
Post-Mortem Detection of SARS-CoV-2 RNA in Long-Buried Lung Samples

 ,
,  ,
,  ,
,  ,
, 
 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients, Autopsies and Sample Collection
2.2. RNA Extraction
2.3. RNA Quantification and Reverse Transcription
2.4. RT-qPCR and ddPCR Molecular Analyses
2.5. End-Point PCR
2.6. Sequencing
2.7. Correlation Analysis
2.8. Statistical Analyses
3. Results
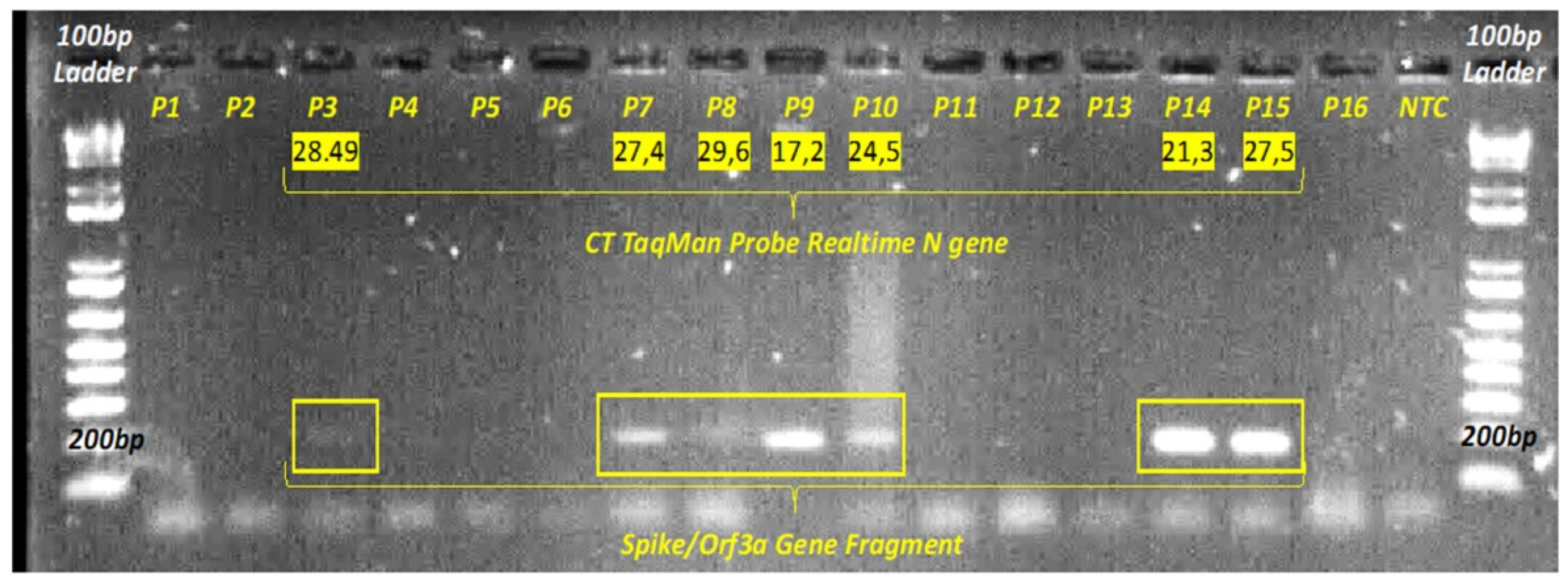
3.1. RT-qPCR and ddPCR SARS-CoV-2 Detection
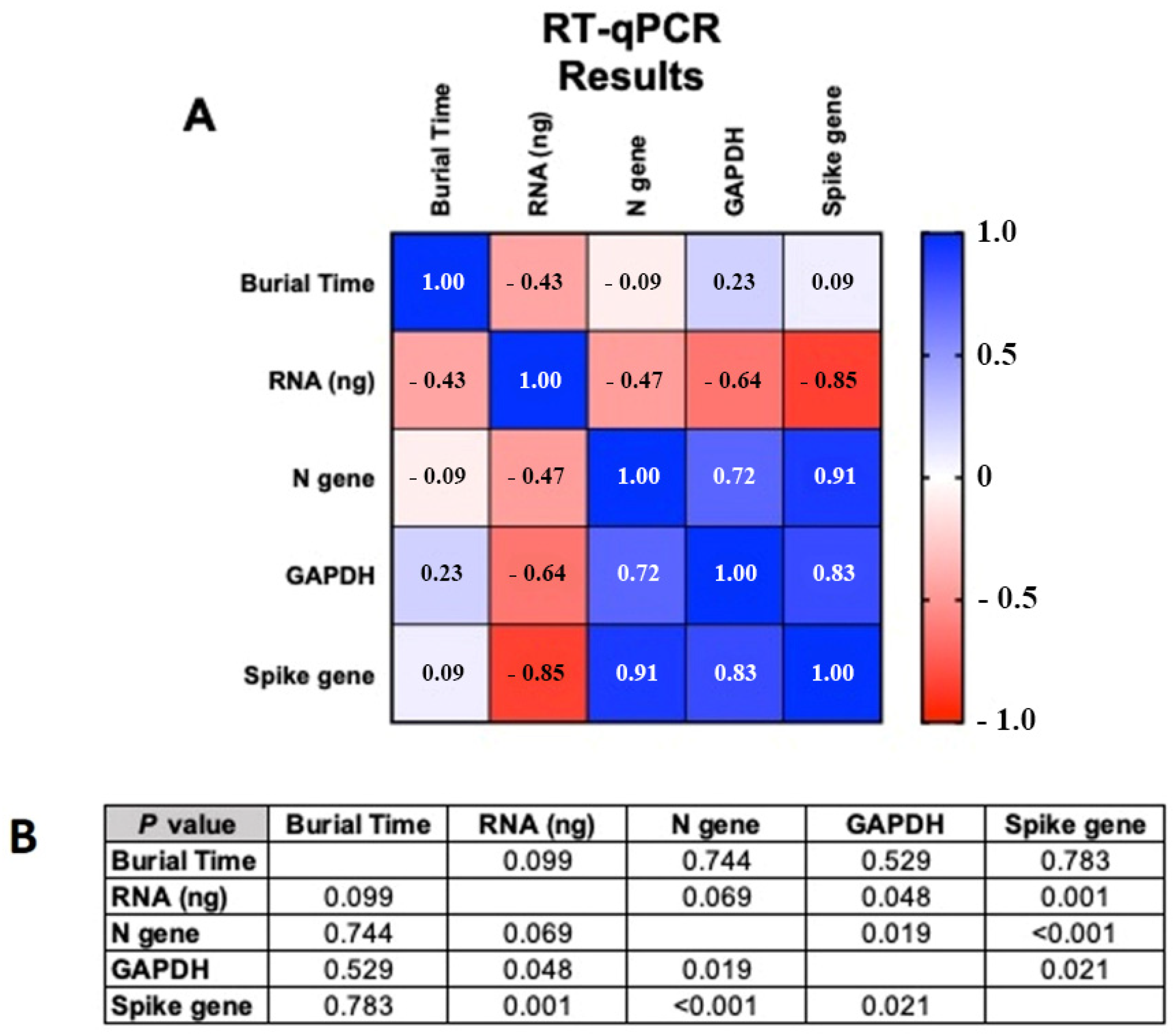
3.2. Correlation Analysis
3.3. SARS-CoV-2 Sequencing
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsatsakis, A.; Calina, D.; Falzone, L.; Petrakis, D.; Mitrut, R.; Siokas, V.; Pennisi, M.; Lanza, G.; Libra, M.; Doukas, S.G.; et al. SARS-CoV-2 pathophysiology and its clinical implications: An integrative overview of the pharmacotherapeutic management of COVID-19. Food Chem. Toxicol. 2020, 146, 111769. [Google Scholar] [CrossRef] [PubMed]
- Pomara, C.; Salerno, M.; Sessa, F.; Esposito, M.; Barchitta, M.; Ledda, C.; Grassi, P.; Liberto, A.; Mattaliano, A.R.; Rapisarda, V.; et al. Safe Management Strategies in Clinical Forensic Autopsies of Confirmed COVID-19 Cases. Diagnostics 2021, 11, 457. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Infection Prevention and Control for the safe management of a dead body in the context of COVID-19. J. Hosp. Infect. 2020, 104, 246–251. [Google Scholar]
- Salerno, M.; Sessa, F.; Piscopo, A.; Montana, A.; Torrisi, M.; Patanè, F.; Murabito, P.; Li Volti, G.; Pomara, C. No Autopsies on COVID-19 Deaths: A Missed Opportunity and the Lockdown of Science. J. Clin. Med. 2020, 9, 1472. [Google Scholar] [CrossRef]
- Sessa, F.; Bertozzi, G.; Cipolloni, L.; Baldari, B.; Cantatore, S.; D’Errico, S.; Di Mizio, G.; Asmundo, A.; Castorina, S.; Salerno, M.; et al. Clinical-Forensic Autopsy Findings to Defeat COVID-19 Disease: A Literature Review. J. Clin. Med. 2020, 9, 2026. [Google Scholar] [CrossRef] [PubMed]
- Cipolloni, L.; Sessa, F.; Bertozzi, G.; Baldari, B.; Cantatore, S.; Testi, R.; D’Errico, S.; Di Mizio, G.; Asmundo, A.; Castorina, S.; et al. Preliminary post-mortem COVID-19 evidence of endothelial injury and factor VIII hyperexpression. Diagnostics 2020, 10, 575. [Google Scholar] [CrossRef] [PubMed]
- D’Errico, S.D.; Zanon, M.; Montanaro, M.; Radaelli, D.; Sessa, F.; Mizio, G.; Di Montana, A.; Corrao, S.; Salerno, M.; Pomara, C. More than Pneumonia: Distinctive Features of SARS-Cov-2 Infection. From Autopsy Findings to Clinical Implications: A Systematic Review. Microorganisms 2020, 8, 1642. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Nolan, T. RT-qPCR Testing of SARS-CoV-2: A Primer. Int. J. Mol. Sci. 2020, 21, 3004. [Google Scholar] [CrossRef]
- Falzone, L.; Gattuso, G.; Tsatsakis, A.; Spandidos, D.A.; Libra, M. Current and innovative methods for the diagnosis of COVID-19 infection (Review). Int. J. Mol. Med. 2021, 47, 100. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention Collection and Submission of Postmortem Specimens from Deceased Persons with Known or Suspected COVID-19. Available online: https://eaaf.org/wp-content/uploads/covid19-PDFs/EEUU/CDC-guidance-postmortem-specimens.pdf (accessed on 18 August 2020).
- Pomara, C.; Fineschi, V. Forensic and Clinical Forensic Autopsy. An Atlas and Handbook, 2nd ed.; Pomara, C., Fineschi, V., Eds.; CRC Press: Boca Raton, FL, USA, 2020; ISBN 9780367330712. [Google Scholar]
- Falzone, L.; Musso, N.; Gattuso, G.; Bongiorno, D.; Palermo, C.I.; Scalia, G.; Libra, M.; Stefani, S. Sensitivity assessment of droplet digital PCR for SARS-CoV-2 detection. Int. J. Mol. Med. 2020, 46, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-P.; Wang, M.; Wang, Y.; Zhu, J.; Zhang, N. Focus on the 2019 novel coronavirus (SARS-CoV-2). Future Microbiol. 2020, 15, 905–918. [Google Scholar] [CrossRef]
- Carpenter, T.O.; Ellis, B.K.; Insogna, K.L.; Philbrick, W.M.; Sterpka, J.; Shimkets, R. Fibroblast Growth Factor 7: An Inhibitor of Phosphate Transport Derived from Oncogenic Osteomalacia-Causing Tumors. J. Clin. Endocrinol. Metab. 2005, 90, 1012–1020. [Google Scholar] [CrossRef] [Green Version]
- Crimi, S.; Falzone, L.; Gattuso, G.; Grillo, C.M.; Candido, S.; Bianchi, A.; Libra, M. Droplet Digital PCR Analysis of Liquid Biopsy Samples Unveils the Diagnostic Role of hsa-miR-133a-3p and hsa-miR-375-3p in Oral Cancer. Biology 2020, 9, 379. [Google Scholar] [CrossRef]
- Salemi, R.; Falzone, L.; Madonna, G.; Polesel, J.; Cinà, D.; Mallardo, D.; Ascierto, P.A.; Libra, M.; Candido, S. MMP-9 as a Candidate Marker of Response to BRAF Inhibitors in Melanoma Patients with BRAF V600E Mutation Detected in Circulating-Free DNA. Front. Pharmacol. 2018, 9, 856. [Google Scholar] [CrossRef]
- Musso, N.; Caronia, F.P.; Castorina, S.; Lo Monte, A.I.; Barresi, V.; Condorelli, D.F. Somatic loss of an EXT2 gene mutation during malignant progression in a patient with hereditary multiple osteochondromas. Cancer Genet. 2015, 208, 62–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steele, J.D.; Meyer, M.; Ebmeier, K.P. Neural predictive error signal correlates with depressive illness severity in a game paradigm. Neuroimage 2004, 23, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Utsunomiya, Y.T.; Utsunomiya, A.T.H.; Torrecilha, R.B.P.; de Paulan, S.C.; Milanesi, M.; Garcia, J.F. Growth Rate and Acceleration Analysis of the COVID-19 Pandemic Reveals the Effect of Public Health Measures in Real Time. Front. Med. 2020, 7, 247. [Google Scholar] [CrossRef]
- Yaacoub, S.; Schünemann, H.J.; Khabsa, J.; El-Harakeh, A.; Khamis, A.M.; Chamseddine, F.; El Khoury, R.; Saad, Z.; Hneiny, L.; Cuello Garcia, C.; et al. Safe management of bodies of deceased persons with suspected or confirmed COVID-19: A rapid systematic review. BMJ Glob. Health 2020, 5, e002650. [Google Scholar] [CrossRef] [PubMed]
- Sessa, F.; Salerno, M.; Pomara, C. Autopsy Tool in Unknown Diseases: The Experience with Coronaviruses (SARS-CoV, MERS-CoV, SARS-CoV-2). Medicina 2021, 57, 309. [Google Scholar] [CrossRef] [PubMed]
- Barth, R.F.; Xu, X.; Buja, L.M. A Call to Action: The Need for Autopsies to Determine the Full Extent of Organ Involvement Associated With COVID-19. Chest 2020, 158, 43–44. [Google Scholar] [CrossRef] [PubMed]
- Mahallawi, W.H. Case report: Detection of the Middle East respiratory syndrome corona virus (MERS-CoV) in nasal secretions of a dead human. J. Taibah. Univ. Med. Sci. 2018, 13, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Gabbrielli, M.; Gandolfo, C.; Anichini, G.; Candelori, T.; Benvenuti, M.; Savellini, G.G.; Cusi, M.G. How long can SARS-CoV-2 persist in human corpses? Int. J. Infect. Dis. 2021, 106, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Hyde, J.L.; Diamond, M.S. Innate immune restriction and antagonism of viral RNA lacking 2′-O methylation. Virology 2015, 479–480, 66–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, M.; Ruggiero, A.; Squeglia, F.; Maga, G.; Berisio, R. A Structural View of SARS-CoV-2 RNA Replication Machinery: RNA Synthesis, Proofreading and Final Capping. Cells 2020, 9, 1267. [Google Scholar] [CrossRef]
- Lazzaroni, M.G.; Piantoni, S.; Masneri, S.; Garrafa, E.; Martini, G.; Tincani, A.; Andreoli, L.; Franceschini, F. Coagulation dysfunction in COVID-19: The interplay between inflammation, viral infection and the coagulation system. Blood Rev. 2021, 46, 100745. [Google Scholar] [CrossRef] [PubMed]
- Battinelli, E.M. COVID-19 concerns aggregate around platelets. Blood 2020, 136, 1221–1223. [Google Scholar] [CrossRef]
- Pomara, C.; Sessa, F.; Ciaccio, M.; Dieli, F.; Esposito, M.; Giammanco, G.M.; Garozzo, S.F.; Giarratano, A.; Prati, D.; Rappa, F.; et al. COVID-19 Vaccine and Death: Causality Algorithm According to the WHO Eligibility Diagnosis. Diagnostics 2021, 11, 955. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | ng/µL | ng for cDNA Synthesis |
|---|---|---|
| P1 | 8.96 | 44.80 |
| P2 | 0.88 | 4.40 |
| P3 | 1.41 | 7.06 |
| P4 | 1.47 | 7.36 |
| P5 | 0.94 | 4.72 |
| P6 | 0.93 | 4.63 |
| P7 | 2.05 | 10.24 |
| P8 | 2.19 | 10.96 |
| P9 | 7.04 | 35.20 |
| P10 | 6.24 | 31.20 |
| P11 | 5.04 | 25.20 |
| P12 | 2.25 | 11.26 |
| P13 | 1.10 | 5.49 |
| P14 | 1.52 | 7.60 |
| P15 | 3.00 | 15.01 |
| P16 | 0.34 | 1.68 |
| P17 | 46.63 | 233 |
| P18 | 43.96 | 220 |
| Gene | Primers | Oligonucleotide Sequence (5′→3′) | Label | Working Concentration | Amplicon Size bp | Used for | References |
|---|---|---|---|---|---|---|---|
| N | 2019-nCov_N-F | TTACAAACATTGGCCGCAAA | None | 20 μM | End-Point PCR | CDC, 2020 [10] | |
| 2019-nCov_N-R | GCGCGACATTCCGAAGAA | None | 20 μM | 66 | RT-qPCR | ||
| 2019-nCov_N-P | FAM-ACAATTTGCCCCCAGCGCTTCAG -BHQ1 | FAM, BHQ-1 | 5 μM | ddPCR | |||
| Spike | 2019-nCov_Spike-F | CGGCCTTACTGTTTTGCCAC | None | 20 μM | 75 bp | Syber Green | Falzone et al., 2020 [12] |
| 2019-nCov_Spike-R | TGTACCCGCTAACAGTGCAG | None | 20 μM | RT-qPCR | |||
| Spike/Orfa3 | 2019-nCov_Spike/Orfa3-F | TGAGCCAGTGCTCAAAGGAG | None | 20 μM | 195 | End-Point PCR | this paper |
| 2019-nCov_Spike/Orfa3-R | CGCCAACAATAAGCCATCCG | None | 20 μM | ||||
| GAPDH | hGAPDH-F | CATGAGAAGTATGACAACAGCC | None | 20 μM | 115 | RT-qPCR | Carpenter et al., 2005 [14] |
| hGAPDH-R | TGAGTCCTTCCACGATACC | None | 20 μM | ddPCR | |||
| hGAPDH-P | FAM- AGCAATGCCTCCTGCACCACCAA -BHQ1 | FAM, BHQ-1 | 5 μM |
| Sample ID | TaqMan RT-qPCR | ddPCR Absolute Quantification | TaqMan RT-qPCR | ddPCR Absolute Quantification | Spike Gene | |||
|---|---|---|---|---|---|---|---|---|
| GAPDH | GAPDH | N Gene | N Gene | SYBR RT-qPCR | Death Date | Autopsy Date | Burial Days | |
| (Ct Value) | (Copies/μL Reaction) | (Ct Value) | (Copies/μL Reaction) | (Ct Value) | ||||
| Decomposed Tissues | ||||||||
| P1 | 26.6 | 6.0 | 28.9 | 10.2 | ND | 10/05/2020 | 11/06/2020 | 32 |
| P2 | ND | 3.9 | 29.9 | 3.1 | ND | 02/05/2020 | 16/06/2020 | 45 |
| P3 | ND | 2.3 | 28.5 | 160 | 28.98 | 20/04/2020 | 17/06/2020 | 58 |
| P4 | ND | 3.0 | 30.8 | 9.6 | 29.1 | 03/05/2020 | 11/06/2020 | 39 |
| P5 | ND | 3.1 | 29 | 6.4 | 30.1 | 18/04/2020 | 18/06/2020 | 61 |
| P6 | ND | 2.8 | 29.2 | 5 | 30.9 | 15/04/2020 | 18/06/2020 | 64 |
| P7 | ND | 3.1 | 27.4 | 169 | 25.9 | 29/04/2020 | 18/06/2020 | 50 |
| P8 | 27.5 | 3.3 | 29.6 | 4.5 | 30.9 | 01/05/2020 | 16/06/2020 | 46 |
| P9 | 25.4 | 2.5 | 17.2 | 50600 | 15.5 | 22/04/2020 | 17/06/2020 | 56 |
| P10 | 26.4 | 3.0 | 24.5 | 410 | 19.4 | 24/04/2020 | 17/06/2020 | 54 |
| P11 | 28.6 | 2.4 | 28.2 | 1.1 | 24.9 | 04/05/2020 | 11/06/2020 | 38 |
| P12 | 31.6 | 2.5 | 29.8 | 1 | ND | 26/04/2020 | 16/06/2020 | 51 |
| P13 | 28.9 | 1.9 | 30.2 | 3.2 | ND | 25/04/2020 | 16/06/2020 | 52 |
| P14 | 26.6 | 3.5 | 21.3 | 1090 | 22.2 | 23/04/2020 | 17/06/2020 | 55 |
| P15 | 28.5 | 3.0 | 27.5 | 135 | 28.2 | 18/05/2020 | 11/06/2020 | 24 |
| P16 | 30.7 | 2.4 | 30.2 | 4.7 | 31.3 | 01/04/2020 | 18/06/2020 | 78 |
| Fresh Cadaveric Tissue | ||||||||
| P17 | 28.6 | 19.5 | 26.5 | 621 | 26.8 | |||
| P18 | 24.0 | 14.1 | 29.3 | 12.2 | 35.2 | |||
| Decomposed vs. Fresh Cadaveric Tissue | ||||||||
| p-value | 0.578 | 0.121 | 0.888 | 0.363 | 0.2699 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musso, N.; Falzone, L.; Stracquadanio, S.; Bongiorno, D.; Salerno, M.; Esposito, M.; Sessa, F.; Libra, M.; Stefani, S.; Pomara, C. Post-Mortem Detection of SARS-CoV-2 RNA in Long-Buried Lung Samples. Diagnostics 2021, 11, 1158. https://doi.org/10.3390/diagnostics11071158
Musso N, Falzone L, Stracquadanio S, Bongiorno D, Salerno M, Esposito M, Sessa F, Libra M, Stefani S, Pomara C. Post-Mortem Detection of SARS-CoV-2 RNA in Long-Buried Lung Samples. Diagnostics. 2021; 11(7):1158. https://doi.org/10.3390/diagnostics11071158
Chicago/Turabian StyleMusso, Nicolò, Luca Falzone, Stefano Stracquadanio, Dafne Bongiorno, Monica Salerno, Massimiliano Esposito, Francesco Sessa, Massimo Libra, Stefania Stefani, and Cristoforo Pomara. 2021. "Post-Mortem Detection of SARS-CoV-2 RNA in Long-Buried Lung Samples" Diagnostics 11, no. 7: 1158. https://doi.org/10.3390/diagnostics11071158
APA StyleMusso, N., Falzone, L., Stracquadanio, S., Bongiorno, D., Salerno, M., Esposito, M., Sessa, F., Libra, M., Stefani, S., & Pomara, C. (2021). Post-Mortem Detection of SARS-CoV-2 RNA in Long-Buried Lung Samples. Diagnostics, 11(7), 1158. https://doi.org/10.3390/diagnostics11071158