P.; Horn, C.; Rieder, O.; Jäger, H. Improvement of techno-functional properties of edible insect protein from migratory locust by enzymatic hydrolysis. Eur. Food Res. Technol. 2018, 244, 999–1013. [Google Scholar] [CrossRef]
- Tang, Y.; Debnath, T.; Choi, E.-J.; Kim, Y.W.; Ryu, J.P.; Jang, S.; Chung, S.U.; Choi, Y.-J.; Kim, E.-K. Changes in the amino acid profiles and free radical scavenging activities of Tenebrio molitor larvae following enzymatic hydrolysis. PLoS ONE 2018, 13, e0196218. [Google Scholar] [CrossRef] [PubMed]
- Caligiani, A.; Marseglia, A.; Leni, G.; Baldassarre, S.; Maistrello, L.; Dossena, A.; Sforza, S. Composition of black soldier fly prepupae and systematic approaches for extraction and fractionation of proteins, lipids and chitin. Food Res. Int. 2018, 105, 812–820. [Google Scholar] [CrossRef]
- Firmansyah, M.; Abduh, M.Y. Production of protein hydrolysate containing antioxidant activity from Hermetia illucens. Heliyon 2019, 5, e02005. [Google Scholar] [CrossRef]
- Mintah, B.K.; He, R.; Dabbour, M.; Xiang, J.; Hui, J.; Agyekum, A.A.; Ma, H. Characterization of edible soldier fly protein and hydrolysate altered by multiple-frequency ultrasound: Structural, physical, and functional attributes. Process Biochem. 2020, 95, 157–165. [Google Scholar] [CrossRef]
- Zhu, D.; Huang, X.; Tu, F.; Wang, C.; Yang, F. Preparation, antioxidant activity evaluation, and identification of antioxidant peptide from black soldier fly (Hermetia illucens L.) larvae. J. Food Biochem. 2020, 44, e13186. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.H. Formol titration: An evaluation of its various modifications. Analyst 1957, 82, 488–498. [Google Scholar] [CrossRef]
- Sathivel, S.; Smiley, S.; Prinyawiwatkul, W.; Bechtel, P.J. Functional and nutritional properties of red salmon (Oncorhynchus nerka) enzymatic hydrolysates. J. Food Sci. 2005, 70, c401–c406. [Google Scholar] [CrossRef]
- Shahidi, F.; Han, X.-Q.; Synowiecki, J. Production and characteristics of protein hydrolysates from capelin (Mallotus villosus). Food Chem. 1995, 53, 285–293. [Google Scholar] [CrossRef]
- Yasumatsu, K.; Sawada, K.; Moritaka, S.; Misaki, M.; Toda, J.; Wada, T.; Ishii, K. Whipping and emulsifying properties of soybean products. Agric. Biol. Chem. 1972, 36, 719–727. [Google Scholar] [CrossRef]
- Pacheco-Aguilar, R.; Mazorra-Manzano, M.A.; Ramírez-Suárez, J.C. Functional properties of fish protein hydrolysates from Pacific whiting (Merluccius productus) muscle produced by a commercial protease. Food Chem. 2008, 109, 782–789. [Google Scholar] [CrossRef]
- Leni, G.; Soetemans, L.; Caligiani, A.; Sforza, S.; Bastiaens, L. Degree of Hydrolysis Affects the Techno-Functional Properties of Lesser Mealworm Protein Hydrolysates. Foods 2020, 9, 381. [Google Scholar] [CrossRef]
- Ovissipour, M.; Abedian, A.; Motamedzadegan, A.; Rasco, B.; Safari, R.; Shahiri, H. The effect of enzymatic hydrolysis time and temperature on the properties of protein hydrolysates from Persian sturgeon (Acipenser persicus) viscera. Food Chem. 2009, 115, 238–242. [Google Scholar] [CrossRef]
- Valencia, P.; Pinto, M.; Almonacid, S. Identification of the key mechanisms involved in the hydrolysis of fish protein by Alcalase. Process Biochem. 2014, 49, 258–264. [Google Scholar] [CrossRef]
- Kechaou, E.S.; Dumay, J.; Donnay-Moreno, C.; Jaouen, P.; Gouygou, J.-P.; Bergé, J.-P.; Amar, R. Ben Enzymatic hydrolysis of cuttlefish (Sepia officinalis) and sardine (Sardina pilchardus) viscera using commercial proteases: Effects on lipid distribution and amino acid composition. J. Biosci. Bioeng. 2009, 107, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Vioque, J.; Sánchez-Vioque, R.; Clemente, A.; Pedroche, J.; Millán, F. Partially hydrolyzed rapeseed protein isolates with improved functional properties. J. Am. Oil Chem. Soc. 2000, 77, 447–450. [Google Scholar] [CrossRef]
- Latorres, J.M.; Rios, D.G.; Saggiomo, G.; Wasielesky, W.; Prentice-Hernandez, C. Functional and antioxidant properties of protein hydrolysates obtained from white shrimp (Litopenaeus vannamei). J. Food Sci. Technol. 2018, 55, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Zayas, J.F. Emulsifying Properties of Proteins. In Functionality of Proteins in Food; Springer: Berlin/Heidelberg, Germany, 1997; pp. 134–227. ISBN 978-3-642-59116-7. [Google Scholar]
- Zielińska, E.; Karaś, M.; Baraniak, B. Comparison of functional properties of edible insects and protein preparations thereof. LWT 2018, 91, 168–174. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-protein conversion factors for three edible insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Dang, X.; Zheng, X.; Zhang, W. Housefly larvae hydrolysate: Orthogonal optimization of hydrolysis, antioxidant activity, amino acid composition and functional properties. BMC Res. Notes 2013, 6, 197. [Google Scholar] [CrossRef]
- Saadi, S.; Saari, N.; Anwar, F.; Hamid, A.A.; Ghazali, H.M. Recent advances in food biopeptides: Production, biological functionalities and therapeutic applications. Biotechnol. Adv. 2015, 33, 80–116. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-T.; Chen, S.Y.; Hsiao, S.C.; Wang, K.-T. Application of industrial protease “Alcalase” in peptide synthesis. Biomed. Biochim. Acta 1991, 50, S181–S186. [Google Scholar] [PubMed]
- Dong, S.; Zeng, M.; Wang, D.; Liu, Z.; Zhao, Y.; Yang, H. Antioxidant and biochemical properties of protein hydrolysates prepared from Silver carp (Hypophthalmichthys molitrix). Food Chem. 2008, 107, 1485–1493. [Google Scholar] [CrossRef]
- Ovissipour, M.; Rasco, B.; Tang, J.; Sablani, S. Kinetics of protein degradation and physical changes in thermally processed Atlantic salmon (Salmo salar). Food Bioprocess Technol. 2017, 10, 1865–1882. [Google Scholar] [CrossRef]
- Movasaghi, Z.; Rehman, S.; Rehman, I. Raman Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2007, 42, 493–541. [Google Scholar] [CrossRef]
- Hall, F.; Liceaga, A. Effect of microwave-assisted enzymatic hydrolysis of cricket (Gryllodes sigillatus) protein on ACE and DPP-IV inhibition and tropomyosin-IgG binding. J. Funct. Foods 2020, 64, 103634. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Enzyme | pH | Temperature (°C) | DH (%) | Yield (Wet Weight Basis%) | ||
|---|---|---|---|---|---|---|
| Hydrolysates | Oil | Solid Layer | ||||
| Alcalase 2.4 L | 6.85 | 60 | 18.4 ± 1.5 a | 51.4 ± 1.7 a | 7.1 ± 1.6 a | 41.6 ± 2.9 a |
| Papain | 6.85 | 60 | 15.34 ± 1.1 b | 37.8 ± 1.1 b | 4.6 ± 0.3 b | 57.5 ± 1.1 b |
| Pepsin | 3 | 37 | 9.8 ± 2.3 c | 44 ± 3.2 c | 3.2 ± 0.6 c | 52.8 ± 3.8 c |
| Amino Acid | Quantity (mg/g) | |||||
|---|---|---|---|---|---|---|
| Intact Protein | Protein Concentrate | Alcalase Hydrolysate | Pepsin Hydrolysates | Papain Hydrolysates | Reference Protein FAO/WHO 1985 | |
| ALA | 10.95 | 38.94 | 41.90 | 11.95 | 34.71 | |
| ARG | 6.34 | 31.81 | 28.86 | 9.46 | 22.86 | |
| ASP | 13.97 | 82.42 | 58.51 | 18.71 | 46.29 | |
| GLU | 16.75 | 95.48 | 74.86 | 35.19 | 68.31 | |
| GLY | 7.82 | 31.67 | 33.15 | 11.61 | 32.16 | |
| HIS | 5.09 | 19.35 | 20.71 | 10.39 | 18.69 | 15 |
| ILE | 6.91 | 35.97 | 25.44 | 5.17 | 16.94 | 30 |
| LEU | 9.86 | 50.63 | 37.43 | 5.54 | 24.97 | 59 |
| LYS | 8.64 | 54.08 | 36.12 | 7.52 | 27.04 | 45 |
| PHE | 6.06 | 34.43 | 20.21 | 3.78 | 12.31 | 38 |
| PRO | 9.15 | 31.11 | 38.47 | 14.51 | 34.30 | |
| SER | 3.84 | 13.74 | 14.41 | 4.11 | 11.99 | |
| THR | 4.42 | 16.63 | 18.14 | 4.45 | 14.14 | 23 |
| TYR | 7.10 | 27.62 | 30.51 | 7.01 | 23.85 | |
| VAL | 9.79 | 43.40 | 38.60 | 7.75 | 30.31 | 39 |
| HAA 1 | 59.83 | 262.11 | 232.56 | 55.70 | 177.37 | |
| PCAA | 20.06 | 105.23 | 85.68 | 27.37 | 68.59 | |
| NCAA | 30.72 | 177.90 | 133.37 | 53.91 | 114.60 | |
| TEAA | 50.78 | 254.49 | 196.65 | 44.60 | 144.39 | |
| EAAI | 0.3 | 1.1 | 0.85 | 0.27 | 0.7 | |
| AAA | 19.31 | 79.38 | 77.45 | 22.59 | 63.14 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batish, I.; Brits, D.; Valencia, P.; Miyai, C.; Rafeeq, S.; Xu, Y.; Galanopoulos, M.; Sismour, E.; Ovissipour, R. Effects of Enzymatic Hydrolysis on the Functional Properties, Antioxidant Activity and Protein Structure of Black Soldier Fly (Hermetia illucens) Protein. Insects 2020, 11, 876. https://doi.org/10.3390/insects11120876
Batish I, Brits D, Valencia P, Miyai C, Rafeeq S, Xu Y, Galanopoulos M, Sismour E, Ovissipour R. Effects of Enzymatic Hydrolysis on the Functional Properties, Antioxidant Activity and Protein Structure of Black Soldier Fly (Hermetia illucens) Protein. Insects. 2020; 11(12):876. https://doi.org/10.3390/insects11120876
Chicago/Turabian StyleBatish, Inayat, Devon Brits, Pedro Valencia, Caio Miyai, Shamil Rafeeq, Yixiang Xu, Magdalini Galanopoulos, Edward Sismour, and Reza Ovissipour. 2020. "Effects of Enzymatic Hydrolysis on the Functional Properties, Antioxidant Activity and Protein Structure of Black Soldier Fly (Hermetia illucens) Protein" Insects 11, no. 12: 876. https://doi.org/10.3390/insects11120876
APA StyleBatish, I., Brits, D., Valencia, P., Miyai, C., Rafeeq, S., Xu, Y., Galanopoulos, M., Sismour, E., & Ovissipour, R. (2020). Effects of Enzymatic Hydrolysis on the Functional Properties, Antioxidant Activity and Protein Structure of Black Soldier Fly (Hermetia illucens) Protein. Insects, 11(12), 876. https://doi.org/10.3390/insects11120876