The Dynamics of Deformed Wing Virus Concentration and Host Defensive Gene Expression after Varroa Mite Parasitism in Honey Bees, Apis mellifera

 ,
, 
 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Honey Bees Sample Collection
2.2. Varroa Mite Collection
2.3. Varroa Challenging Experiment
2.4. RNA Extraction and qRT-PCR
2.5. Data Analysis
3. Results
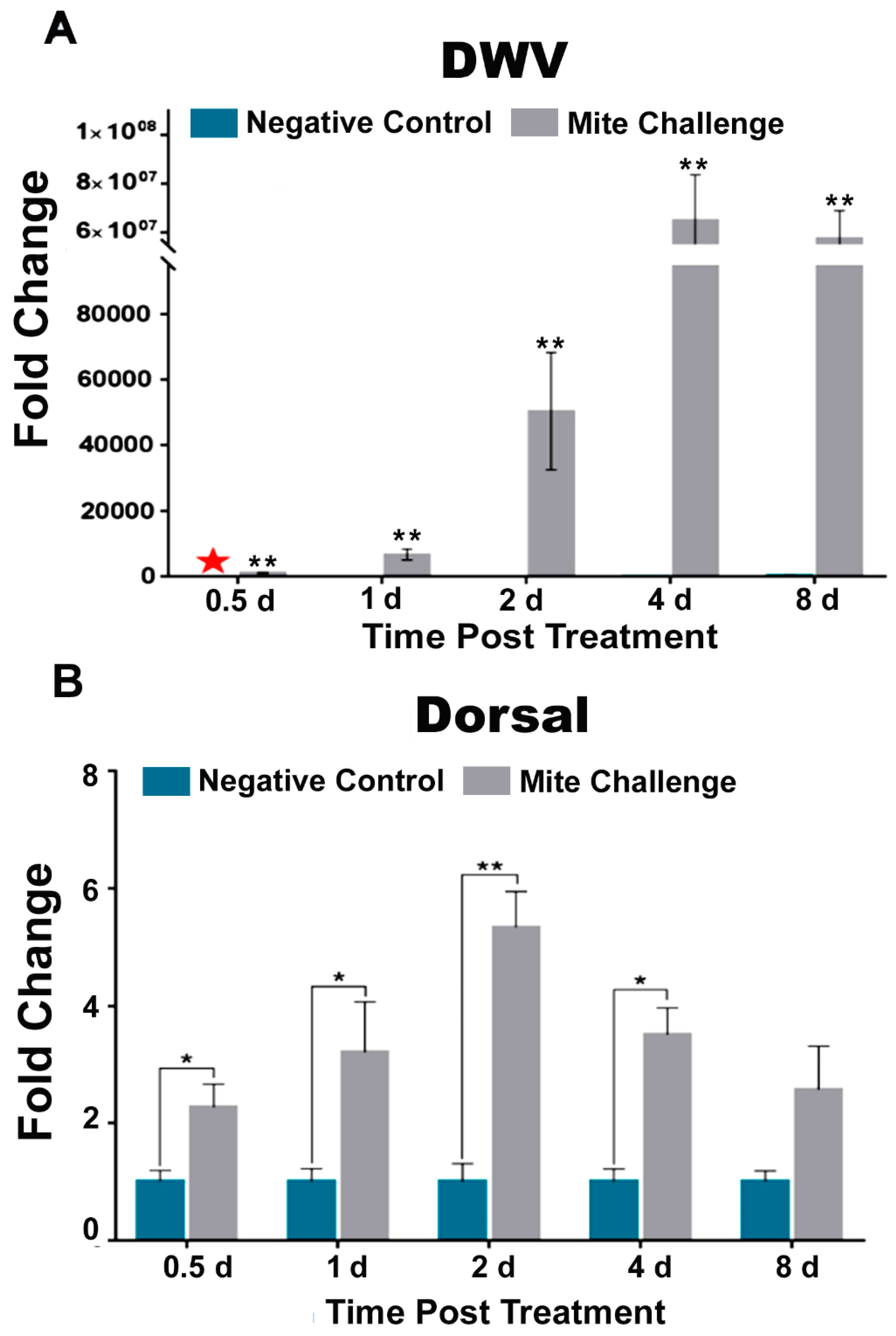
3.1. Rapid Increase in DWV Concentration Coincided with Initial Increase Followed by Subsequent Downregulation of Dorsal and Concomitant AMP
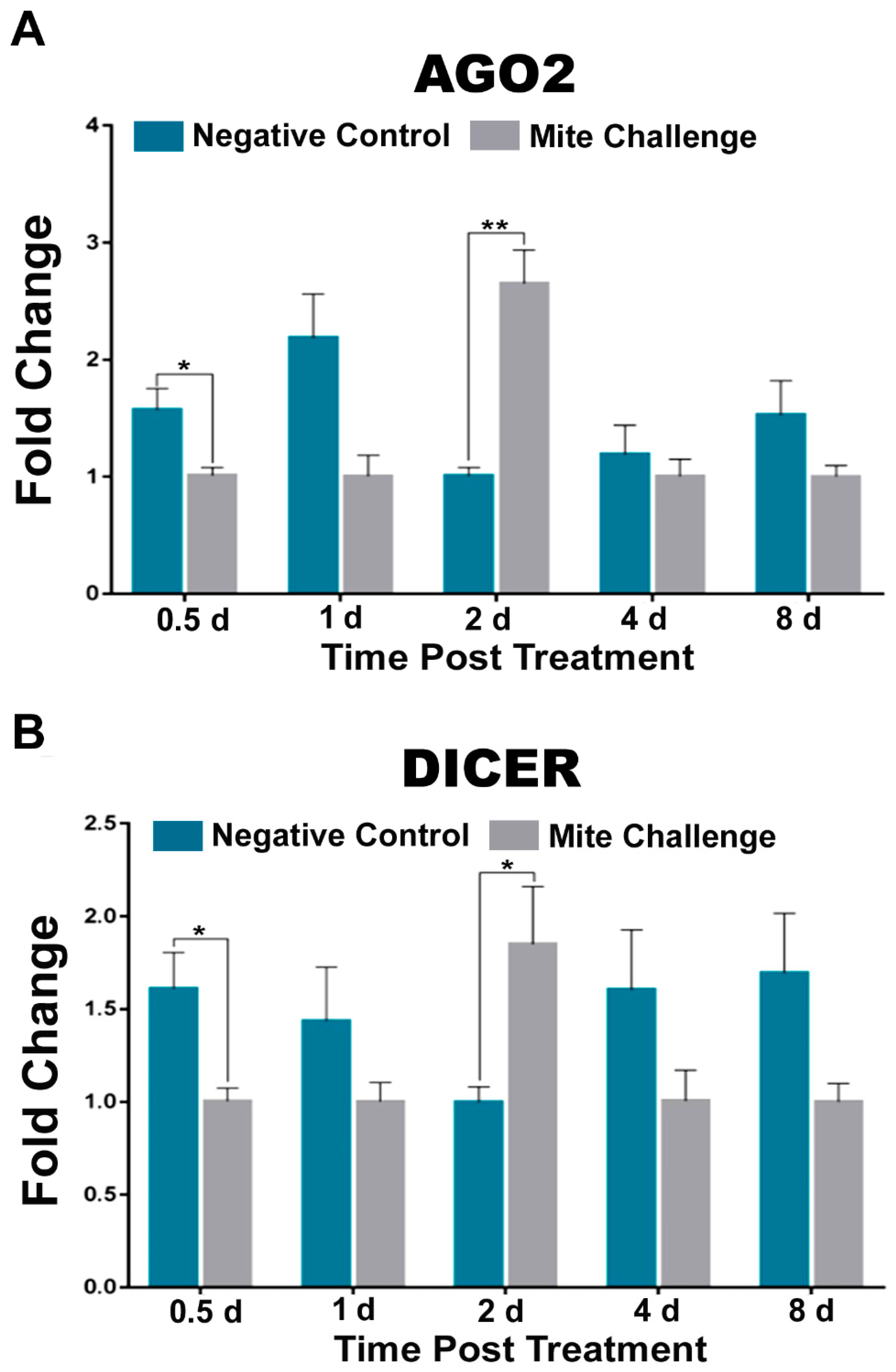
3.2. RNAi Pathway Transcript Levels Peak at Approximately the Same Time Post Challenge as Dorsal and AMPs
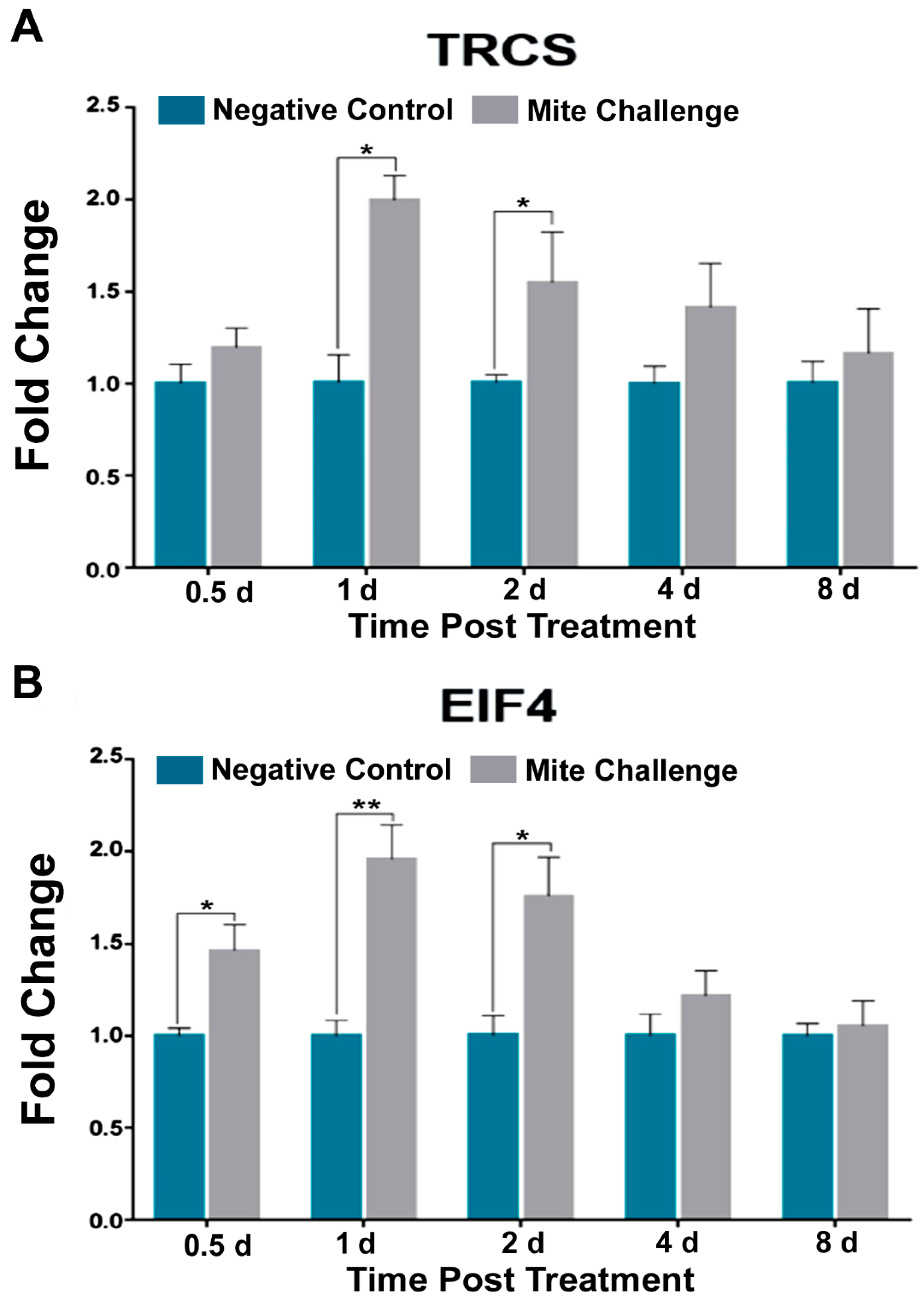
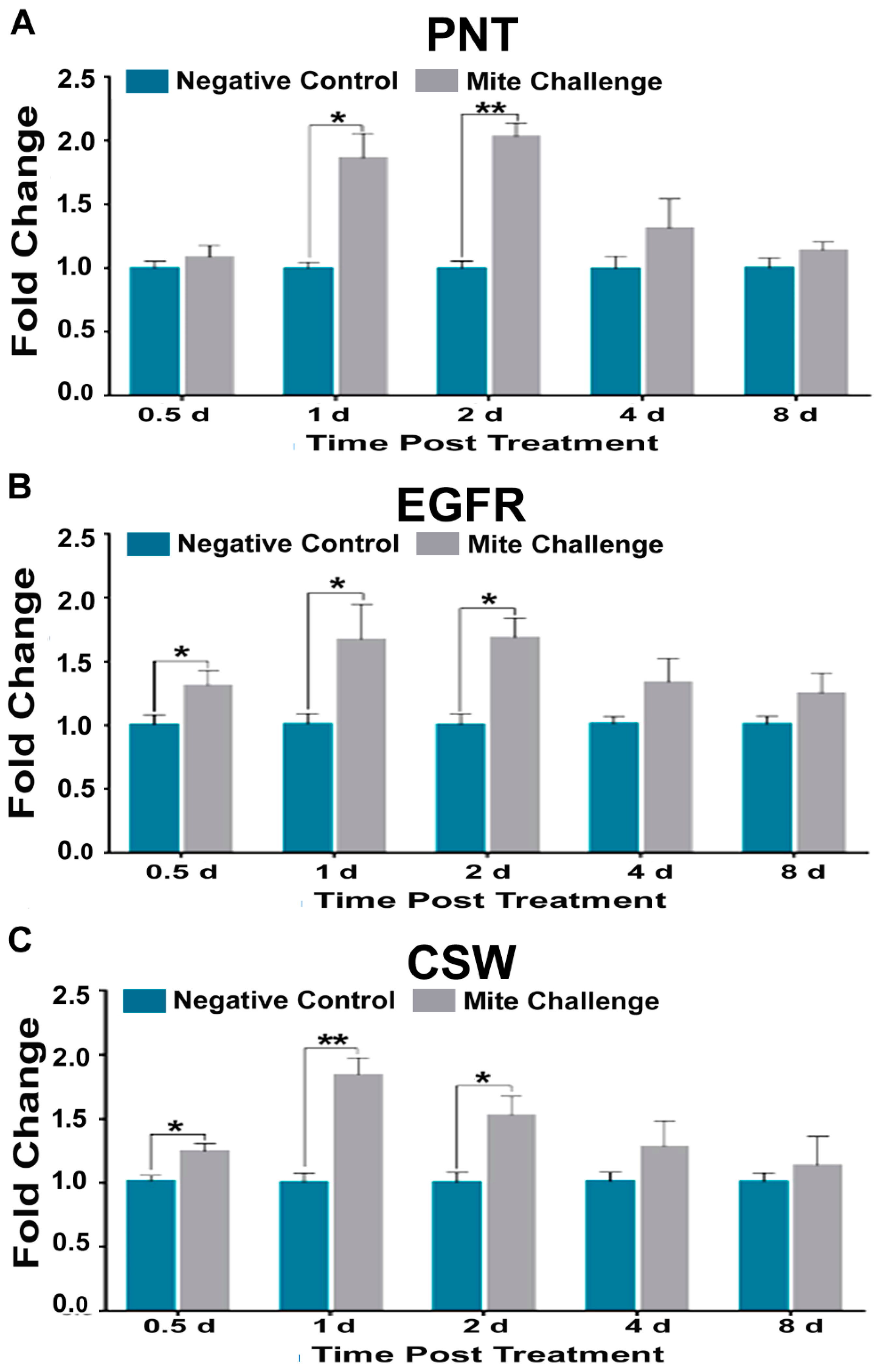
3.3. TOR, Jak-STAT, and EGFR Pathways Mimic, to Varying Degrees, Dorsal Transcription in the Presence of Varroa Infestation
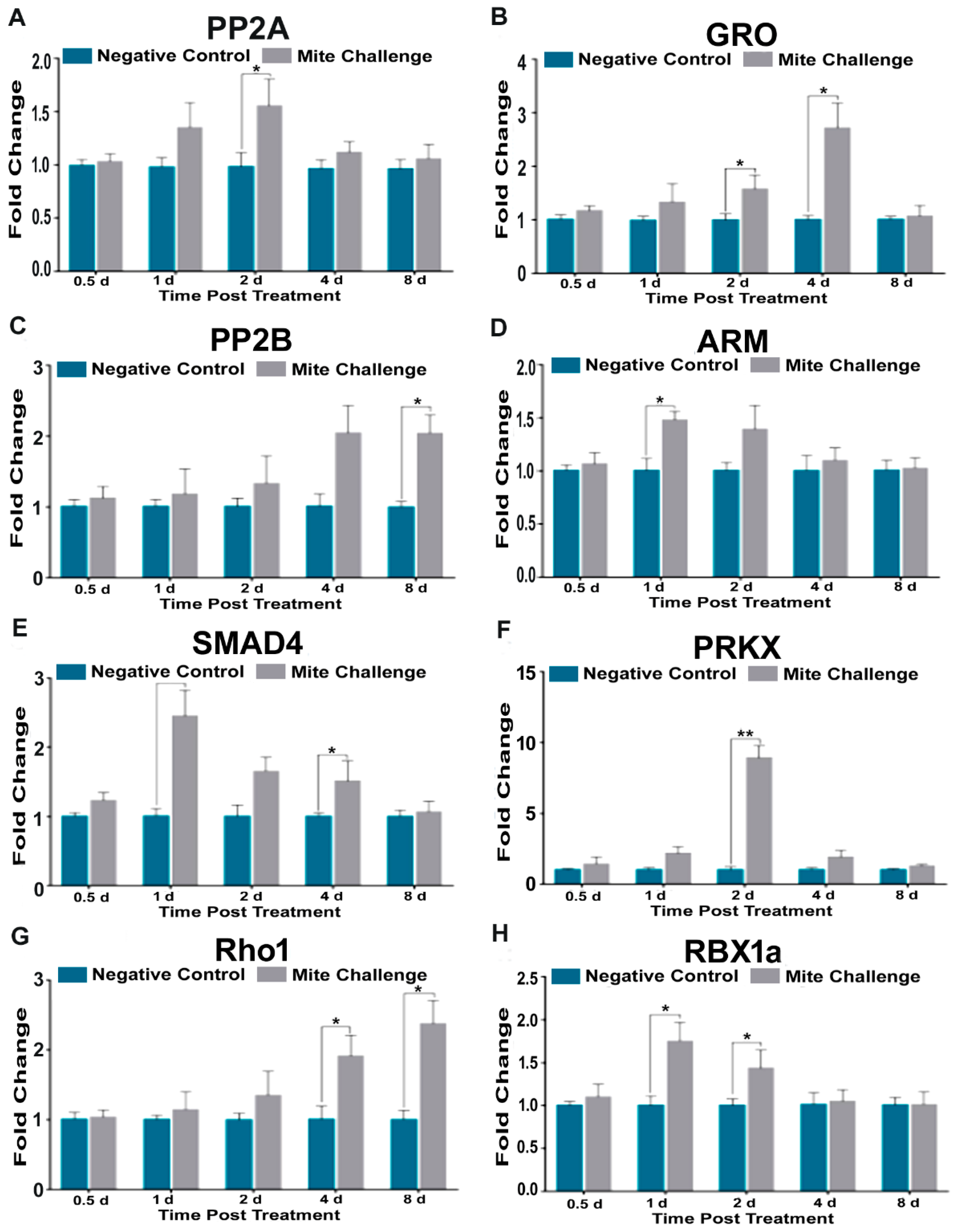
3.4. Groucho, Protein Phosphatase 2b, and Rho1 Upregulation in Late Infections Correspond to High DWV Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gallai, N.; Salles, J.-M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Moritz, R.F.; Erler, S. Lost colonies found in a data mine: Global honey trade but not pests or pesticides as a major cause of regional honeybee colony declines. Agric. Ecosyst. Environ. 2016, 216, 44–50. [Google Scholar] [CrossRef]
- Aizen, M.A.; Harder, L.D. The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef] [PubMed]
- Kelly Kulhanek, N.S.; Rennich, K.; Caron, D.M.; Sagili, R.R.; Pettis, J.S.; Ellis, J.D.; Wilson, M.E.; Wilkes, J.T.; Tarpy, D.R.; Rose, R.; et al. A national survey of managed honey bee 2015–2016 annual colony losses in the USA. J. Apic. Res. 2017, 56, 328–340. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botias, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.-F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A Common Pesticide Decreases Foraging Success and Survival in Honey Bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef]
- Le Féon, V.; Schermann-Legionnet, A.; Delettre, Y.; Aviron, S.; Billeter, R.; Bugter, R.; Hendrickx, F.; Burel, F. Intensification of agriculture, landscape composition and wild bee communities: A large scale study in four European countries. Agric. Ecosyst. Environ. 2010, 137, 143–150. [Google Scholar] [CrossRef]
- Le Conte, Y.; Navajas, M. Climate change: Impact on honey bee populations and diseases. Rev. Sci. Tech. 2008, 27, 499–510. [Google Scholar]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Mondet, F.; de Miranda, J.R.; Kretzschmar, A.; Le Conte, Y.; Mercer, A.R. On the front line: Quantitative virus dynamics in honeybee (Apis mellifera L.) colonies along a new expansion front of the parasite Varroa destructor. PLoS Pathog. 2014, 10, e1004323. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103 (Suppl. 1), S96–S119. [Google Scholar] [CrossRef]
- Villalobos, E.M. The mite that jumped, the bee that traveled, the disease that followed. Science 2016, 351, 554–556. [Google Scholar] [CrossRef] [PubMed]
- Wilfert, L.; Long, G.; Leggett, H.C.; Schmid-Hempel, P.; Butlin, R.; Martin, S.J.; Boots, M. Deformed wing virus is a recent global epidemic in honeybees driven by Varroa mites. Science 2016, 351, 594–597. [Google Scholar] [CrossRef] [PubMed]
- De Miranda, J.R.; Fries, I. Venereal and vertical transmission of deformed wing virus in honeybees (Apis mellifera L.). J. Invertebr. Pathol. 2008, 98, 184–189. [Google Scholar] [CrossRef]
- Brettell, L.E.; Mordecai, G.J.; Schroeder, D.C.; Jones, I.M.; Da Silva, J.R.; Vicente-Rubiano, M.; Martin, S.J. A comparison of deformed wing virus in deformed and asymptomatic honey bees. Insects 2017, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Bowen-Walker, P.L.; Martin, S.J.; Gunn, A. The transmission of deformed wing virus between honeybees (Apis mellifera L.) by the ectoparasitic mite varroa jacobsoni Oud. J. Invertebr. Pathol. 1999, 73, 101–106. [Google Scholar] [CrossRef]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 102, 7470–7475. [Google Scholar] [CrossRef]
- Di Prisco, G.; Annoscia, D.; Margiotta, M.; Ferrara, R.; Varricchio, P.; Zanni, V.; Caprio, E.; Nazzi, F.; Pennacchio, F. A mutualistic symbiosis between a parasitic mite and a pathogenic virus undermines honey bee immunity and health. Proc. Natl. Acad. Sci. USA 2016, 113, 3203–3208. [Google Scholar] [CrossRef]
- Kuster, R.D.; Boncristiani, H.F.; Rueppell, O. Immunogene and viral transcript dynamics during parasitic Varroa destructor mite infection of developing honey bee (Apis mellifera) pupae. J. Exp. Biol. 2014, 217, 1710–1718. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global honey bee viral landscape altered by a parasitic mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.; Jironkin, A.; Chandler, D.; Burroughs, N.; Evans, D.J.; Ryabov, E.V. Recombinants between Deformed wing virus and Varroa destructor virus-1 may prevail in Varroa destructor-infested honeybee colonies. J. Gen. Virol. 2011, 92, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Brutscher, L.M.; Daughenbaugh, K.F.; Flenniken, M.L. Antiviral defense mechanisms in honey bees. Curr. Opin. Insect. Sci. 2015, 10, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.J.; Kenny, N.J.; Dearden, P.K. Components of the dorsal-ventral pathway also contribute to anterior-posterior patterning in honeybee embryos (Apis mellifera). EvoDevo 2014, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Clevers, H. Wnt/beta-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Zhang, X. The Wnt signaling pathway is involved in the regulation of phagocytosis of virus in Drosophila. Sci. Rep. 2013, 3, 2069. [Google Scholar] [CrossRef] [PubMed]
- Nazzi, F.; Brown, S.P.; Annoscia, D.; Del Piccolo, F.; Di Prisco, G.; Varricchio, P.; Della Vedova, G.; Cattonaro, F.; Caprio, E.; Pennacchio, F. Synergistic parasite-pathogen interactions mediated by host immunity can drive the collapse of honeybee colonies. PLoS Pathog. 2012, 8, e1002735. [Google Scholar] [CrossRef]
- Gammon, D.B.; Mello, C.C. RNA interference-mediated antiviral defense in insects. Curr. Opin. Insect Sci. 2015, 8, 111–120. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Chen, Y. Nutrition, immunity and viral infections in honey bees. Curr. Opin. Insect Sci. 2015, 10, 170–176. [Google Scholar] [CrossRef]
- Li, J.; Qin, H.; Wu, J.; Sadd, B.M.; Wang, X.; Evans, J.D.; Peng, W.; Chen, Y. The prevalence of parasites and pathogens in Asian honeybees Apis cerana in China. PLoS ONE 2012, 7, e47955. [Google Scholar] [CrossRef]
- Prisco, G.D.; Zhang, X.; Pennacchio, F.; Caprio, E.; Li, J.; Evans, J.D.; Degrandi-Hoffman, G.; Hamilton, M.; Chen, Y.P. Dynamics of persistent and acute deformed wing virus infections in honey bees, Apis mellifera. Viruses 2011, 3, 2425–2441. [Google Scholar] [CrossRef]
- Di Prisco, G.; Pennacchio, F.; Caprio, E.; Boncristiani, H.F., Jr.; Evans, J.D.; Chen, Y. Varroa destructor is an effective vector of Israeli acute paralysis virus in the honeybee, Apis mellifera. J. Gen. Virol. 2011, 92, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.P.; Pettis, J.S.; Evans, J.D.; Kramer, M.; Feldlaufer, M.F. Molecular evidence for transmission of Kashmir bee virus in honey bee colonies by ectoparasitic mite, Varroa destructor. Apidologies 2004, 35, 441–448. [Google Scholar] [CrossRef]
- Evans, J.D.; Chen, Y.P.; Di Prisco, G.; Pettis, J.; Williams, V. Bee cups: Single-use cages for honey bee experiments. J. Apic. Res. 2009, 48, 300–302. [Google Scholar] [CrossRef]
- Simone, M.; Evans, J.D.; Spivak, M. Resin collection and social immunity in honey bees. Evolution Int. J. Org. Evolution 2009, 63, 3016–3022. [Google Scholar] [CrossRef] [PubMed]
- Boncristiani, H.F.; Evans, J.D.; Chen, Y.; Pettis, J.; Murphy, C.; Lopez D., L.; Simone-Finstrom, M.; Strand, M.; Tarpy, D.R.; Rueppell, O. In vitro infection of pupae with Israeli acute paralysis virus suggests disturbance of transcriptional homeostasis in honey bees (Apis mellifera). PLoS One 2013, 8, e73429. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Jang, H.D.; Yoon, K.; Shin, Y.J.; Kim, J.; Lee, S.Y. PIAS3 suppresses NF-κB-mediated transcription by interacting with the p65/RelA subunit. J. Biol. Chem. 2004, 279, 24873–24880. [Google Scholar] [CrossRef]
- Gatschenberger, H.; Gimple, O.; Tautz, J.; Beier, H. Honey bee drones maintain humoral immune competence throughout all life stages in the absence of vitellogenin production. J. Exp. Biol. 2012, 215, 1313–1322. [Google Scholar] [CrossRef]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Evans, J.D.; Huang, Q.; Rodríguez-García, C.; Liu, J.; Hamilton, M.; Grozinger, C.M.; Webster, T.C.; Su, S.; Chen, Y.P. Silencing the Honey Bee (Apis mellifera) Naked Cuticle Gene (nkd) Improves Host Immune Function and Reduces Nosema ceranae Infections. Appl. Eviron. Microbiol. 2016, 82, 6779–6787. [Google Scholar] [CrossRef] [PubMed]
- Zanni, V.; Galbraith, D.A.; Annoscia, D.; Grozinger, C.M.; Nazzi, F. Transcriptional signatures of parasitization and markers of colony decline in Varroa-infested honey bees (Apis mellifera). Insect Biochem. Mol. Biol. 2017, 87, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bridges, R.; Wortham, A.; Kulesz-Martin, M. NF-κB Repression by PIAS3 Mediated RelA SUMOylation. PLoS ONE 2012, 7, e37636. [Google Scholar] [CrossRef] [PubMed]
- Lupberger, J.; Zeisel, M.B.; Xiao, F.; Thumann, C.; Fofana, I.; Zona, L.; Davis, C.; Mee, C.J.; Turek, M.; Gorke, S. EGFR and EphA2 are host factors for hepatitis C virus entry and possible targets for antiviral therapy. Nat. Med. 2011, 17, 589. [Google Scholar] [CrossRef] [PubMed]
- Maori, E.; Paldi, N.; Shafir, S.; Kalev, H.; Tsur, E.; Glick, E.; Sela, I. IAPV, a bee-affecting virus associated with colony collapse disorder can be silenced by dsRNA ingestion. Insect Mol. Biol. 2009, 18, 55–60. [Google Scholar] [CrossRef]
- Hunter, W.; Ellis, J.; vanEngelsdorp, D.; Hayes, J.; Westervelt, D.; Glick, E.; Williams, M.; Sela, I.; Maori, E.; Pettis, J.; et al. Large-Scale Field Application of RNAi Technology Reducing Israeli Acute Paralysis Virus Disease in Honey Bees (Apis mellifera, Hymenoptera: Apidae). PLoS Pathog. 2010, 6, e1001160. [Google Scholar] [CrossRef]
- Chen, Y.; Evans, J.D. RNAi in treating honey bee diseases. Bee Cult. 2012, 140, 27–29. [Google Scholar]
- Desai, S.D.; Eu, Y.J.; Whyard, S.; Currie, R.W. Reduction in deformed wing virus infection in larval and adult honey bees (Apis mellifera L.) by double-stranded RNA ingestion. Insect Mol. Biol. 2012, 21, 446–455. [Google Scholar] [CrossRef]
- Wang, X.-H.; Aliyari, R.; Li, W.-X.; Li, H.-W.; Kim, K.; Carthew, R.; Atkinson, P.; Ding, S.-W. RNA interference directs innate immunity against viruses in adult Drosophila. Science 2006, 312, 452–454. [Google Scholar] [CrossRef]
- Campbell, C.L.; Keene, K.M.; Brackney, D.E.; Olson, K.E.; Blair, C.D.; Wilusz, J.; Foy, B.D. Aedes aegypti uses RNA interference in defense against Sindbis virus infection. BMC Microbiol. 2008, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vargas, I.; Scott, J.C.; Poole-Smith, B.K.; Franz, A.W.; Barbosa-Solomieu, V.; Wilusz, J.; Olson, K.E.; Blair, C.D. Dengue virus type 2 infections of Aedes aegypti are modulated by the mosquito’s RNA interference pathway. PLoS Pathog. 2009, 5, e1000299. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.D.; Kraft, J.J.; Miller, W.A.; Goss, D.J. Recruitment of the 40S ribosome subunit to the 3′-untranslated region (UTR) of a viral mRNA, via the eIF4 complex, facilitates cap-independent translation. J. Biol. Chem. 2015, 290, 11268–11281. [Google Scholar] [CrossRef]
- Lamphear, B.J.; Yan, R.; Yang, F.; Waters, D.; Liebig, H.D.; Klump, H.; Kuechler, E.; Skern, T.; Rhoads, R.E. Mapping the cleavage site in protein synthesis initiation factor eIF-4 gamma of the 2A proteases from human Coxsackievirus and rhinovirus. J. Biol. Chem. 1993, 268, 19200–19203. [Google Scholar] [PubMed]
- Kirchweger, R.; Ziegler, E.; Lamphear, B.; Waters, D.; Liebig, H.; Sommergruber, W.; Sobrino, F.; Hohenadl, C.; Blaas, D.; Rhoads, R. Foot-and-mouth disease virus leader proteinase: Purification of the Lb form and determination of its cleavage site on eIF-4 gamma. J. Virol. 1994, 68, 5677–5684. [Google Scholar] [PubMed]
- Martin, K.M.; Barandoc-Alviar, K.; Schneweis, D.J.; Stewart, C.L.; Rotenberg, D.; Whitfield, A.E. Transcriptomic response of the insect vector, Peregrinus maidis, to Maize mosaic rhabdovirus and identification of conserved responses to propagative viruses in hopper vectors. Virology 2017, 509, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Annoscia, D.; Zanni, V.; Galbraith, D.; Quirici, A.; Grozinger, C.; Bortolomeazzi, R.; Nazzi, F. Elucidating the mechanisms underlying the beneficial health effects of dietary pollen on honey bees (Apis mellifera) infested by Varroa mite ectoparasites. Sci. Rep. 2017, 7, 6258. [Google Scholar] [CrossRef]
- Verginelli, F.; Perin, A.; Dali, R.; Fung, K.H.; Lo, R.; Longatti, P.; Guiot, M.-C.; Del Maestro, R.F.; Rossi, S.; Di Porzio, U. Transcription factors FOXG1 and Groucho/TLE promote glioblastoma growth. Nat. Commun. 2013, 4, 2956. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Abbr. | Accession No. | Primers (5′-3′) | Tm (°C) | Pathway | Reference |
|---|---|---|---|---|---|---|
| Actin related protein 1 | β-actin | NM_001185145 | F: TTGTATGCCAACACTGTCCTTT | — | Reference | [36] |
| R: TGGCGCGATGATCTTAATTT | ||||||
| Deformed wing virus | DWV | NC_004830.2 | F: CGAAACCAACTTCTGAGGAA | 55 | Viral | [37] |
| R: GTGTTGATCCCTGAGGCTTA | ||||||
| Signal transducer and activator of transcription 5B | STAT | XM_397181 | F: ATCAGTGGCTTCCAGCTACG | 59 | Jak-STAT | This study |
| R: ATCGAAGGCAGCTCAGGATG | ||||||
| E3 SUMO-protein ligase PIAS3 | PIAS3 | XM_623568 | F: ACCACCTGCTCATCAAGCAT | 59 | Jak-STAT | This study |
| R: AGCTGCACTTGTTGTTTGTTGT | ||||||
| Signal transducing adapter molecule 1 | STAM | XM_623536 | F: GTCAGCCAGTACCGGAACAA | 59 | Jak-STAT | This study |
| R: AGGTTGTTGGCTTACCGGAG | ||||||
| Tyrosine-protein kinase hopscotch | HOP | XM_001121783 | F: TTGTGCTCCTGAAAATGCTG | 59 | Jak-STAT | This study |
| R: AACCTCCAAATCGCTCTGTG | ||||||
| E3 ubiquitin-protein ligase CBL | CBL | XM_016911574 | F: CCGGACACCGAGGAAATCAT | 59 | Jak-STAT | This study |
| R: CGGTGGTGGAGCATTCTCTT | ||||||
| ETS-like protein pointed | PNT | XM_016914525 | F: TATACCGCGTTAGTGCTGCC | 59 | MAPK | This study |
| R: CTAGCATCCCGCTTGATGGA | ||||||
| Epidermal growth factor receptor-like | EGFR | XM_006560026 | F: GTGAACAGTGCGAAGACGAA | 59 | MAPK | This study |
| R: GGAACAATACGGTTCGCTGT | ||||||
| Tyrosine-protein phosphatase corkscrew | CSW | XM_006561366 | F: TTGCTGCTTCTCTTGCTTCA | 59 | MAPK | This study |
| R: GTTCTGCTTGCATTCGTTGA | ||||||
| Target of rapamycin complex subunit lst8 | TRCS | XM_393223 | F: TGTGGATGGCACTAACAGCA | 59 | TOR | This study |
| R: ACCCTCTTCCTGAAAACCCA | ||||||
| Eukaryotic translation initiation factor 4 E type 3-A-like | EIF4 | XM_392604 | F: TGGTGCTTGCAGCTATTGGA | 59 | TOR | This study |
| R: GGCATGATGTGATTGATGAGGTTT | ||||||
| Embryonic polarity protein | Dorsal | XM_016913118 | F: CTCATCGGAAGACATGACAGTGA | 59 | NF-kB | This study |
| R: TGAATTCAAAGCCAGTTCGAAAA | ||||||
| Protein argonaute-2 | AGO2 | XM_395048 | F: ACCTGCTGAGTTATGCACAGT | 59 | RNAi | This study |
| R: AGCCTTTAGAACTCTTGCTGGT | ||||||
| Endoribonuclease Dicer | Dicer | XM_016917734 | F: AGCAGTAGCTGATTGTGTGGA | 59 | RNAi | This study |
| R: TGAAGGATGTGTAAACGCCTGT | ||||||
| Hymenoptaecin | HYMEN | NM_001011615 | F: CTCTTCTGTGCCGTTGCATA | 59 | Toll | This study |
| R: GCGTCTCCTGTCATTCCATT | ||||||
| Abaecin | ABAE | NM_001011617 | F: ATCTTCGCACTACTCGCCAC | 59 | Toll | This study |
| R: AGCCTTGAGGCCATTTAATTTTCG | ||||||
| Apidaecin | APID | XM_006572699 | F: GGCACGAGAAGAATTTTGCCT | 59 | Toll | This study |
| R: GAAGGCGCGTAGGTCGAGTA | ||||||
| Defensin 1 | Def1 | NM_001011616 | F: TGCGCTGCTAACTGTCTCAG | 59 | Toll | This study |
| R: AATGGCACTTAACCGAAACG | ||||||
| Protein groucho-1 | PG1 | XM_006565410 | F: ACAAGGATATAGGCCATAGCGAC | 59 | Wnt | This study |
| R: GGTGGCGGGACTTCTTTCTT | ||||||
| Armadillo segment polarity protein | ARM | XM_006557800 | F: ATTCGAGGCAAGAAAGGCC | 59 | Wnt | This study |
| R: ATCTACCCACACCGTATCGC | ||||||
| cAMP-dependent protein kinase catalytic subunit PRKX | PRKX | XM_393711 | F: ATATAGTCGAGCGACGAGCG | 59 | Wnt | This study |
| R: CGAAAGTGCCTGTACCTATCGT | ||||||
| RING-box protein 1A | RBP | XM_016910739 | F: GTGGGAGTTCCAAAAATACGGT | 59 | Wnt | This study |
| R: GAAGTGCTGCAAAGACGCTC | ||||||
| Protein phosphatase 2A at 29B | PP2A | XM_006562757 | F: AGGGTAATGCTTCCAACAGTCT | 59 | Wnt | This study |
| R: CTTGTTCAACCTGCGATGCC | ||||||
| Protein phosphatase 2B at 14D | PP2B | XM_016911698 | F: TCCTCCTGTTTGTGTGGCAG | 59 | Wnt | This study |
| R: GCTTTCCTCTCTCAGGACGG | ||||||
| Mothers against decapentaplegic homolog 4 | SMAD4 | XM_006563458 | F: CACACCGTAGGTAGCCAACA | 59 | Wnt | This study |
| R:GCAGTACCTACTAAAGATGCTGCT | ||||||
| Ras-like GTP-binding protein Rho1 | Rho1 | XM_016911392 | F: GCGTGTGAGTGTCAAGCTGT | 59 | Wnt | This study |
| R: CCTTCAAATCCAGCTCTTGC |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Heerman, M.; Peng, W.; Evans, J.D.; Rose, R.; DeGrandi-Hoffman, G.; Simone-Finstrom, M.; Li, J.; Li, Z.; Cook, S.C.; et al. The Dynamics of Deformed Wing Virus Concentration and Host Defensive Gene Expression after Varroa Mite Parasitism in Honey Bees, Apis mellifera. Insects 2019, 10, 16. https://doi.org/10.3390/insects10010016
Zhao Y, Heerman M, Peng W, Evans JD, Rose R, DeGrandi-Hoffman G, Simone-Finstrom M, Li J, Li Z, Cook SC, et al. The Dynamics of Deformed Wing Virus Concentration and Host Defensive Gene Expression after Varroa Mite Parasitism in Honey Bees, Apis mellifera. Insects. 2019; 10(1):16. https://doi.org/10.3390/insects10010016
Chicago/Turabian StyleZhao, Yazhou, Matthew Heerman, Wenjun Peng, Jay D. Evans, Robyn Rose, Gloria DeGrandi-Hoffman, Michael Simone-Finstrom, Jianghong Li, Zhiguo Li, Steven C. Cook, and et al. 2019. "The Dynamics of Deformed Wing Virus Concentration and Host Defensive Gene Expression after Varroa Mite Parasitism in Honey Bees, Apis mellifera" Insects 10, no. 1: 16. https://doi.org/10.3390/insects10010016
APA StyleZhao, Y., Heerman, M., Peng, W., Evans, J. D., Rose, R., DeGrandi-Hoffman, G., Simone-Finstrom, M., Li, J., Li, Z., Cook, S. C., Su, S., Rodríguez-García, C., Banmeke, O., Hamilton, M., & Chen, Y. (2019). The Dynamics of Deformed Wing Virus Concentration and Host Defensive Gene Expression after Varroa Mite Parasitism in Honey Bees, Apis mellifera. Insects, 10(1), 16. https://doi.org/10.3390/insects10010016