Endophytic Beauveria bassiana in Foliar-Treated Citrus limon Plants Acting as a Growth Suppressor to Three Successive Generations of Diaphorina citri Kuwayama (Hemiptera: Liviidae)

 ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and methods
2.1. Source of Fungi
2.2. Source of Psyllids for Laboratory Assays
2.3. Citrus Plants
2.4. Preparation of Conidial Suspensions
2.5. Pathogenicity Test against D. citri Adults
2.6. Inoculation of Citrus Plants
2.7. Effects of Endophytic Entomopathogenic Fungi Inoculation on Plant Growth
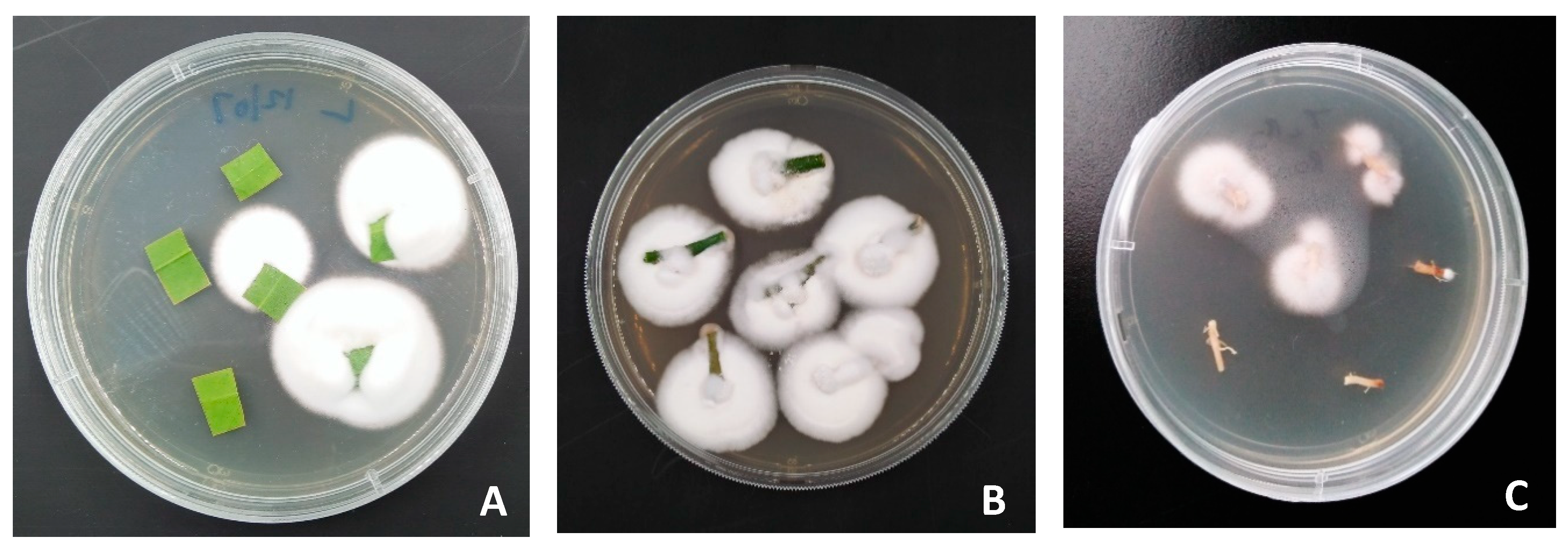
2.8. Assessment of Endophytic Colonization
2.9. Effects of Endophytically-Colonized Citrus Plants on the Development and Fecundity of Three Successive Generations of the Asian Citrus Psyllid
2.10. Statistical Analysis
3. Results
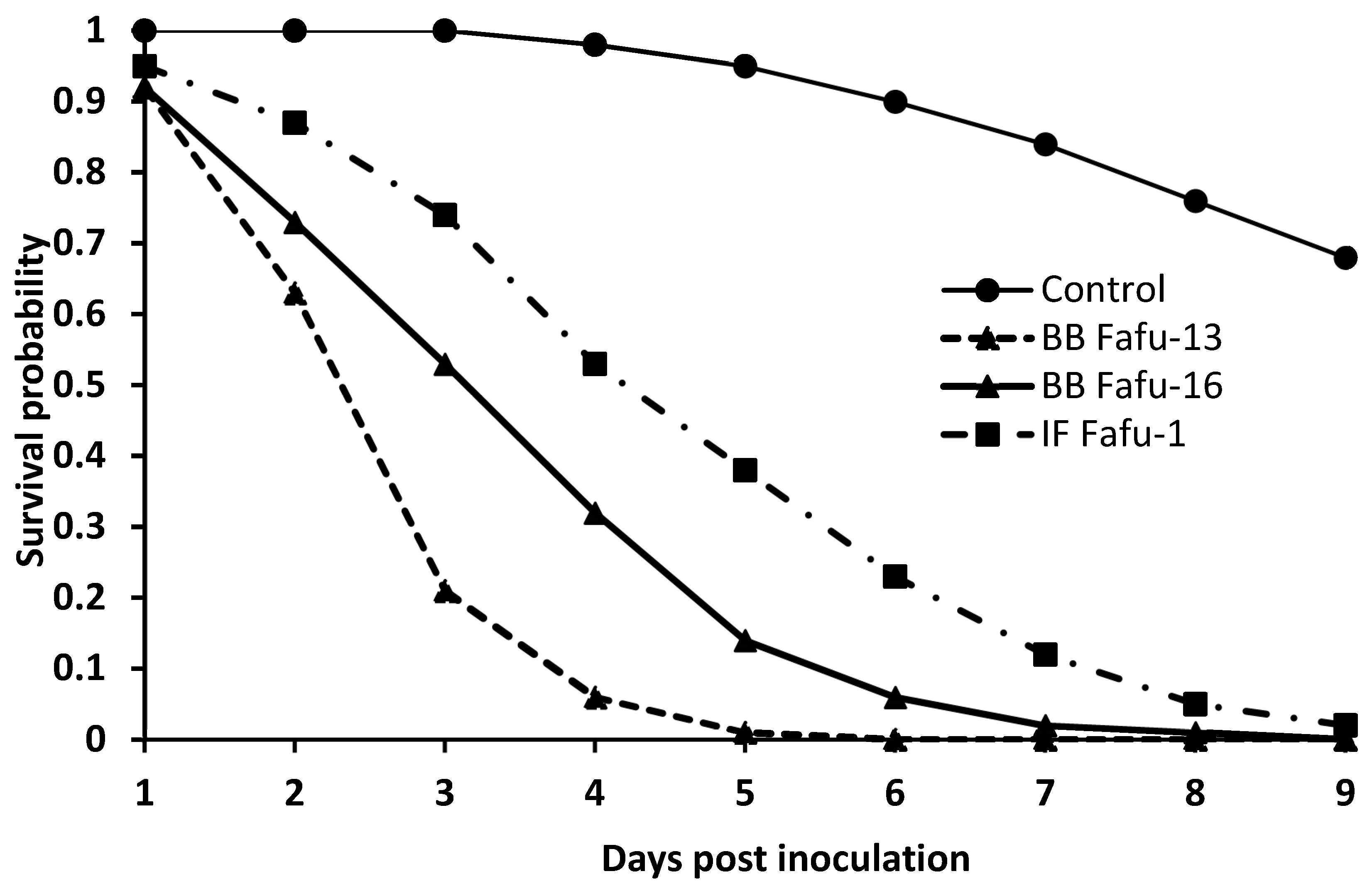
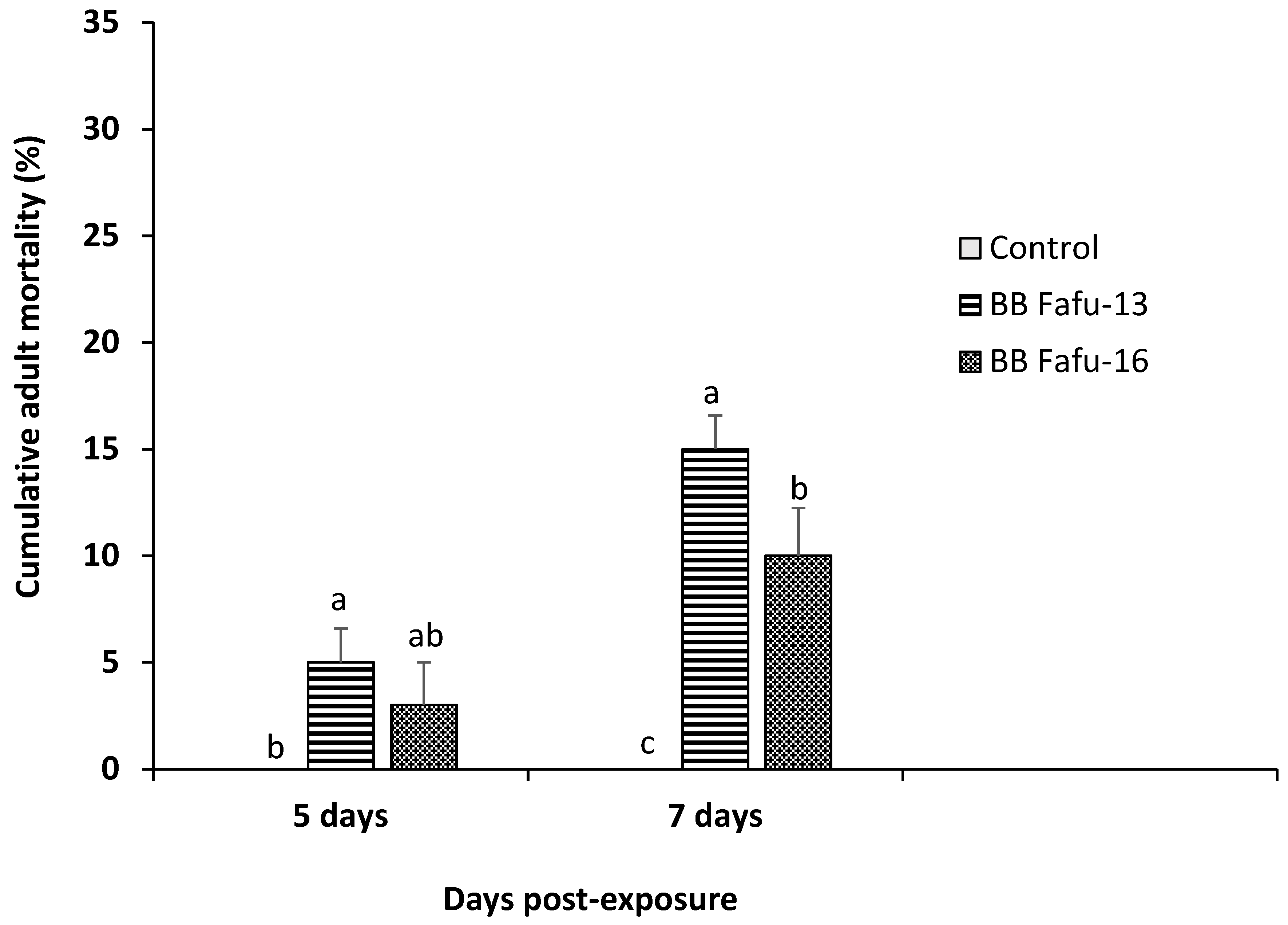
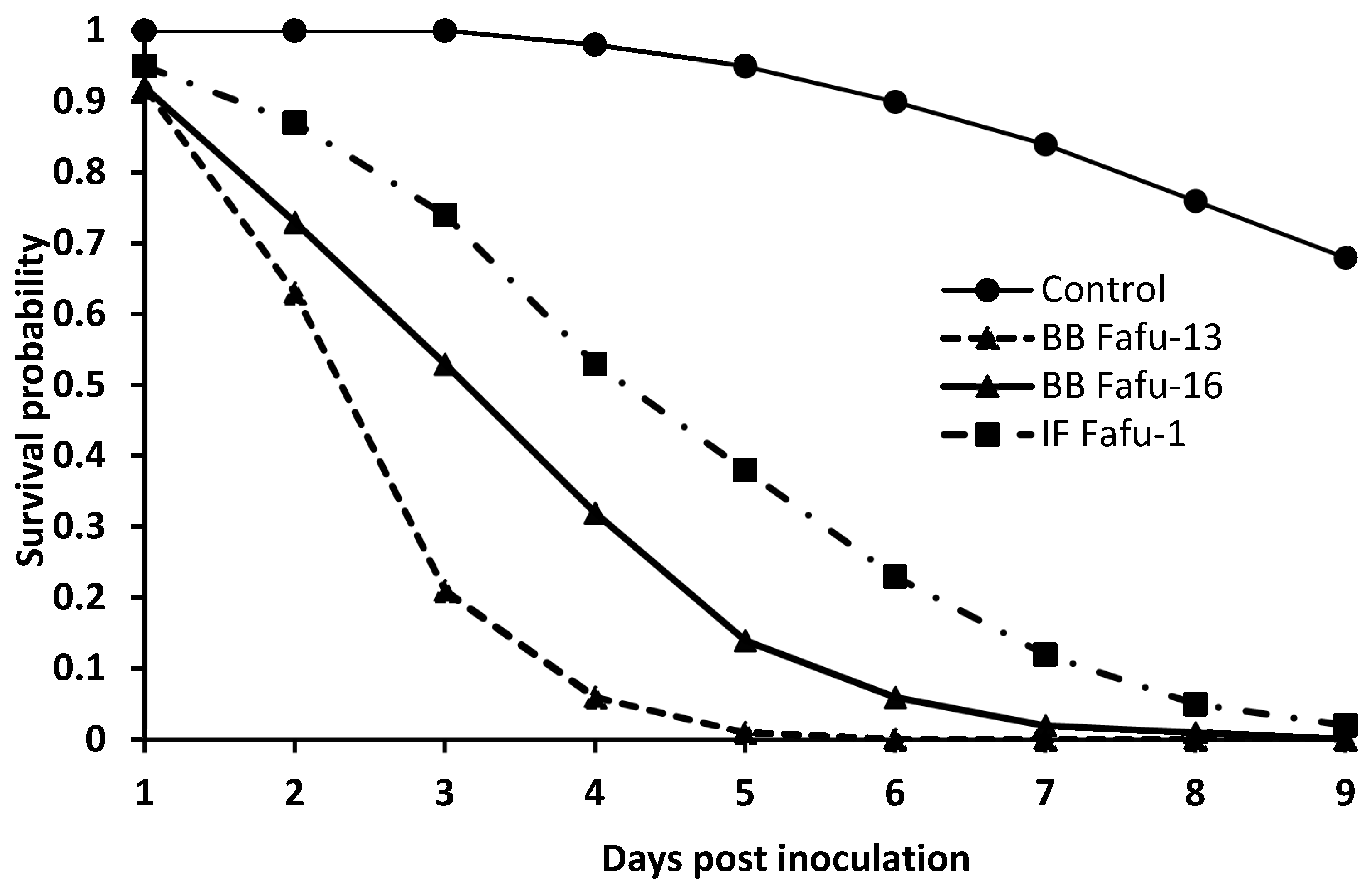
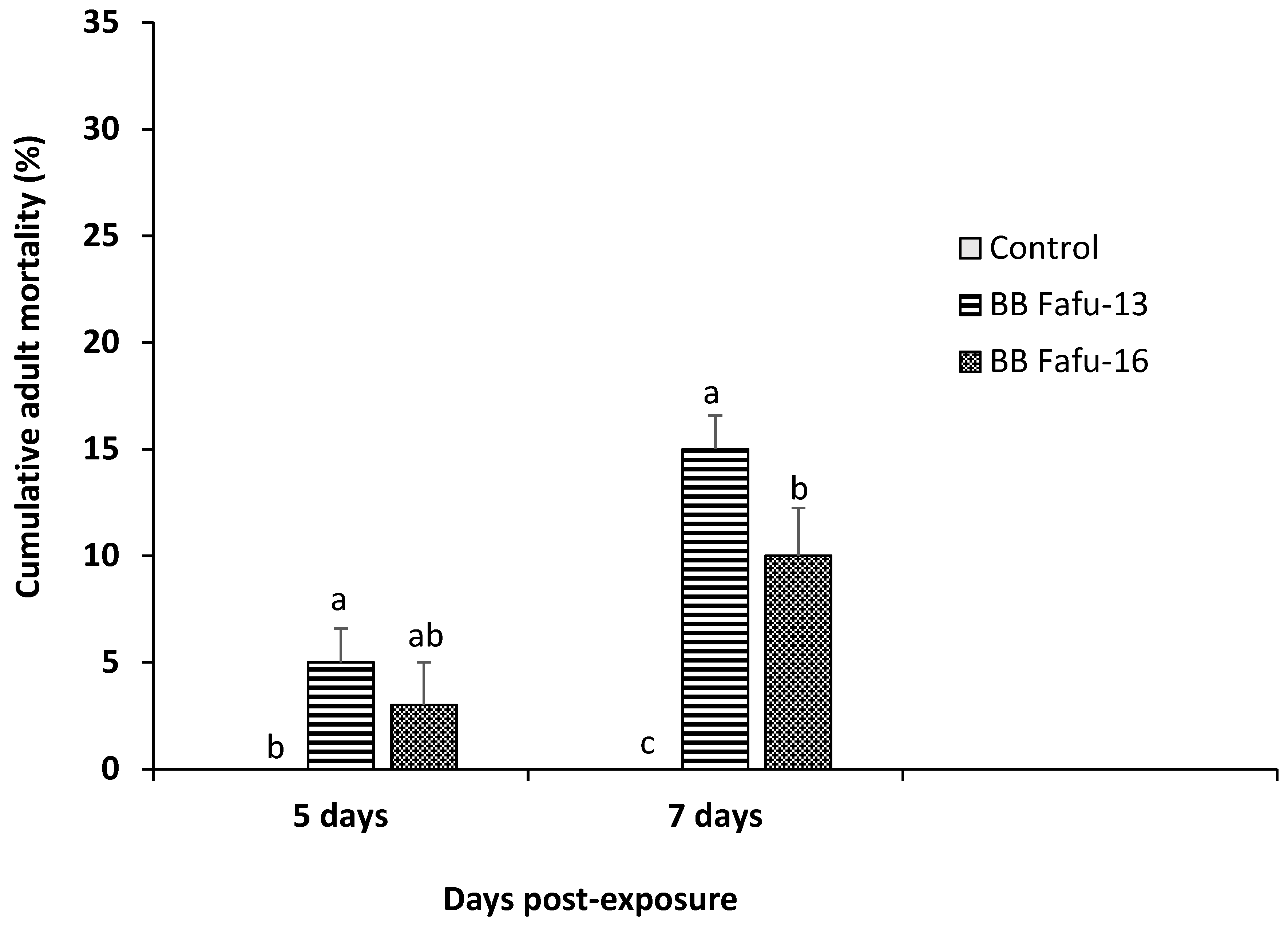
3.1. Pathogenicity of Entomopathogenic Fungal Strains against D. citri Adults
3.2. Assessment of Systemic Endophytic Colonization of Citrus Seedlings by Entomopathogenic Fungi
3.3. Effects of Endophytic Fungal Strains on Seedling Growth
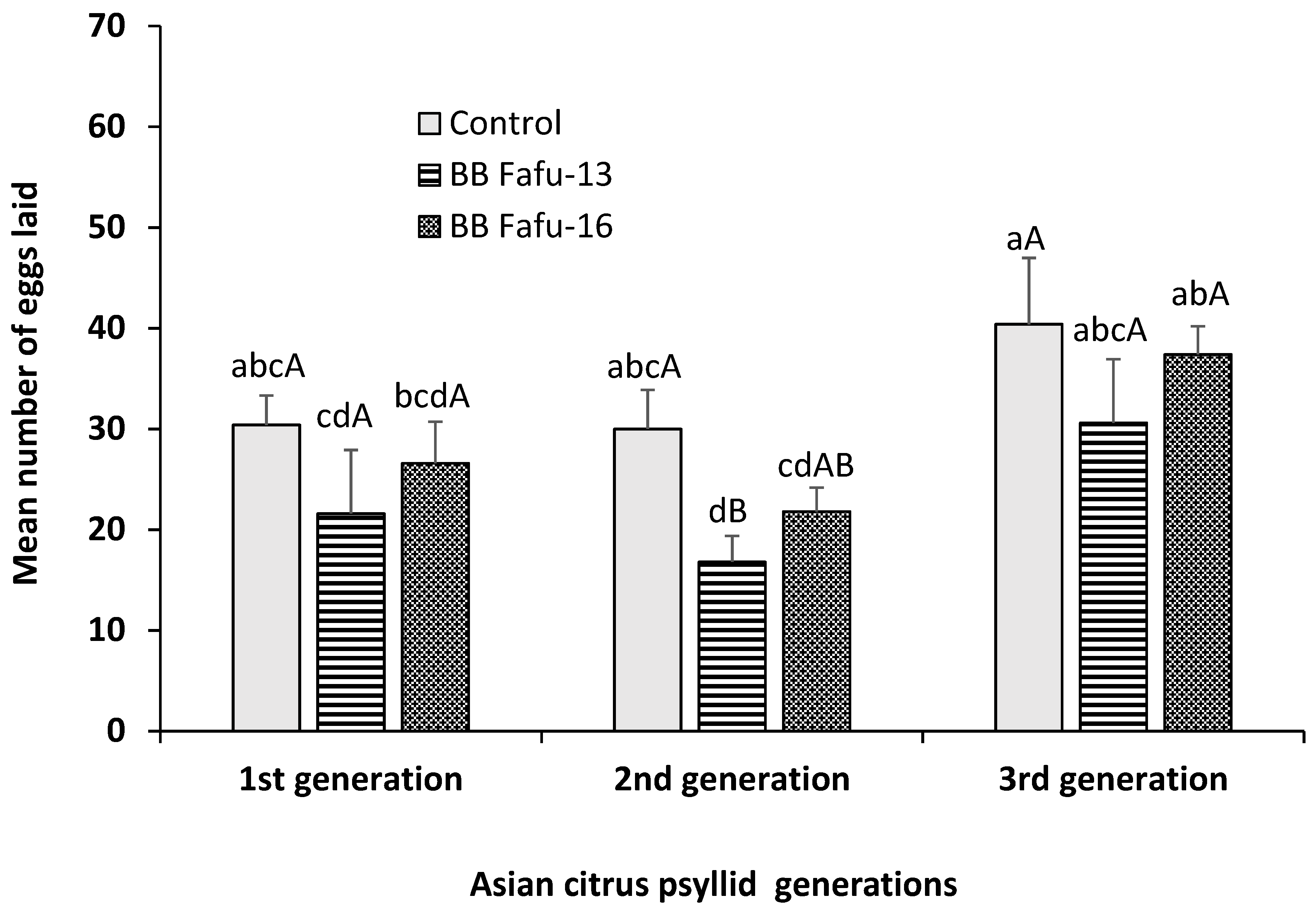
3.4. Assessment of the Effects of Endophytically-Colonized Citrus Plants on the Development and Fecundity of Three Successive Generations of the Asian Citrus Psyllid
4. Discussion
5. Conclusions and Future Research Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Donkersley, P.; Silva, F.W.S.; Carvalho, C.M.; Al-Sadi, A.M.; Elliot, S.L. Biological, environmental and socioeconomic threats to citrus lime production. J. Plant Dis. Prot. 2018, 125, 339–356. [Google Scholar] [CrossRef] [Green Version]
- Grafton-Cardwell, E.E.; Godfrey, K.E.; Rogers, M.E.; Childers, C.C.; Stansly, P.A. Asian Citrus Psyllid; U.S. Department of Agriculture: Washington, DC, USA, 2006.
- Gatineau, F.; Bonnot, F.; Yen, T.T.H.; Tuan, D.H.; Tuyen, N.D.; Truc, N.T.N. Effects of imidacloprid and fenobucarb on the dynamics of the psyllid Diaphorina citri Kuwayama and on the incidence of Candidatus Liberibacter asiaticus. Fruits 2010, 65, 209–220. [Google Scholar] [CrossRef]
- Kanga, L.H.B.; Eason, J.; Haseeb, M.; Qureshi, J.; Stansly, P. Monitoring for Insecticide Resistance in Asian Citrus Psyllid (Hemiptera: Psyllidae) Populations in Florida. J. Econ. Èntomol. 2015, 109, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Ruan, C.; Xia, Y.; Liu, B.; Chen, J.; Zhu, Y.; Fan, G.; Sequeira, R. Identification and entomopathogenicity of newly-isolated fungi infecting diaphorina citri kuwayama (homotera: Psyllidae) in murraya orchards of Fujian, China. J. Citrus Pathol. 2014, 1. Available online: https://escholarship.org/uc/item/5xf1z4s9 (accessed on 1 January 2014).
- Parra, J.; Gómez-Torres, M.; Paiva, P. Efeciência do parasitóide tamarixia radiata para o controle do psilídeo diaphorina citri em são paulo. Citric. Atual 2007, 61, 18–19. [Google Scholar]
- Lezama-Gutiérrez, R.; Molina-Ochoa, J.; Chávez-Flores, O.; Ángel-Sahagún, C.A.; Skoda, S.R.; Reyes-Martínez, G.; Barba-Reynoso, M.; Rebolledo-Domínguez, O.; Ruíz-Aguilar, G.M.; Foster, J.E. Use of the entomopathogenic fungi Metarhizium anisopliae, Cordyceps bassiana and Isaria fumosorosea to control Diaphorina citri (Hemiptera: Psyllidae) in Persian lime under field conditions. Int. J. Trop. Insect Sci. 2012, 32, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Hunter, W.B.; Avery, P.B.; Pick, D.; Powell, C.A. Broad Spectrum Potential of Isaria fumosorosea Against Insect Pests of Citrus. Fla. Èntomol. 2011, 94, 1051–1054. [Google Scholar] [CrossRef]
- Moran, P.J.; Patt, J.M.; Cabanillas, H.E.; Adamczyk, J.L.; Jackson, M.A.; Dunlap, C.A.; Hunter, W.B.; Avery, P.B. Localized autoinoculation and dissemination of isaria fumosorosea for control of the Asian citrus psyllid in South Texas. Subtrop. Plant Sci. 2011, 63, 23–35. [Google Scholar]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monzón, A.; Ownley, B.H.; et al. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Afolabi, O.G.; Hussain, M.; Qasim, M.; Wang, L. Prospects of endophytic fungal entomopathogens as biocontrol and plant growth promoting agents: An insight on how artificial inoculation methods affect endophytic colonization of host plants. Microbiol. Res. 2018, 217, 34–50. [Google Scholar] [CrossRef]
- Jaber, L.R.; Araj, S.-E. Interactions among endophytic fungal entomopathogens (Ascomycota: Hypocreales), the green peach aphid Myzus persicae Sulzer (Homoptera: Aphididae), and the aphid endoparasitoid Aphidius colemani Viereck (Hymenoptera: Braconidae). Biol. Control 2018, 116, 53–61. [Google Scholar] [CrossRef]
- Van Bael, S.; Mejía, L.; Bischoff, J.; Rojas, E.; Arnold, A.; Robbins, N.; Herre, E.; Kyllo, D.; Maynard, Z. Emerging Perspectives on the Ecological Roles of Endophytic Fungi in Tropical Plants. In The Fungal Community: Its Organization and Role in the Ecosystem; Dighton, J., White, J.F., Oudemans, P., Eds.; Taylor & Francis: London, UK, 2005; pp. 181–191. [Google Scholar]
- Jones, K.D. Aspects of the Biology and Biological Control of the European Corn Borer in North Carolina. Ph.D. Thesis, North Carolina State University, Raleigh, NC, USA, 1994. [Google Scholar]
- Evans, H.C.; Holmes, K.A.; Thomas, S.E. Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potential as biocontrol agents of cocoa diseases. Mycol. Prog. 2003, 2, 149–160. [Google Scholar] [CrossRef]
- Polishook, J.D.; Bills, G.F. Microfungi from Carpinus caroliniana. Can. J. Bot. 1991, 69, 1477–1482. [Google Scholar]
- Ganley, R.J.; Newcombe, G. Fungal endophytes in seeds and needles of Pinus monticola. Mycol. Res. 2006, 110, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Bing, L.A.; Lewis, L.C. Endophytic beauveria bassiana (balsamo) vuillemin in corn: The influence of the plant growth stage and ostrinia nubilalis (hübner). Biocontrol Sci. Technol. 1992, 2, 39–47. [Google Scholar] [CrossRef]
- Russo, M.L.; Pelizza, S.A.; Cabello, M.N.; Stenglein, S.A.; Scorsetti, A.C. Endophytic colonisation of tobacco, corn, wheat and soybeans by the fungal entomopathogen beauveria bassiana (ascomycota, hypocreales). Biocontrol Sci. Technol. 2015, 25, 475–480. [Google Scholar] [CrossRef]
- Reddy, G.V.; Tangtrakulwanich, K.; Wu, S.; Miller, J.H.; Ophus, V.L.; Prewett, J.; Jaronski, S.T. Evaluation of the effectiveness of entomopathogens for the management of wireworms (Coleoptera: Elateridae) on spring wheat. J. Invertebr. Pathol. 2014, 120, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Rodríguez, A.R.; Raya-Díaz, S.; Zamarreño, Á.M.; García-Mina, J.M.; del Campillo, M.C.; Quesada-Moraga, E. An endophytic beauveria bassiana strain increases spike production in bread and durum wheat plants and effectively controls cotton leafworm (spodoptera littoralis) larvae. Biol. Control 2018, 116, 90–102. [Google Scholar] [CrossRef]
- Posada, F.; Vega, F.E. Inoculation and colonization of coffee seedlings (Coffea arabica L.) with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycoscience 2006, 47, 284–289. [Google Scholar] [CrossRef]
- Posada, F.; Aime, M.C.; Peterson, S.W.; Rehner, S.A.; Vega, F.E. Inoculation of coffee plants with the fungal entomopathogen Beauveria bassiana (Ascomycota: Hypocreales). Mycol. Res. 2007, 111, 748–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akutse, K.; Maniania, N.; Fiaboe, K.; Berg, J.V.D.; Ekesi, S. Endophytic colonization of Vicia faba and Phaseolus vulgaris (Fabaceae) by fungal pathogens and their effects on the life-history parameters of Liriomyza huidobrensis (Diptera: Agromyzidae). Fungal Ecol. 2013, 6, 293–301. [Google Scholar] [CrossRef]
- Dash, C.K.; Bamisile, B.S.; Keppanan, R.; Qasim, M.; Lin, Y.; Islam, S.U.; Hussain, M.; Wang, L.; Ravindran, K. Endophytic entomopathogenic fungi enhance the growth of Phaseolus vulgaris L. (Fabaceae) and negatively affect the development and reproduction of Tetranychus urticae Koch (Acari: Tetranychidae). Microb. Pathog. 2018, 125, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Rondot, Y.; Reineke, A. Endophytic Beauveria bassiana in grapevine Vitis vinifera (L.) reduces infestation with piercing-sucking insects. Biol. Control 2018, 116, 82–89. [Google Scholar] [CrossRef]
- Clifton, E.H.; Jaronski, S.T.; Coates, B.S.; Hodgson, E.W.; Gassmann, A.J. Effects of endophytic entomopathogenic fungi on soybean aphid and identification of Metarhizium isolates from agricultural fields. PLoS ONE 2018, 13, e0194815. [Google Scholar] [CrossRef] [PubMed]
- Kwasna, H.; Szewczyk, W. Effects of fungi isolated from Quercus robur roots on growth of oak seedlings. Dendrobiology 2016, 75, 99–112. [Google Scholar] [CrossRef] [Green Version]
- Kwaśna, H.; Szewczyk, W.; Behnke-Borowczyk, J. Fungal root endophytes of q uercus robur subjected to flooding. For. Pathol. 2016, 46, 35–46. [Google Scholar] [CrossRef]
- Gathage, J.W.; Lagat, Z.O.; Fiaboe, K.K.M.; Akutse, K.S.; Ekesi, S.; Maniania, N.K. Prospects of fungal endophytes in the control of Liriomyza leafminer flies in common bean Phaseolus vulgaris under field conditions. BioControl 2016, 61, 741–753. [Google Scholar] [CrossRef]
- Klieber, J.; Reineke, A. The entomopathogen beauveria bassiana has epiphytic and endophytic activity against the tomato leaf miner tuta absoluta. J. Appl. Entomol. 2016, 140, 580–589. [Google Scholar] [CrossRef]
- Powell, W.A.; Klingeman, W.E.; Ownley, B.H.; Gwinn, K.D. Evidence of Endophytic Beauveria bassiana in Seed-treated Tomato Plants Acting as a Systemic Entomopathogen to Larval Helicoverpa zea (Lepidoptera: Noctuidae). J. Èntomol. Sci. 2009, 44, 391–396. [Google Scholar] [CrossRef]
- Parsa, S.; Ortiz, V.; Gómez-Jiménez, M.I.; Kramer, M.; Vega, F.E. Root environment is a key determinant of fungal entomopathogen endophytism following seed treatment in the common bean, Phaseolus vulgaris. Biol. Control 2018, 116, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Jaber, L.R.; Enkerli, J. Effect of seed treatment duration on growth and colonization of Vicia faba by endophytic Beauveria bassiana and Metarhizium brunneum. Biol. Control 2016, 103, 187–195. [Google Scholar] [CrossRef]
- Chandler, D. Basic and Applied Research on Entomopathogenic Fungi. In Microbial Control of Insect and Mite Pests; Elsevier: Amsterdam, The Netherlands, 2017; pp. 69–89. [Google Scholar]
- Krell, V.; Jakobs-Schoenwandt, D.; Vidal, S.; Patel, A.V. Encapsulation of Metarhizium brunneum enhances endophytism in tomato plants. Biol. Control 2018, 116, 62–73. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Wang, L. Fungal Endophytes: Beyond Herbivore Management. Front. Microbiol. 2018, 9, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humber, R.A. Fungi: Identification. In Manual of Techniques in Insect Pathology; Lacey, L.A., Ed.; Academic Press: London, UK, 1997. [Google Scholar]
- Humber, R.A. Identification of entomopathogenic fungi. In Manual of Techniques in Invertebrate Pathology, 2nd ed.; Lacey, L.A., Ed.; Academic Press: San Diego, CA, USA, 2012. [Google Scholar]
- Qasim, M.; Lin, Y.; Dash, C.K.; Bamisile, B.S.; Ravindran, K.; Islam, S.U.; Ali, H.; Wang, F.; Wang, L. Temperature-dependent development of Asian citrus psyllid on various hosts, and mortality by two strains of Isaria. Microb. Pathog. 2018, 119, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Parsa, S.; Ortiz, V.; Vega, F.E. Establishing Fungal Entomopathogens as Endophytes: Towards Endophytic Biological Control. J. Vis. Exp. 2013, e50360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, M.L.; Scorsetti, A.C.; Vianna, M.F.; Cabello, M.; Ferreri, N.; Pelizza, S. Endophytic effects of beauveria bassiana on corn (zea mays) and its herbivore, rachiplusia nu (lepidoptera: Noctuidae). Insects 2019, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- McKinnon, A.C.; Saari, S.; Moran-Diez, M.E.; Meyling, N.V.; Raad, M.; Glare, T.R. Beauveria bassiana as an endophyte: A critical review on associated methodology and biocontrol potential. BioControl 2017, 62, 1–7. [Google Scholar] [CrossRef]
- Schulz, B.; Guske, S.; Dammann, U.; Boyle, C. Endophyte-host interactions. Ii. Defining symbiosis of the endophyte-host interaction. Symbiosis 1998, 25, 213–227. [Google Scholar]
- Greenfield, M.; Gómez-Jiménez, M.I.; Ortiz, V.; Vega, F.E.; Kramer, M.; Parsa, S. Beauveria bassiana and Metarhizium anisopliae endophytically colonize cassava roots following soil drench inoculation. Biol. Control 2016, 95, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Barta, M. In planta bioassay on the effects of endophytic Beauveria strains against larvae of horse-chestnut leaf miner (Cameraria ohridella). Biol. Control 2018, 121, 88–98. [Google Scholar] [CrossRef]
- Petrini, O.; Fisher, P.J. Fungal endophytes in salicornia perennis. Trans. Br. Mycol. Soc. 1987, 87, 647–651. [Google Scholar] [CrossRef]
- Gandarilla-Pacheco, F.L.; López-Arroyo, J.I.; Galán-Wong, L.J.; Quintero-Zapata, I. Patogenicity of Native Entomopathogenic Fungi from the Mexican Citrus-Growing Area against Diaphorina citri Kuwayama (Hemiptera: Liviidae). Southwest. Èntomol. 2013, 38, 325–338. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, L.; Lin, J.; Chen, X.; Ji, J. Virulence of beauveria bassiana cqbb111 strain to diaphorina citri and neoseiulus cucumeris. Chin. J. Biol. Control 2013, 29, 56–60. [Google Scholar]
- Mantzoukas, S.; Chondrogiannis, C.; Grammatikopoulos, G. Effects of three endophytic entomopathogens on sweet sorghum and on the larvae of the stalk borer sesamia nonagrioides. Entomol. Exp. Appl. 2015, 154, 78–87. [Google Scholar] [CrossRef]
- Wagner, B.L.; Lewis, L.C. Colonization of Corn, Zea mays, by the Entomopathogenic Fungus Beauveria bassiana. Appl. Environ. Microbiol. 2000, 66, 3468–3473. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Zhou, J.-Y.; He, J.-X.; Du, W.; Bu, Y.-Q.; Liu, C.-H.; Dai, C.-C. Distribution of the Entomopathogenic Fungus Beauveria bassiana in Rice Ecosystems and Its Effect on Soil Enzymes. Curr. Microbiol. 2013, 67, 631–636. [Google Scholar] [CrossRef]
- Resquín-Romero, G.; Garrido-Jurado, I.; Delso, C.; Ríos-Moreno, A.; Quesada-Moraga, E. Transient endophytic colonizations of plants improve the outcome of foliar applications of mycoinsecticides against chewing insects. J. Invertebr. Pathol. 2016, 136, 23–31. [Google Scholar] [CrossRef]
- Vidal, S.; Jaber, L.R. Entomopathogenic fungi as endophytes: Plant-endophyte-herbivore interactions and prospects for use in biological control. Curr. Sci. 2015, 109, 46–54. [Google Scholar]
- Brownbridge, M.; Reay, S.D.; Nelson, T.L.; Glare, T.R. Persistence of Beauveria bassiana (Ascomycota: Hypocreales) as an endophyte following inoculation of radiata pine seed and seedlings. Biol. Control 2012, 61, 194–200. [Google Scholar] [CrossRef]
- Biswas, C.; Dey, P.; Satpathy, S.; Satya, P. Establishment of the fungal entomopathogen beauveria bassiana as a season long endophyte in jute (corchorus olitorius) and its rapid detection using scar marker. BioControl 2012, 57, 565–571. [Google Scholar] [CrossRef]
- Lopez, D.C.; Sword, G.A. The endophytic fungal entomopathogens Beauveria bassiana and Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum) and negatively affect survival of the cotton bollworm (Helicoverpa zea). Biol. Control 2015, 89, 53–60. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Fungal entomopathogens as endophytes: Can they promote plant growth? Biocontrol Sci. Technol. 2017, 27, 28–41. [Google Scholar] [CrossRef]
- Ríos-Moreno, A.; Garrido-Jurado, I.; Resquín-Romero, G.; Arroyo-Manzanares, N.; Arce, L.; Quesada-Moraga, E. Destruxin a production by metarhizium brunneum strains during transient endophytic colonisation of solanum tuberosum. Biocontrol Sci. Technol. 2016, 26, 1574–1585. [Google Scholar] [CrossRef]
- Ravindran, K.; Sivaramakrishnan, S.; Hussain, M.; Dash, C.K.; Bamisile, B.S.; Qasim, M.; Liande, W. Investigation and molecular docking studies of Bassianolide from Lecanicillium lecanii against Plutella xylostella (Lepidoptera: Plutellidae). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2018, 206, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and Management of Asian Citrus Psyllid, Vector of the Huanglongbing Pathogens. Annu. Rev. Èntomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Colonization Frequency in Different Plant Segments | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaf | Stem | Root | |||||||||
| UL | LL | US | LS | ||||||||
| n ** | + | % | + | % | + | % | + | % | + | % | |
| 4 wpi | |||||||||||
| BB Fafu-13 | 4 | 2 | 50 | 3 | 75 | 1 | 25 | 2 | 50 | 0 | 0 |
| BB Fafu-16 | 4 | 2 | 50 | 3 | 75 | 1 | 25 | 1 | 25 | 0 | 0 |
| 8 wpi | |||||||||||
| BB Fafu-13 | 4 | 1 | 25 | 3 | 75 | 0 | 0 | 2 | 50 | 0 | 0 |
| BB Fafu-16 | 4 | 1 | 25 | 2 | 50 | 0 | 0 | 1 | 25 | 0 | 0 |
| 12 wpi | |||||||||||
| BB Fafu-13 | 4 | 1 | 25 | 2 | 50 | 0 | 0 | 2 | 50 | 3 | 75 |
| BB Fafu-16 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Total | 24 | 7 | 29.2 | 13 | 54.2 | 2 | 8.3 | 8 | 33.3 | 3 | 12.5 |
| Treatments | Seedling Height (cm) | Leaf Number | ||||
|---|---|---|---|---|---|---|
| 4 wpi | 8 wpi | 12 wpi | 4 wpi | 8 wpi | 12 wpi | |
| BB Fafu-13 | 30 ± 1.5a | 38 ± 2.5a | 44 ± 2.9a | 29 ± 1.2a | 43 ± 1.4a | 54 ± 2.5a |
| BB Fafu-16 | 27 ± 0.9a | 37 ± 1.4ab | 41 ± 1.3ab | 25 ± 1.2ab | 41 ± 1.2ab | 52 ± 1.4a |
| IF Fafu-1 | 29 ± 1.0a | 34 ± 1.7ab | 38 ± 2.2bc | 24 ± 2.0b | 39 ± 1.7b | 48 ± 3.0ab |
| Control | 27 ± 0.4a | 32 ± 1.5b | 34 ± 1.5c | 23 ± 0.7b | 37 ± 1.4b | 44 ± 1.7b |
| Days Post Exposure | Treatments | Number of D. citri Nymphs and Adults | |||||
|---|---|---|---|---|---|---|---|
| 1st Generation Psyllids | 2nd Generation Psyllids | 3rd Generation Psyllids | |||||
| Nymphs | Adults | Nymphs | Adults | Nymphs | Adults | ||
| 7 days | Control | 5.2 ± 1.4a | 0.0 ± 0.0 | 4.4 ± 0.9a | 0.0 ± 0.0 | 5.2 ± 1.3a | 0.0 ± 0.0 |
| BB Fafu-13 | 1.0 ± 0.6b | 0.0 ± 0.0 | 1.0 ± 0.4b | 0.0 ± 0.0 | 1.6 ± 0.6b | 0.0 ± 0.0 | |
| BB Fafu-16 | 1.6 ± 0.5b | 0.0 ± 0.0 | 1.2 ± 0.5b | 0.0 ± 0.0 | 1.4 ± 0.4b | 0.0 ± 0.0 | |
| 10 days | Control | 13.2 ± 2.5a | 0.8 ± 0.5a | 12.2 ± 2.9a | 0.8 ± 0.5a | 12.0 ± 2.0a | 0.2 ± 0.2a |
| BB Fafu-13 | 1.2 ± 0.8b | 0.0 ± 0.0a | 1.2 ± 0.6b | 0.0 ± 0.0a | 5.4 ± 0.7b | 0.0 ± 0.0a | |
| BB Fafu-16 | 3.2 ± 0.7b | 0.0 ± 0.0a | 1.4 ± 0.6b | 0.0 ± 0.0a | 6.6 ± 0.6b | 0.0 ± 0.0a | |
| 14 days | Control | 24.0 ± 4.0a | 9.4 ± 2.0a | 16.4 ± 3.0a | 11.6 ± 2.1a | 19.4 ± 3.1a | 12.2 ± 0.7a |
| BB Fafu-13 | 2.4 ± 1.7b | 0.0 ± 0.0b | 2.4 ± 1.5b | 0.0 ± 0.0b | 3.4 ± 0.2b | 1.6 ± 0.5b | |
| BB Fafu-16 | 5.4 ± 1.3b | 2.0 ± 0.7b | 3.2 ± 1.0b | 0.8 ± 0.6b | 7.4 ± 0.9b | 2.4 ± 0.2b | |
| 17 days | Control | 26.0 ± 6.4a | 16.6 ± 6.3a | 19.4 ± 2.5a | 22.6 ± 5.4a | 19.2 ± 2.3a | 17.4 ± 2.6a |
| BB Fafu-13 | 2.6 ± 1.7b | 0.0 ± 0.0b | 2.4 ± 1.5b | 0.0 ± 0.0b | 5.6 ± 0.6c | 3.4 ± 0.5b | |
| BB Fafu-16 | 5.4 ± 0.9b | 3.6 ± 1.1b | 3.0 ± 1.0b | 1.6 ± 0.5b | 11.0 ± 1.6b | 4.6 ± 1.0b | |
| 21 days | Control | 27.6 ± 8.4a | 23.8 ± 6.8a | 27.0 ± 4.3a | 24.2 ± 3.9a | 24.4 ± 2.8a | 29.8 ± 4.1a |
| BB Fafu-13 | 1.8 ± 1.2b | 1.2 ± 1.0b | 0.8 ± 0.6b | 1.2 ± 0.8b | 6.0 ± 0.8b | 4.4 ± 0.5b | |
| BB Fafu-16 | 8.8 ± 1.0b | 6.0 ± 0.5b | 3.6 ± 2.0b | 2.4 ± 0.8b | 10.2 ± 1.5b | 6.6 ± 1.1b | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Qasim, M.; Ramos Aguila, L.C.; Wang, F.; Keppanan, R.; Wang, L. Endophytic Beauveria bassiana in Foliar-Treated Citrus limon Plants Acting as a Growth Suppressor to Three Successive Generations of Diaphorina citri Kuwayama (Hemiptera: Liviidae). Insects 2019, 10, 176. https://doi.org/10.3390/insects10060176
Bamisile BS, Dash CK, Akutse KS, Qasim M, Ramos Aguila LC, Wang F, Keppanan R, Wang L. Endophytic Beauveria bassiana in Foliar-Treated Citrus limon Plants Acting as a Growth Suppressor to Three Successive Generations of Diaphorina citri Kuwayama (Hemiptera: Liviidae). Insects. 2019; 10(6):176. https://doi.org/10.3390/insects10060176
Chicago/Turabian StyleBamisile, Bamisope Steve, Chandra Kanta Dash, Komivi Senyo Akutse, Muhammad Qasim, Luis Carlos Ramos Aguila, Fangfei Wang, Ravindran Keppanan, and Liande Wang. 2019. "Endophytic Beauveria bassiana in Foliar-Treated Citrus limon Plants Acting as a Growth Suppressor to Three Successive Generations of Diaphorina citri Kuwayama (Hemiptera: Liviidae)" Insects 10, no. 6: 176. https://doi.org/10.3390/insects10060176
APA StyleBamisile, B. S., Dash, C. K., Akutse, K. S., Qasim, M., Ramos Aguila, L. C., Wang, F., Keppanan, R., & Wang, L. (2019). Endophytic Beauveria bassiana in Foliar-Treated Citrus limon Plants Acting as a Growth Suppressor to Three Successive Generations of Diaphorina citri Kuwayama (Hemiptera: Liviidae). Insects, 10(6), 176. https://doi.org/10.3390/insects10060176