
Genetic Factors Associated with COPD Depend on the Ancestral Caucasian/Amerindian Component in the Mexican Population

 ,
,  ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Genomic DNA Extraction and Concentration Adjustment
2.3. SNPs Selection and Microarray Design
2.4. Statistical Analysis
3. Results
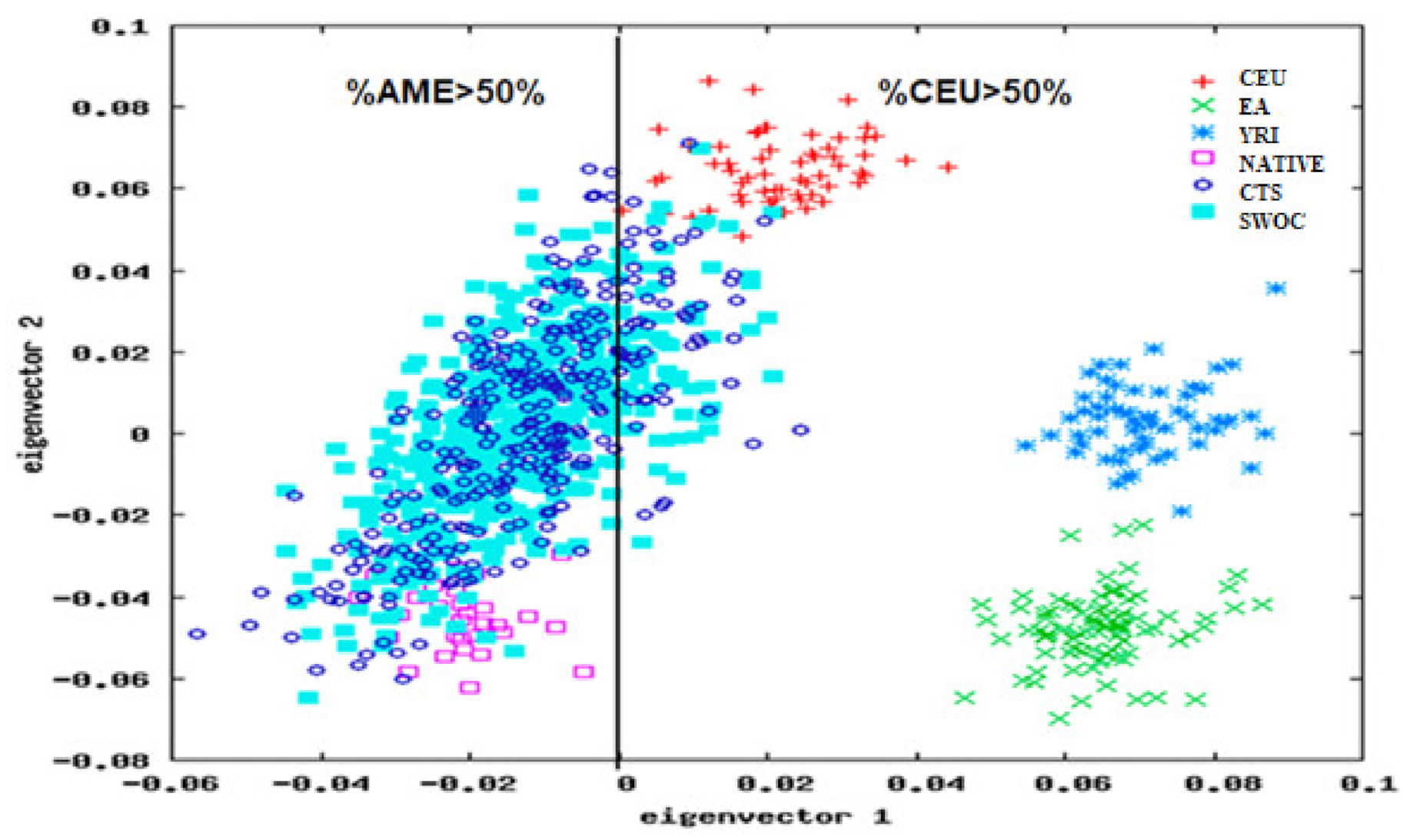
3.1. Study Population
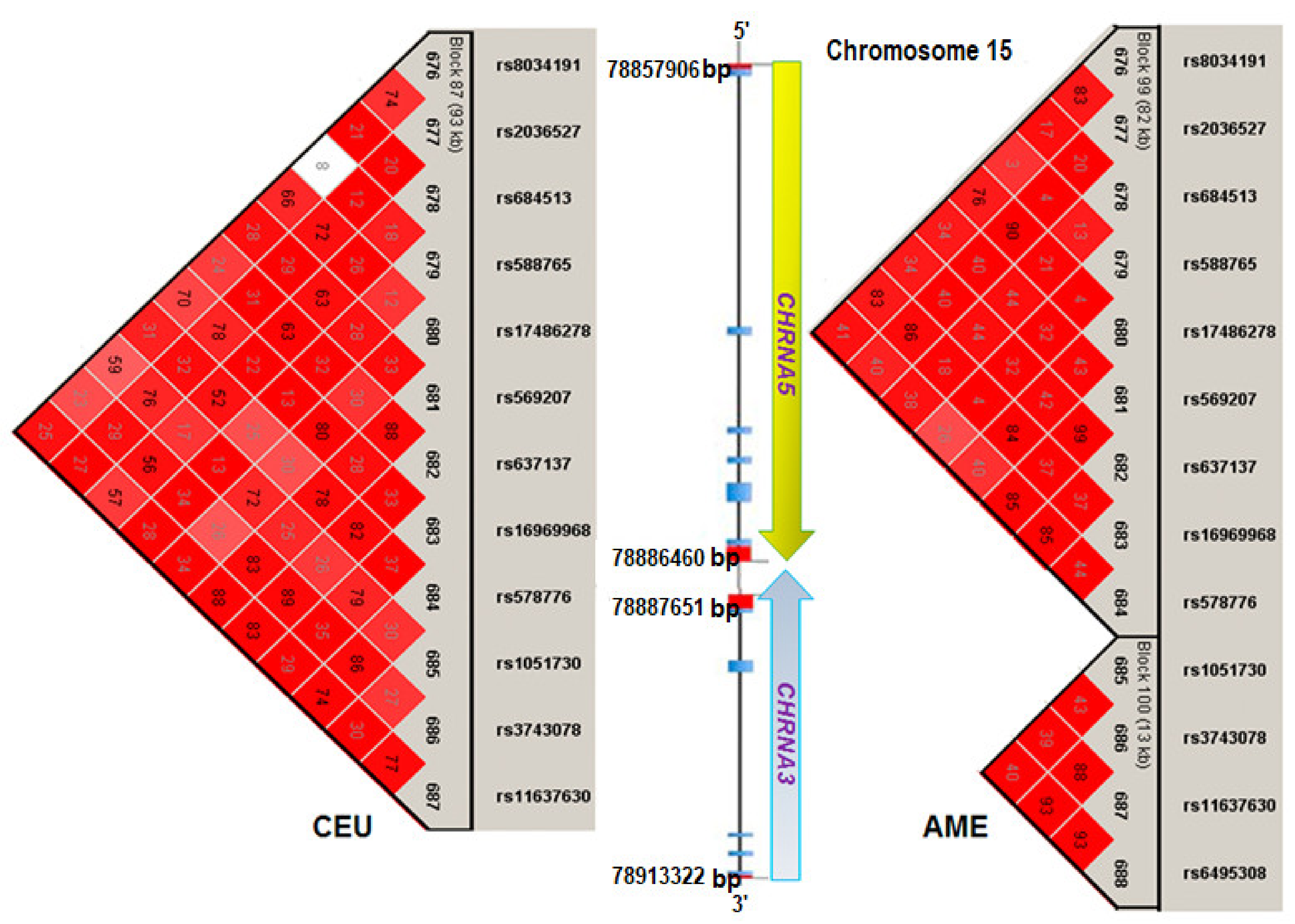
3.2. Alleles and Haplotype Analysis
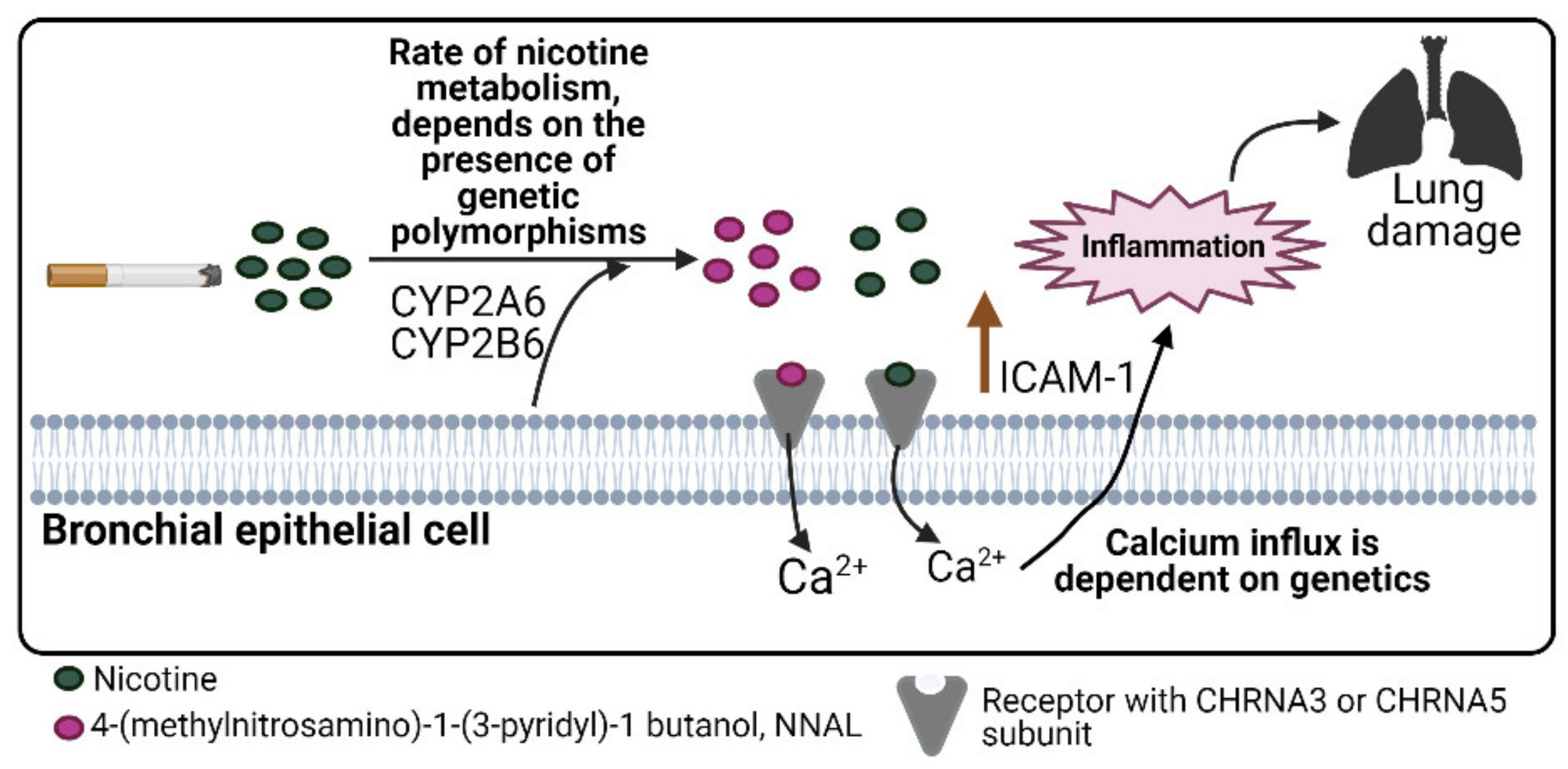
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Global Initiative for Chronic Obstructive Lung Disease. Global Strategy for the Diagnosis and Prevention of Chronic Obstructive Pulmonary Disease 2017 Report. Available online: https://goldcopd.org/wp-content/uploads/2017/02/wms-GOLD-2017-FINAL.pdf (accessed on 14 December 2020).
- Ingebrigtsen, T.; Thomsen, S.F.; Vestbo, J.; van der Sluis, S.; Kyvik, K.O.; Silverman, E.K.; Svartengren, M.; Backer, V. Genetic influences on Chronic Obstructive Pulmonary Disease-a twin study. Respir. Med. 2010, 104, 1890–1895. [Google Scholar] [CrossRef]
- Lange, P.; Celli, B.; Agustí, A.; Boje Jensen, G.; Divo, M.; Faner, R.; Guerra, S.; Marott, J.L.; Martinez, F.D.; Martinez-Camblor, P.; et al. Lung-Function Trajectories Leading to Chronic Obstructive Pulmonary Disease. N. Engl. J. Med. 2015, 373, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Brehm, J.M.; Celedón, J.C. Chronic obstructive pulmonary disease in Hispanics. Am. J. Respir. Crit. Care Med. 2008, 177, 473–478. [Google Scholar] [CrossRef] [PubMed]
- López Varela, M.V.; Montes de Oca, M. Variability in COPD: The PLATINO study viewpoint. Arch. Bronconeumol. 2012, 48, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Silva-Zolezzi, I.; Hidalgo-Miranda, A.; Estrada-Gil, J.; Fernandez-Lopez, J.C.; Uribe-Figueroa, L.; Contreras, A.; Balam-Ortiz, E.; del Bosque-Plata, L.; Velazquez-Fernandez, D.; Lara, C.; et al. Analysis of genomic diversity in Mexican Mestizo populations to develop genomic medicine in Mexico. Proc. Natl. Acad. Sci. USA 2009, 106, 8611–8616. [Google Scholar] [CrossRef]
- American Thoracic Society. Standards for the diagnosis and care of patients with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 1995, 152, S77–S121. [Google Scholar]
- Pérez-Padilla, J.R.; Regalado-Pineda, J.; Vázquez-García, J.C. Reproducibilidad de espirometrías en trabajadores mexicanos y valores de referencia internacionales [Reproducibility of spirometry in Mexican workers and international reference values]. Salud Publica Mex. 2001, 43, 113–121. [Google Scholar] [CrossRef]
- Beeckman-Wagner-LF. NIOSH Spirometry Training Guide. 2004. Available online: https://www.cdc.gov/niosh/nioshtic-2/20025502.html (accessed on 21 December 2020).
- Pérez-Rubio, G.; Silva-Zolezzi, I.; Fernández-López, J.C.; Camarena, Á.; Velázquez-Uncal, M.; Morales-Mandujano, F.; Hernández-Zenteno Rde, J.; Flores-Trujillo, F.; Sánchez-Romero, C.; Velázquez-Montero, A.; et al. Genetic Variants in IL6R and ADAM19 are Associated with COPD Severity in a Mexican Mestizo Population. COPD 2016, 13, 610–615. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef]
- Moreno-Estrada, A.; Gignoux, C.R.; Fernández-López, J.C.; Zakharia, F.; Sikora, M.; Contreras, A.V.; Acuña-Alonzo, V.; Sandoval, K.; Eng, C.; Romero-Hidalgo, S.; et al. Human genetics. The genetics of Mexico recapitulates Native American substructure and affects biomedical traits. Science 2014, 344, 1280–1285. [Google Scholar] [CrossRef]
- National Human Genome Research Institute. International HapMap Project. 2012. Available online: http://hapmap.ncbi.nlm.nih.gov (accessed on 7 July 2016).
- Patterson, N.; Price, A.L.; Reich, D. Population structure and eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Dominant markers and null alleles. Mol. Ecol. Notes 2007, 7, 574–578. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [PubMed]
- Global Initiative for Chronic Obstructive Lung Disease (GOLD). GOLD 2021. Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Pulmonary Disease, (2021 Report). Available online: https://goldcopd.org/2021-gold-reports/ (accessed on 13 February 2021).
- Shamah-Levy, T.; Vielma-Orozco, E.; Heredia-Hernández, O.; Romero-Martínez, M.; Mojica-Cuevas, J.; Cuevas-Nasu, L.; Santaella-Castell, J.A.; Rivera-Dommarco, J. Encuesta Nacional de Salud y Nutrición 2018–2019: Resultados Nacionales; Instituto Nacional de Salud Pública: Cuernavaca, México, 2020.
- Gilkes, A.; Hull, S.; Durbaba, S.; Schofield, P.; Ashworth, M.; Mathur, R.; White, P. Ethnic differences in smoking intensity and COPD risk: An observational study in primary care. NPJ Prim. Care Respir. Med. 2017, 27, 50. [Google Scholar] [CrossRef]
- Díaz-Peña, R.; Boekstegers, F.; Silva, R.S.; Jaime, S.; Hosgood Iii, H.D.; Miravitlles, M.; Agustí, À.; Lorenzo Bermejo, J.; Olloquequi, J. Amerindian Ancestry Influences Genetic Susceptibility to Chronic Obstructive Pulmonary Disease. J. Pers. Med. 2020, 10, 93. [Google Scholar] [CrossRef] [PubMed]
- Raunio, H.; Rautio, A.; Gullstén, H.; Pelkonen, O. Polymorphisms of CYP2A6 and its practical consequences. Br. J. Clin. Pharmacol. 2001, 52, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Hukkanen, J.; Jacob, P., 3rd; Benowitz, N.L. Metabolism and disposition kinetics of nicotine. Pharmacol. Rev. 2005, 57, 79–115. [Google Scholar] [CrossRef] [PubMed]
- Thorgeirsson, T.E.; Gudbjartsson, D.F.; Surakka, I.; Vink, J.M.; Amin, N.; Geller, F.; Sulem, P.; Rafnar, T.; Esko, T.; Walter, S.; et al. Sequence variants at CHRNB3-CHRNA6 and CYP2A6 affect smoking behavior. Nat. Genet. 2010, 42, 448–453. [Google Scholar] [CrossRef]
- Swan, G.E.; Lessov-Schlaggar, C.N.; Bergen, A.W.; He, Y.; Tyndale, R.F.; Benowitz, N.L. Genetic and environmental influences on the ratio of 3′hydroxycotinine to cotinine in plasma and urine. Pharm. Genom. 2009, 19, 388–398. [Google Scholar] [CrossRef]
- Styn, M.A.; Nukui, T.; Romkes, M.; Perkins, K.A.; Land, S.R.; Weissfeld, J.L. CYP2A6 genotype and smoking behavior in current smokers screened for lung cancer. Subst Use Misuse 2013, 48, 490–494. [Google Scholar] [CrossRef]
- Pezzuto, A.; Lionetto, L.; Ricci, A.; Simmaco, M.; Borro, M. Inter-individual variation in CYP2A6 activity and chronic obstructive pulmonary disease in smokers: Perspectives for an early predictive marker. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 165990. [Google Scholar] [CrossRef] [PubMed]
- Eisner, M.D.; Jacob, P., 3rd; Benowitz, N.L.; Balmes, J.; Blanc, P.D. Longer term exposure to secondhand smoke and health outcomes in COPD: Impact of urine 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanol. Nicotine Tob. Res. 2009, 11, 945–953. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, Q.; Feng, Q. CYP2B6 genetic polymorphisms influence chronic obstructive pulmonary disease susceptibility in the Hainan population. Int. J. Chron Obstruct Pulmon. Dis. 2019, 14, 2103–2115. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Tobacco Atlas 8. Cigarette Consumption. Available online: https://www.who.int/tobacco/en/atlas8.pdf (accessed on 15 February 2021).
- Perez-Padilla, R.; Menezes, A.M.B. Chronic Obstructive Pulmonary Disease in Latin America. Ann. Glob. Health. 2019, 85, 7. [Google Scholar] [CrossRef]
- Powell, R.; Barr, R.G. Response letter to: The Hispanic paradox further unraveled? Thorax 2014, 69, 185–186. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mroziewicz, M.; Tyndale, R.F. Pharmacogenetics: A tool for identifying genetic factors in drug dependence and response to treatment. Addict. Sci. Clin. Pract. 2010, 5, 17–29. [Google Scholar] [PubMed]
- Butland, B.K.; Fehily, A.M.; Elwood, P.C. Diet, lung function, and lung function decline in a cohort of 2512 middle aged men. Thorax 2000, 55, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Leng, S.; Picchi, M.A.; Tesfaigzi, Y.; Wu, G.; Gauderman, W.J.; Xu, F.; Gilliland, F.D.; Belinsky, S.A. Dietary nutrients associated with preservation of lung function in Hispanic and non-Hispanic white smokers from New Mexico. Int. J. Chron Obstruct Pulmon Dis. 2017, 12, 3171–3181. [Google Scholar] [CrossRef]
- Watson, L.; Margetts, B.; Howarth, P.; Dorward, M.; Thompson, R.; Little, P. The association between diet and chronic obstructive pulmonary disease in subjects selected from general practice. Eur. Respir. J. 2002, 20, 313–318. [Google Scholar] [CrossRef]
- Esmaillzadeh, A.; Azadbakht, L. Legume consumption is inversely associated with serum concentrations of adhesion molecules and inflammatory biomarkers among Iranian women. J. Nutr. 2012, 142, 334–339. [Google Scholar] [CrossRef]
- Hirayama, F.; Lee, A.H.; Binns, C.W.; Zhao, Y.; Hiramatsu, T.; Tanikawa, Y.; Nishimura, K.; Taniguchi, H. Soy consumption and risk of COPD and respiratory symptoms: A case-control study in Japan. Respir. Res. 2009, 10, 56. [Google Scholar] [CrossRef]
- Sim, Y.S.; Lee, J.H.; Kim, K.U.; Ra, S.W.; Park, H.Y.; Lee, C.H.; Kim, D.K.; Shin, K.C.; Lee, S.H.; Hwang, H.G.; et al. Determinants of Nicotine Dependence in Chronic Obstructive Pulmonary Disease. Tuberc. Respir. Dis. 2017, 80, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Siedlinski, M.; Tingley, D.; Lipman, P.J.; Cho, M.H.; Litonjua, A.A.; Sparrow, D.; Bakke, P.; Gulsvik, A.; Lomas, D.A.; Anderson, W. Dissecting direct and indirect genetic effects on chronic obstructive pulmonary disease (COPD) susceptibility. Hum. Genet. 2013, 132, 431–441. [Google Scholar] [CrossRef]
- Gwilt, C.R.; Donnelly, L.E.; Rogers, D.F. The non-neuronal cholinergic system in the airways: An unappreciated regulatory role in pulmonary inflammation? Pharmacol. Ther. 2007, 115, 208–222. [Google Scholar] [CrossRef] [PubMed]
- Saccone, N.L.; Culverhouse, R.C.; Schwantes-An, T.H.; Cannon, D.S.; Chen, X.; Cichon, S.; Giegling, I.; Han, S.; Han, Y.; Keskitalo-Vuokko, K.; et al. Multiple independent loci at chromosome 15q25.1 affect smoking quantity: A meta-analysis and comparison with lung cancer and COPD. PLoS Genet. 2010, 6, e1001053. [Google Scholar] [CrossRef]
- Wilk, J.B.; Shrine, N.R.; Loehr, L.R.; Zhao, J.H.; Manichaikul, A.; Lopez, L.M.; Smith, A.V.; Heckbert, S.R.; Smolonska, J.; Tang, W.; et al. Genome-wide association studies identify CHRNA5/3 and HTR4 in the development of airflow obstruction. Am. J. Respir. Crit. Care Med. 2012, 186, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Kaur-Knudsen, D.; Nordestgaard, B.G.; Bojesen, S.E. CHRNA3 genotype, nicotine dependence, lung function and disease in the general population. Eur. Respir. J. 2012, 40, 1538–1544. [Google Scholar] [CrossRef]
- Ambrechts, D.; Buysschaert, I.; Zanen, P.; Coolen, J.; Lays, N.; Cuppens, H.; Groen, H.J.; Dewever, W.; van Klaveren, R.J.; Verschakelen, J.; et al. The 15q24/25 susceptibility variant for lung cancer and chronic obstructive pulmonary disease is associated with emphysema. Am. J. Respir. Crit. Care Med. 2010, 181, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Keskitalo, K.; Broms, U.; Heliövaara, M.; Ripatti, S.; Surakka, I.; Perola, M.; Pitkäniemi, J.; Peltonen, L.; Aromaa, A.; Kaprio, J. Association of serum cotinine level with a cluster of three nicotinic acetylcholine receptor genes (CHRNA3/CHRNA5/CHRNB4) on chromosome 15. Hum. Mol. Genet. 2009, 18, 4007–4012. [Google Scholar] [CrossRef] [PubMed]
- Korytina, G.F.; Akhmadishina, L.Z.; Viktorova, E.V.; Kochetova, O.V.; Viktorova, T.V. IREB2, CHRNA5, CHRNA3, FAM13A & hedgehog interacting protein genes polymorphisms & risk of chronic obstructive pulmonary disease in Tatar population from Russia. Indian J. Med. Res. 2016, 144, 865–876. [Google Scholar]
- Pérez-Rubio, G.; Pérez-Rodríguez, M.E.; Fernández-López, J.C.; Ramírez-Venegas, A.; García-Colunga, J.; Ávila-Moreno, F.; Camarena, A.; Sansores, R.H.; Falfán-Valencia, R. SNPs in NRXN1 and CHRNA5 are associated to smoking and regulation of GABAergic and glutamatergic pathways. Pharmacogenomics 2016, 17, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Gim, J.; An, J.; Sung, J.; Silverman, E.K.; Cho, M.H.; Won, S. A Between Ethnicities Comparison of Chronic Obstructive Pulmonary Disease Genetic Risk. Front. Genet. 2020, 11, 329. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, M.J.; Strange, C.; Jones, Y.; Dickson, M.R.; Carter, C.; Moseley, M.A.; Gilbert, G.E. Does genetic testing result in behavioral health change? Changes in smoking behavior following testing for alpha-1 antitrypsin deficiency. Ann. Behav. Med. 2007, 33, 22–28. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chr | Genes | Number of SNPs |
|---|---|---|
| 1 | EPHX1 and IL6R | 24 |
| 2 | GNLY, SERPINE2, and SFTPB | 62 |
| 3 | PDZRN3 | 30 |
| 4 | GYPA and HHIP | 30 |
| 5 | ADAM19 and ADRB2 | 51 |
| 6 | GPR126 and TNF | 32 |
| 7 | HIP1, IL6, and SERPINE1 | 37 |
| 8 | CHRNB3 | 4 |
| 9 | TNFSF8 | 24 |
| 10 | SFTPD | 15 |
| 13 | FOXO1 | 13 |
| 15 | CHRNA3, CHRNA5 and CHRNB4 | 16 |
| 16 | MMP15 | 15 |
| 19 | CYP2A6, CYP2B6, EGLN2, GPATCH1, and TGFB1 | 61 |
| 20 | ADAM33 | 12 |
| 21 | ADARB1 | 26 |
| Variable | CEU (n = 118) | AME (n = 712) | ||
|---|---|---|---|---|
| CTS-CEU (n = 51) | SWOC-CEU (n = 67) | CTS-AME (n = 248) | SWOC-AME (n = 464) | |
| Age (years) 1 | 63 (50–80) | 56 (52–71) | 65 (35–84) | 65 (48–76) |
| Sex, male (%) 1,2 | 43.7 | 39.4 | 42.0 | 50.0 |
| BMI (kg/m2) 2 | 25 (17–35) | 30 (22–33) | 25 (18–36) | 30 (21–36) |
| Time smoking (years) 1,2 | 40 (10–68) | 44 (37–56) | 41 (10–62) | 45 (10–67) |
| Cpd 2 | 20 (10–80) | 40 (20–60) | 20 (10–80) | 17 (10–60) |
| Onset of smoking (years) 2 | 15 (8–30) | 15 (12–16) | 15 (7–60) | 17 (11–50) |
| Pack-years smoked 1,2 | 36 (5–170) | 74 (44–168) | 40 (6–200) | 33 (6–132) |
| FVC (%) *,1,2 | 82 (18–146) | 90 (66–92) | 86 (35–162) | 65 (20–114) |
| FEV1 (%) *, 1,2 | 53 (21–110) | 80 (65–81) | 58 (15–119) | 65 (62–120) |
| FEV1/FVC *, 1,2 | 53 (25–69) | 71 (70–75) | 54 (19–69) | 73 (70–84) |
| Population ancestry (%) | ||||
| Caucasian 2 | 64.2 | 57.0 | 36.3 | 37.0 |
| Amerindian 1 | 31.5 | 37.8 | 61.0 | 61.4 |
| African | 1.0 | 1.1 | 0.4 | 0.4 |
| East Asian | 1.5 | 1.8 | 0.6 | 0.6 |
| SNP | Gen | Risk Allele | CEU | AME | ||
|---|---|---|---|---|---|---|
| p | OR | p | OR | |||
| rs10426235 | CYP2B6 | A | 6.83E-03 | 0.221 | 9.17E-10 | 0.044 |
| rs4105144 | CYP2A6 | A | 6.43E-01 | 0.774 | 2.11E-03 | 0.485 |
| Haplotype * (CHRNA5-CHRNA3) | Haplotype Frequency (%) | p | |
|---|---|---|---|
| CTS (n = 51) | SWOC (n = 67) | ||
| AGGAAGAGGGCA | 26 | 13 | 0.007 |
| AGCGAATGAGGG | 30 | 39 | 0.174 |
| GAGGCGAAGACA | 23 | 20 | 0.667 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Rubio, G.; Falfán-Valencia, R.; Fernández-López, J.C.; Ramírez-Venegas, A.; Hernández-Zenteno, R.d.J.; Flores-Trujillo, F.; Silva-Zolezzi, I. Genetic Factors Associated with COPD Depend on the Ancestral Caucasian/Amerindian Component in the Mexican Population. Diagnostics 2021, 11, 599. https://doi.org/10.3390/diagnostics11040599
Pérez-Rubio G, Falfán-Valencia R, Fernández-López JC, Ramírez-Venegas A, Hernández-Zenteno RdJ, Flores-Trujillo F, Silva-Zolezzi I. Genetic Factors Associated with COPD Depend on the Ancestral Caucasian/Amerindian Component in the Mexican Population. Diagnostics. 2021; 11(4):599. https://doi.org/10.3390/diagnostics11040599
Chicago/Turabian StylePérez-Rubio, Gloria, Ramcés Falfán-Valencia, Juan Carlos Fernández-López, Alejandra Ramírez-Venegas, Rafael de Jesús Hernández-Zenteno, Fernando Flores-Trujillo, and Irma Silva-Zolezzi. 2021. "Genetic Factors Associated with COPD Depend on the Ancestral Caucasian/Amerindian Component in the Mexican Population" Diagnostics 11, no. 4: 599. https://doi.org/10.3390/diagnostics11040599
APA StylePérez-Rubio, G., Falfán-Valencia, R., Fernández-López, J. C., Ramírez-Venegas, A., Hernández-Zenteno, R. d. J., Flores-Trujillo, F., & Silva-Zolezzi, I. (2021). Genetic Factors Associated with COPD Depend on the Ancestral Caucasian/Amerindian Component in the Mexican Population. Diagnostics, 11(4), 599. https://doi.org/10.3390/diagnostics11040599