Halyomorpha halys (Hemiptera: Pentatomidae) Genetic Diversity in North America and Europe

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Material
2.2. DNA Isolation, Amplification, and Sequencing
2.3. Phylogenetic Analysis
3. Results
3.1. Analysis of Cytochrome Oxidase I Gene
3.2. Analysis of Internal Transcribed Spacer 1
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Josifov, M.V.; Kerzhner, I.M. Heteroptera aus Korea: II. Teil (Aradidae, Berytidae, Lygaeidae, Pyrrhocoridae, Rhopalidae, Alydidae, Coreidae, Urostylidae, Acanthosomatidae, Scutelleridae, Pentatomidae, Cydnidae, Plataspidae. Fragm. Faun. 1978, 23, 137–196. [Google Scholar] [CrossRef]
- Rider, D.A.; Zheng, L.Y.; Kerzhner, I.M. Checklist and nomenclatural notes on the Chinese Pentatomidae (Heteroptera). II. Pentatominae. Zoosyst. Ross. 2002, 11, 135–153. [Google Scholar]
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C.; Adachi, I.; Uchino, K.; Mochizuki, F.; Arakawa, R.; Namura, Y.; et al. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Hoebeke, E.R.; Carter, M.E. Halyomorpha halys (Stal) (Heteroptera: Pentatomidae): A polyphagous plant pest from Asia newly detected in North America. Proc. Entomol. Soc. Wash. 2003, 105, 225–237. [Google Scholar]
- Fogain, R.; Graff, S. First records of the invasive pest, Halyomorpha halys (Hemiptera: Pentatomidae), in Ontario and Quebec. J. Entomol. Soc. Ont. 2011, 142, 45–48. [Google Scholar]
- Cesari, M.; Maistrello, L.; Ganzerli, F.; Dioli, P.; Rebecchi, L.; Guidetti, R. A pest alien invasion in progress: Potential pathways of origin of the brown marmorated stink bug Halyomorpha halys populations in Italy. J. Pest Sci. 2015, 88, 1–7. [Google Scholar] [CrossRef]
- Milonas, P.G.; Partsinevelos, G.K. First report of brown marmorated stink bug Halyomorpha halys Stål (Hemiptera: Pentatomidae) in Greece. EPPO Bull. 2014, 44, 183–186. [Google Scholar] [CrossRef]
- Seat, J. Halyomorpha halys (Stal, 1855) (Heteroptera: Pentatomidae) a new invasive species in Serbia. Acta Entomol. Serb. 2015, 20, 167–171. [Google Scholar]
- Dioli, P.; Leo, P.; Maistrello, L. Prime segnalazioni in Spagna e in Sardegna della specie aliena Halyomorpha halys (Stal, 1855) e note sulla sua distribuzione in Europa (Hemiptera, Pentatomidae). Rev. Gaditana Entomol. 2016, 7, 539–548. [Google Scholar]
- Simov, N. The invasive brown marmorated stink bug Halyomorpha halys (Stal 1855 (Heteroptera: Pentatomidae) already in Bulgaria. Ecol. Montenegrina 2016, 9, 51–53. [Google Scholar]
- Maistrello, L.; Dioli, P.; Bariselli, M.; Mazzoli, G.L.; Giacalone-Forini, I. Citizen science and early detection of invasive species: Phenology of first occurrences of Halyomorpha halys in Southern Europe. Biol. Invasions 2016, 18, 3109–3116. [Google Scholar] [CrossRef]
- Wermelinger, B.; Wyniger, D.; Forster, B. First records of an invasive bug in Europe: Halyomorpha halys Stål (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Bull. Soc. Entomol. Suisse 2008, 81, 1–8. [Google Scholar]
- Vétek, G.; Papp, V.; Haltrich, A.; Rédei, D. First record of the brown marmorated stink bug, Halyomorpha halys (Hemiptera: Heteroptera: Pentatomidae), in Hungary, with description of the genitalia of both sexes. Zootaxa 2014, 3780, 194–200. [Google Scholar] [CrossRef]
- Gariepy, T.D.; Haye, T.; Fraser, H.; Zhang, J. Occurrence, genetic diversity, and potential pathways of entry of Halyomorpha halys in newly invaded areas of Canada and Switzerland. J. Pest Sci. 2014, 87, 17–28. [Google Scholar] [CrossRef]
- Arnold, K. Halyomorpha halys (Stal, 1855), Eine fur die europaische Fauna neu nachgewiesene Wanzenart (Insecta: Heteroptera: Pentatomidae: Cappaeini). Mitteillungen Thuring. Entomol. 2009, 16, 19. [Google Scholar]
- Heckmann, R. Erster nachweis von Halyomorpha halys (Stål, 1855)(Heteroptera: Pentatomidae) für deutschland. Heteropteron 2012, 36, 17–18. [Google Scholar]
- Callot, H.; Brua, C. Halyomorpha halys (Stal, 1855), la Punaise diabolique, nouvelle espece pour la faune de France (Heteroptera Pentatomidae). L’Entomologiste 2013, 69, 69–71. [Google Scholar]
- Hemala, V.; Kment, P. First record of Halyomorpha halys and mass occurrence of Nezara viridula in Slovakia. Plant Prot. Sci. 2017, 53, 1–7. [Google Scholar]
- Macavei, L.; Baetan, R.; Oltean, I.; Florean, T.; Varga, M.; Costi, E.; Maistrello, L. First detection of Halyomorpha halys, a new invasive species with a high potential of damage on agricultural crops in Romania. Lucr. Stiint. Ser. Agron. 2015, 58, 105–108. [Google Scholar]
- Gapon, D.A. First records of the brown marmorated stink bug, Halyomorpha halys (Stal, 1855) (Heteroptera: Pentatomidae) in Russia, Abkhazia, and Georgia. Entomol. Rev. 2016, 96, 1086–1088. [Google Scholar] [CrossRef]
- Faundez, E.I.; Rider, D.A. The brown marmorated stink bug Halyomorpha halys (Stal, 1855) (Heteroptera: Pentatomidae) in Chile. Arq. Entomol. 2017, 17, 305–307. [Google Scholar]
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Lo, P.L.; Walker, J.T.S.; Rogers, D.J. Risks to pest management in New Zealand’s pipfruit Integrated Fruit Production programme. N. Z. Plant Prot. 2015, 68, 306–312. [Google Scholar]
- Duthie, C.; Michael, T.; Stephenson, B.; Yamoah, E.; McDonald, B. Risk Analysis of Halyomorpha halys (Brown Marmorated Stink Bug) on All Pathways; Biosecurity Risk Analysis Group, Ministry of Agriculture and Forestry of New Zealand: Wellington, New Zealand, 2012.
- Leskey, T.C.; Nielsen, A.L. Impact of the Invasive Brown Marmorated Stink Bug in North America and Europe: History, Biology, Ecology, and Management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [PubMed]
- Gariepy, T.D.; Bruin, A.; Haye, T.; Milonas, P.; Vétek, G. Occurrence and genetic diversity of new populations of Halyomorpha halys in Europe. J. Pest Sci. 2015, 88, 451–460. [Google Scholar] [CrossRef]
- Cesari, M.; Maistrello, L.; Piemontese, L.; Bonini, R.; Dioli, P.; Lee, W.; Park, C.-G.; Partsinevels, G.K.; Rebecchi, L.; Guidetti, R. Genetic diversity of the brown marmorated stink bug Halyomorpha halys in the invaded territories of Europe and its patterns of diffusion in Italy. Biol. Invasions 2017, 20, 1073–1092. [Google Scholar] [CrossRef]
- Xu, J.; Fonseca, D.M.; Hamilton, G.C.; Hoelmer, K.A.; Nielsen, A.L. Tracing the origin of US brown marmorated stink bugs, Halyomorpha halys. Biol. Invasions 2014, 16, 153–166. [Google Scholar] [CrossRef]
- Zhu, G.-P.; Ye, Z.; Du, J.; Zhang, D.-L.; Zhen, Y.; Zheng, C.; Zhao, L.; Li, M.; Bu, W.-J. Range wide molecular data and niche modeling revealed the Pleistocene history of a global invader (Halyomorpha halys). Sci. Rep. 2016, 6, 23192. [Google Scholar] [CrossRef]
- Kurata, A.; Fujiwara, A.; Haruyama, N.; Tsuchida, T. Multiplex PCR method for rapid identification of genetic group and symbiont infection status in Bemisia tabaci (Hemiptera: Aleyrodidae). Appl. Entomol. Zool. 2016, 51, 167–172. [Google Scholar] [CrossRef]
- Kumar, L.S.; Shankar, P.; Kulkarni, V.M. Analyses of the internal transcribed rDNA spacers (ITS1 and ITS2) of Indian weevils of Odoiporus longicollis (Olivier) reveal gene flow between locations. Int. J. Trop. Insect Sci. 2018, 38, 313–329. [Google Scholar] [CrossRef]
- Allio, R.; Donega, S.; Galtier, N.; Nabholz, B. Large variation in the ratio of mitochondrial to nuclear mutation rate across animals: Implications for genetic diversity and the use of mitochondrial DNA as a molecular marker. Mol. Biol. Evol. 2017, 34, 2762–2772. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Penton, E.H.; Burns, J.M.; Janzen, D.H.; Hallwachs, W. Herbertetal_2004. Proc. Natl. Acad. Sci. USA 2004, 101, 14812–14817. [Google Scholar] [CrossRef] [PubMed]
- Valentin, R.E.; Maslo, B.; Lockwood, J.L.; Pote, J.; Fonseca, D.M. Real-time PCR assay to detect brown marmorated stink bug, Halyomorpha halys (Stål), in environmental DNA. Pest Manag. Sci. 2016, 72, 1854–1861. [Google Scholar] [CrossRef] [PubMed]
- Morrison, W.R.; Milonas, P.; Kapantaidaki, D.E.; Cesari, M.; Di Bella, E.; Guidetti, R.; Haye, T.; Maistrello, L.; Moraglio, S.T.; Piemontese, L.; et al. Attraction of Halyomorpha halys (Hemiptera: Pentatomidae) haplotypes in North America and Europe to baited traps. Sci. Rep. 2017, 7, 16941. [Google Scholar] [CrossRef] [PubMed]
- Milligan BG Total DNA Isolation. Molecular Genetic Analysis of Population: A Practical Approach; Hoelzel, A.R., Ed.; Oxford University Press: Oxford, UK; New York, NY, USA; Tokyo, Japan, 1998; pp. 29–64. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Von der Schulenburg, J.H.G.; Hancock, J.M.; Pagnamenta, A.; Sloggett, J.J.; Majerus, M.E.N.; Hurst, G.D.D. Extreme Length and Length Variation in the First Ribosomal Internal Transcribed Spacer of Ladybird Beetles (Coleoptera: Coccinellidae). Mol. Biol. Evol. 2001, 18, 648–660. [Google Scholar] [CrossRef] [PubMed]
- Remigio, E.A.; Blair, D. Relationships among problematic North American stagnicoline snails (Pulmonata: Lymnaeidae) reinvestigated using nuclear ribosomal DNA internal transcribed spacer sequences. Can. J. Zool. 2008, 75, 1540–1545. [Google Scholar] [CrossRef]
- Geneious | Bioinformatics Software for Molecular Sequence Data Analysis. Available online: https://www.geneious.com/ (accessed on 4 April 2019).
- Lee, W.; Kang, J.; Jung, C.; Hoelmer, K.; Lee, S.H.; Lee, S. Complete mitochondrial genome of brown marmorated stink bug Halyomorpha halys (Hemiptera: Pentatomidae), and phylogenetic relationships of hemipteran suborders. Mol. Cells 2009, 28, 155–165. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 36, 1547–1549. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Valentin, R.E.; Nielsen, A.L.; Wiman, N.G.; Lee, D.-H.; Fonseca, D.M. Global invasion network of the brown marmorated stink bug, Halyomorpha halys. Sci. Rep. 2017, 7, 9866. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Guidetti, R.; Cesari, M.; Gariepy, T.D.; Park, Y.-L.; Park, C.-G. Genetic Diversity of Halyomorpha halys (Hemiptera, Pentatomidae) in Korea and Comparison with COI Sequence Datasets from East Asia, Europe, and North America. Florida Entomol. 2018, 101, 49–54. [Google Scholar] [CrossRef]
- Kerdelhué, C.; Boivin, T.; Burban, C. Contrasted invasion processes imprint the genetic structure of an invasive scale insect across southern Europe. Heredity 2014, 113, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Murata, K.; Matsuura, A. Rapid evolution of an introduced insect Ophraella communa LeSage in new environments: Temporal changes and geographical differences in photoperiodic response. Entomol. Sci. 2015, 18, 104–112. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
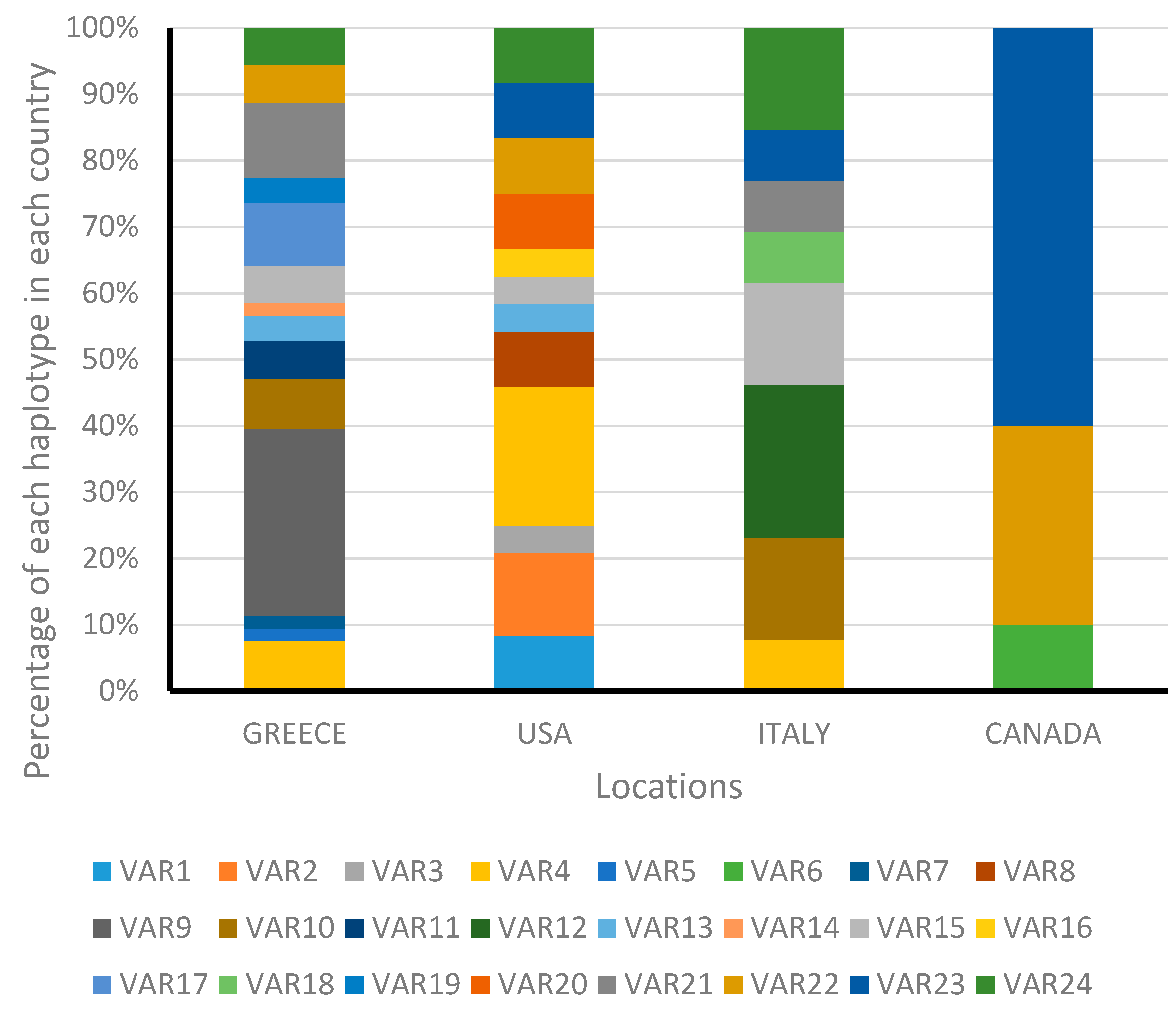{kind=link}
{kind=link}
| Country | Region | COI | ITS1 | ||
|---|---|---|---|---|---|
| Morrison et al. 2017 [35] | This Study | Morrison et al. 2017 [35] | This Study | ||
| Greece | Athens | 195 | 50 | ||
| Peloponnese | 2 | 2 | |||
| Chania | 1 | 1 | |||
| Italy | Modena | 20 | 11 | ||
| Perugia | 3 | 2 | |||
| USA | Smithsburg, Maryland | 60 | 19 | ||
| North Carolina | 20 | 5 | |||
| Canada | London, Ontario | 13 | 10 | ||
| Region | N | n | h | H | Hd ± SD | Pi ± SD | S | k | SVS | PIS | Tajima’s D | p-Value | Fu’s Fs |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Canada | 10 | 10 | 3 | 1 | 0.600 ± 0.131 | 0.011± 0.003 | 12 | 4.800 | 3 | 9 | 0.594 | p > 0.10 | 4.717 |
| Greece | 53 | 53 | 14 | 7 | 0.830 ± 0.034 | 0.022 ± 0.001 | 82 | 7.896 | 2 | 80 | −2.083 | p < 0.05 | 3.967 |
| USA | 24 | 24 | 12 | 6 | 0.728 ± 0.062 | 0.023 ± 0.012 | 64 | 7.909 | 52 | 12 | −2.138 | p < 0.05 | 5.890 |
| Italy | 13 | 13 | 8 | 2 | 0.603 ± 0.131 | 0.005 ± 0.002 | 10 | 1.897 | 8 | 2 | −1.643 | p > 0.05 | 0.934 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kapantaidaki, D.E.; Evangelou, V.I.; Morrison, W.R.; Leskey, T.C.; Brodeur, J.; Milonas, P. Halyomorpha halys (Hemiptera: Pentatomidae) Genetic Diversity in North America and Europe. Insects 2019, 10, 174. https://doi.org/10.3390/insects10060174
Kapantaidaki DE, Evangelou VI, Morrison WR, Leskey TC, Brodeur J, Milonas P. Halyomorpha halys (Hemiptera: Pentatomidae) Genetic Diversity in North America and Europe. Insects. 2019; 10(6):174. https://doi.org/10.3390/insects10060174
Chicago/Turabian StyleKapantaidaki, Despoina Ev., Vassiliki I. Evangelou, William R. Morrison, Tracy C. Leskey, Jacques Brodeur, and Panagiotis Milonas. 2019. "Halyomorpha halys (Hemiptera: Pentatomidae) Genetic Diversity in North America and Europe" Insects 10, no. 6: 174. https://doi.org/10.3390/insects10060174
APA StyleKapantaidaki, D. E., Evangelou, V. I., Morrison, W. R., Leskey, T. C., Brodeur, J., & Milonas, P. (2019). Halyomorpha halys (Hemiptera: Pentatomidae) Genetic Diversity in North America and Europe. Insects, 10(6), 174. https://doi.org/10.3390/insects10060174