Distributions of Invasive Arthropods across Heterogeneous Urban Landscapes in Southern California: Aridity as a Key Component of Ecological Resistance



{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Sampling
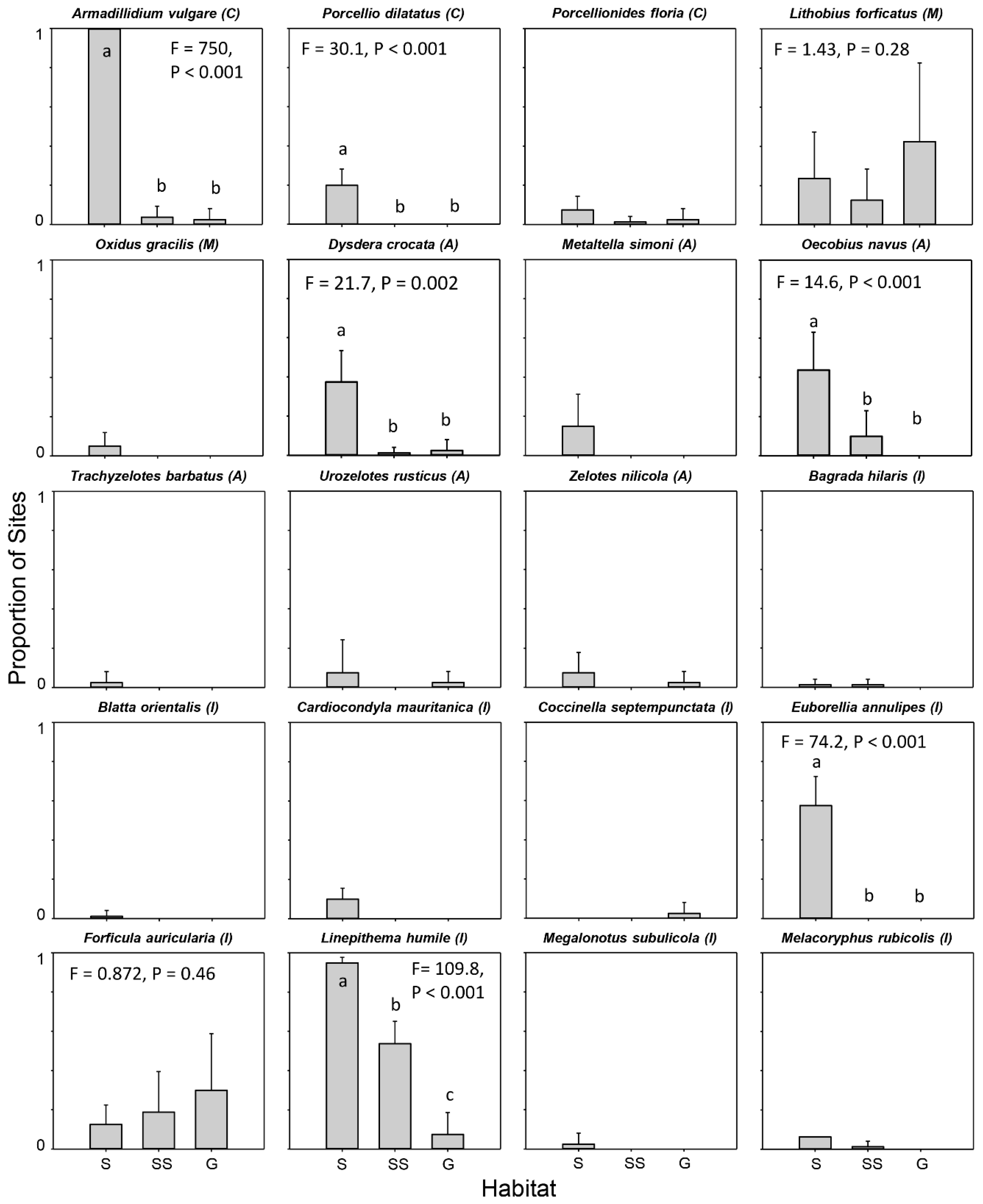
2.3. Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689. [Google Scholar] [CrossRef]
- Lonsdale, W.M. Global patterns of plant invasions and the concept of invasibility. Ecology 1999, 80, 1522. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Blair, R.B.; Launer, A.E. Butterfly diversity and human land use: Species assemblages along an urban gradient. Biol. Conserv. 1997, 80, 113–125. [Google Scholar] [CrossRef]
- Lososová, Z.; Horsák, M.; Chytrý, M.; Čejka, T.; Danihelka, J.; Fajmon, K.; Hájek, O.; Juřičková, L.; Kintrová, K.; Láníková, D.; et al. Diversity of Central European urban biota: Effects of human-made habitat types on plants and land snails. J. Biogeogr. 2011, 38, 1152–1163. [Google Scholar] [CrossRef]
- Spear, D.M.; Adams, T.A.; Boyd, E.S.; Dipman, M.M.; Staubus, W.J.; Meyer, W.M., III. The effects of development, vegetation-type conversion, and fire on low-elevation Southern California spider assemblages. Invertebr. Biol. 2017, 136, 134–145. [Google Scholar] [CrossRef]
- D’Antonio, C.; Levine, J.; Thomsen, M. Ecosystem resistance to invasion and the role of propagule supply: A California perspective. J. Mediterr. Ecol. 2001, 2, 233–245. [Google Scholar]
- Veltman, C.J.; Nee, S.; Crawley, M.J. Correlates of introduction success in exotic New Zealand birds. Am. Nat. 1996, 147, 542–557. [Google Scholar] [CrossRef]
- von Holle, B.; Simberloff, D. Ecological resistance to biological invasion overwhelmed by propagule pressure. Ecology 2005, 86, 3212–3218. [Google Scholar] [CrossRef]
- Colautti, R.I.; Grigorovich, I.A.; MacIsaac, H.J. Propagule pressure: A null model for biological invasions. Biol. Invasions 2006, 8, 1023–1037. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Methuen & Co.: London, UK, 1958; ISBN 9780226206387. [Google Scholar]
- Menke, S.B.; Holway, D.A. Abiotic factors control invasion by Argentine ants at the community scale. J. Anim. Ecol. 2006, 75, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Holway, D.A.; Suarez, A.V.; Case, T.J. Role of abiotic factors in governing susceptibility to invasion: A test with Argentine Ants. Ecology 2002, 83, 1610–1619. [Google Scholar] [CrossRef]
- Soulé, M.E.; Bolger, D.T.; Alberts, A.C.; Wrights, J.; Sorice, M.; Hill, S. Reconstructed dynamics of rapid extinctions of chaparral-requiring birds in urban habitat islands. Conserv. Biol. 1988, 2, 75–92. [Google Scholar] [CrossRef]
- Suarez, A.V.; Bolger, D.T.; Case, T.J. Effects of fragmentation and invasion on native ant communities in coastal southern California. Ecology 1998, 79, 2041–2056. [Google Scholar] [CrossRef]
- Bolger, D.T.; Suarez, A.V.; Crooks, K.R.; Morrison, S.A.; Case, T.J. Arthropods in urban habitat fragments in southern California: Area, age, and edge effects. Ecol. Appl. 2000, 10, 1230–1248. [Google Scholar] [CrossRef]
- Prentice, T.R.; Burger, J.C.; Icenogle, W.R.; Redak, R.A. Spiders from Riversidian coastal sage scrub with comparisons to Diegan scrub fauna (Arachnida: Araneae). Pan-Pac. Entomol. 2001, 77, 90–122. [Google Scholar]
- Staubus, W.J.; Boyd, E.S.; Adams, T.A.; Spear, D.M.; Dipman, M.M.; Meyer, W.M., III. Ant communities in native sage scrub, non-native grassland, and suburban habitats in Los Angeles County, USA: Conservation implications. J. Insect Conserv. 2015, 19, 669–680. [Google Scholar] [CrossRef]
- Alpert, P.; Bone, E.; Holzapfel, C. Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 52–66. [Google Scholar] [CrossRef]
- Zefferman, E.; Stevens, J.T.; Charles, G.K.; Dunbar-Irwin, M.; Emam, T.; Fick, S.; Morales, L.V.; Wolf, K.M.; Young, D.J.N.; Young, T.P. Plant communities in harsh sites are less invaded: A summary of observations and proposed explanations. AoB Plants 2015, 7, 1–21. [Google Scholar] [CrossRef]
- Adams, T.A.; Staubus, W.J.; Meyer, W.M., III. Fire impacts on ant assemblages in California sage scrub. Southwest. Entomol. 2018, 43, 323–334. [Google Scholar] [CrossRef]
- Cardoso, P.; Erwin, T.L.; Borges, P.A.V.; New, T.R. The seven impediments in invertebrate conservation and how to overcome them. Biol. Conserv. 2011, 144, 2647–2655. [Google Scholar] [CrossRef]
- AntWeb. Available online: www.antweb.org/ (accessed on 29 August 2018).
- Hogue, C.L. Insects of the Los Angeles Basin, 2nd ed.; Natural History Museum of Los Angeles: Los Angeles, CA, USA, 1993; ISBN 9780938644446. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Schilman, P.E.; Lighton, J.R.B.; Holway, D.A. Water balance in the Argentine ant (Linepithema humile) compared with five common native ant species from southern California. Physiol. Entomol. 2007, 32, 1–7. [Google Scholar] [CrossRef]
- Wieser, W. Ecophysiological adaptations of terrestrial isopods: A brief review. Symp. Zool. Soc. Lond. 1984, 53, 247–265. [Google Scholar]
- Voss, S.C.; Main, B.Y.; Dadour, I.R. Habitat preferences of the urban wall spider Oecobius navus (Araneae, Oecobiidae). Aust. J. Entomol. 2007, 46, 261–268. [Google Scholar] [CrossRef]
- Cooke, J.A.L. A contribution to the biology of the British spiders belonging to the genus Dysdera. Oikos 1965, 16, 20. [Google Scholar] [CrossRef]
- Human, K.G.; Gordon, D.M. Exploitation and interference competition between the invasive Argentine ant, Linepithema humile, and native ant species. Oecologia 1996, 105, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Human, K.G.; Gordon, D.M. Effects of Argentine ants on invertebrate biodiversity in northern California. Conserv. Biol. 1997, 11, 1242–1248. [Google Scholar] [CrossRef]
- Holway, D.A. Effect of Argentine ant invasions on ground-dwelling arthropods in northern California riparian woodlands. Oecologia 1998, 116, 252–258. [Google Scholar] [CrossRef] [PubMed]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staubus, W.J.; Bird, S.; Meadors, S.; Meyer, W.M., III. Distributions of Invasive Arthropods across Heterogeneous Urban Landscapes in Southern California: Aridity as a Key Component of Ecological Resistance. Insects 2019, 10, 29. https://doi.org/10.3390/insects10010029
Staubus WJ, Bird S, Meadors S, Meyer WM III. Distributions of Invasive Arthropods across Heterogeneous Urban Landscapes in Southern California: Aridity as a Key Component of Ecological Resistance. Insects. 2019; 10(1):29. https://doi.org/10.3390/insects10010029
Chicago/Turabian StyleStaubus, Weston J., Savanah Bird, Savannah Meadors, and Wallace M. Meyer, III. 2019. "Distributions of Invasive Arthropods across Heterogeneous Urban Landscapes in Southern California: Aridity as a Key Component of Ecological Resistance" Insects 10, no. 1: 29. https://doi.org/10.3390/insects10010029
APA StyleStaubus, W. J., Bird, S., Meadors, S., & Meyer, W. M., III. (2019). Distributions of Invasive Arthropods across Heterogeneous Urban Landscapes in Southern California: Aridity as a Key Component of Ecological Resistance. Insects, 10(1), 29. https://doi.org/10.3390/insects10010029