1. Introduction
Studies have shown that predators have both direct (density-mediated) and indirect (trait-mediated indirect) effects on prey populations (see [
1,
2,
3,
4]). In fact, it is widely accepted that ecological communities are replete with trait-mediated indirect effects arising from phenotypic plasticity, and these effects are important to community dynamics (see [
1,
3,
5]). Trait-mediated behavioral responses to predators have the potential to greatly affect the dynamics of a population (see [
1,
3,
4]). One such effect is trait-mediated emigration, wherein the prey changes its emigration patterns due to the presence of a predator [
2,
6,
7,
8]. This can, in turn, modify population dynamics and species interactions (see [
9], for example). Few empirical studies have considered this effect in the predator–prey context, where increased predation risk was shown to increase the emigration rates of prey. For example, ref. [
2] found evidence of predator-induced emigration (PIE) in a spider (predator) and planthopper (prey) system, and concluded that at high predator density, the predator had a greater impact on prey density through induced emigration than consumption (also see [
6,
7,
8]).
Human-dominated habitat fragmentation continues at unprecedented levels, giving rise to the need for a better understanding of the consequences of density dependence and its role in conservation efforts [
10,
11,
12,
13,
14]. Habitat fragmentation reduces viable habitat or patch size and also separates populations among much smaller residual patches which are surrounded by a human-modified “matrix” with varying degrees of hostility [
11]. The modeling of theoretical populations has seen great success in predicting patch- and even landscape-level patterns in response to habitat fragmentation. The reaction diffusion framework has been particularly successful at providing better understanding of the coupling of density dependent growth mechanisms with density dependent movement or dispersal (see [
15]). The framework’s ability to handle space explicitly at the landscape level, including modeling animal movement behavior differences when a patch boundary is reached, has been well demonstrated [
16,
17,
18,
19].
The modeling of predator–prey population dynamics dates back to the classic works of [
20,
21]. A key component of those models is the predator functional response, the relationship between prey consumption per predator per unit of time as prey density increases [
22,
23]. In the original formulation of the Lotka–Volterra model, predators and prey were assumed to encounter each other at random, resulting in a constant rate of prey consumption as the prey density increased. Subsequent derivations of the functional response have included more realistic predator behaviors such as handling time constraints and predator satiation that result in a decreasing rate of prey consumption as the prey density increases (Holling’s type II functional response), or learning or switching to more abundant or profitable prey that results in an increasing, then decreasing, rate of prey consumption as the prey density increases (Holling’s type III functional response) (see [
22,
24,
25]). Over the years, many studies have expanded on Holling’s seminal work and investigated the biological implications of a wide range of predator functional responses (e.g., see [
26,
27,
28,
29,
30]).
In this paper, we employ a model based on the reaction diffusion framework to study the persistence of a prey species that is experiencing both direct (density-mediated) and indirect (predator-induced emigration) effects while facing habitat fragmentation. This framework was derived in [
19] (but also see [
31]) and connects assumptions regarding movement behavior at the individual level to the patch- and landscape levels. To our knowledge, no other study has examined the linkage between predator presence, the prey’s density–emigration relationship and prey population dynamics in a landscape context. Here, we envision a prey species which inhabits a patch
which is a bounded domain in
;
with smooth boundary
or
that is surrounded by a hostile matrix. In the framework, prey grow logistically and move according to an unbiased random walk inside the patch and in the matrix but follow a biased random walk at the patch/matrix interface, making an emigration decision based upon the presence of a predator. We assume that the predator is acting on a different timescale than the prey and thus assume a constant predator population. The nondimensionalized steady-state equation for the model is then given by
where
is the prey population density for
,
is a composite parameter that is proportional to the patch size squared,
is a parameter that quantifies the matrix hostility, and
is the outward normal derivative of
u. Here,
, where
is the probability of the organism staying inside
upon reaching the boundary and represents emigration patterns such as density-independent emigration (DIE) when
is a constant, positive density-dependent emigration (+DDE) when
is increasing, and negative density-dependent emigration (−DDE) where
is decreasing.
Here, represents the predation functional response, and we consider the following forms:
- (i)
Constant-yield predation (CYP): .
- (ii)
Constant-effort predation: Holling Type I functional response: .
- (iii)
Prey switching predation: Holling Type III functional response: ; .
For the emigration form g, we consider the following:
- (i)
Density-independent emigration (DIE):
- (ii)
Positive density-dependent emigration (+DDE):
- (iii)
Negative density-dependent emigration (−DDE):
Where
is a composite parameter that measures the prey’s sensitivity to the presence of the predator (we will also denote this as the strength of the PIE relationship) and
is a composite parameter that dictates the strength of the DDE form (see [
32]). For example, a −DDE form with
is very similar to DIE, while one with
shows a strong negative relationship between density and emigration.
Recently, several studies have either considered models similar to (
1) but with only direct effects of predation or indirect effects such as PIE (see [
29,
33,
34,
35,
36,
37,
38]).
To date, few, if any, studies have considered the combined effects of both the direct and indirect effects of the predation of a prey in the presence of habitat fragmentation. Our motivation here is to compare the structure of the positive solutions for (
1) when predation does not affect the emigration rate (
) to the case when predation does influence the emigration rate (
). We establish several existence, non-existence, and multiplicity results via the method of sub- and supersolutions and then numerically explore the structure of positive steady-state solutions in the one-dimensional case where more complete results and biological interpretation can be obtained.
To present our main results, we first describe a useful eigenvalue problem. Given
and
, let
be the principal eigenvalue of
with corresponding eigenfunction
. Further, let
for
, and
be the positive falling zero of
. Throughout this paper, to ensure the existence of positive solutions for (
1), we assume that
is a fixed number small enough so that this
exists and
, where
. Now, we state our main results.
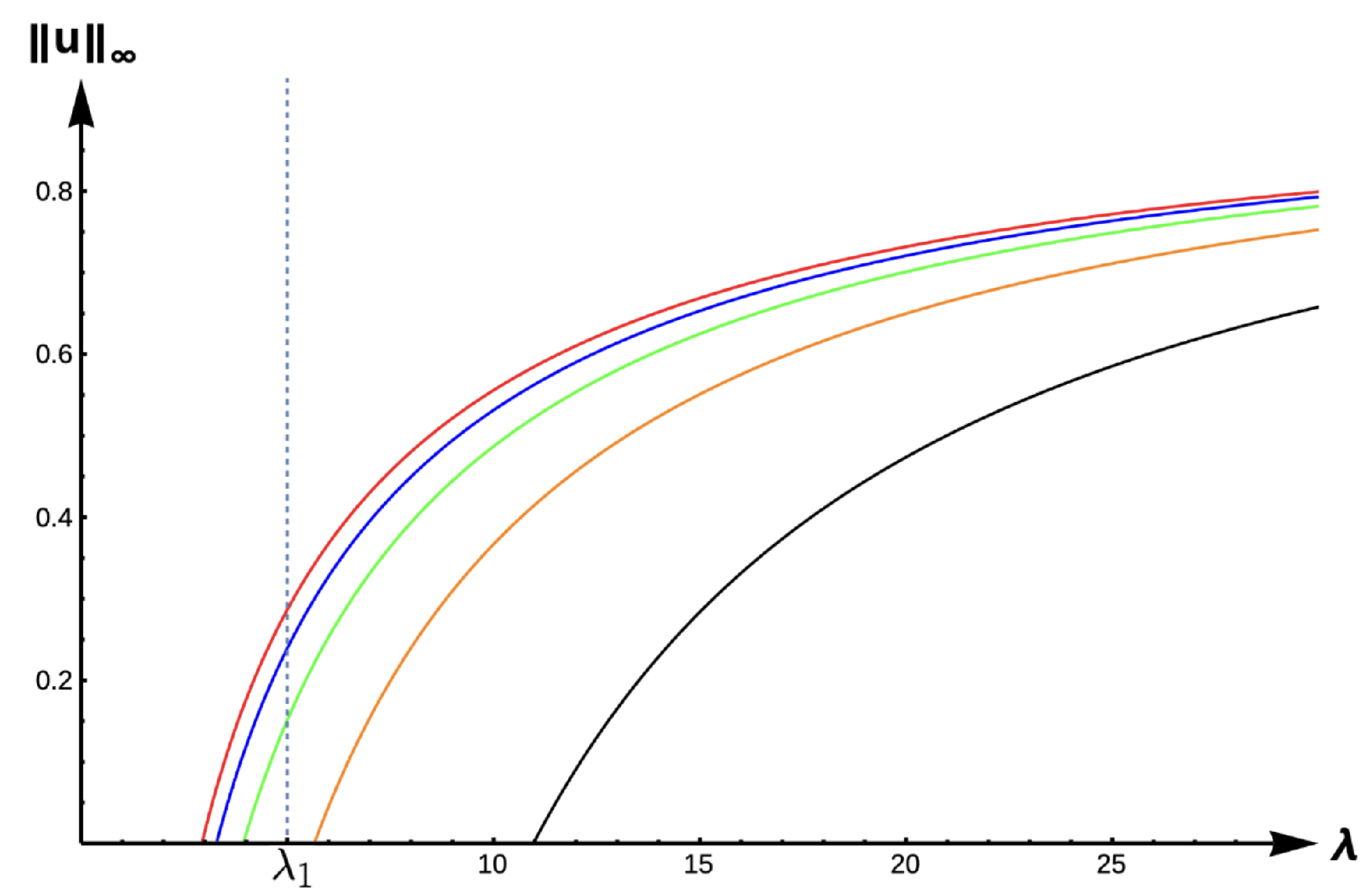
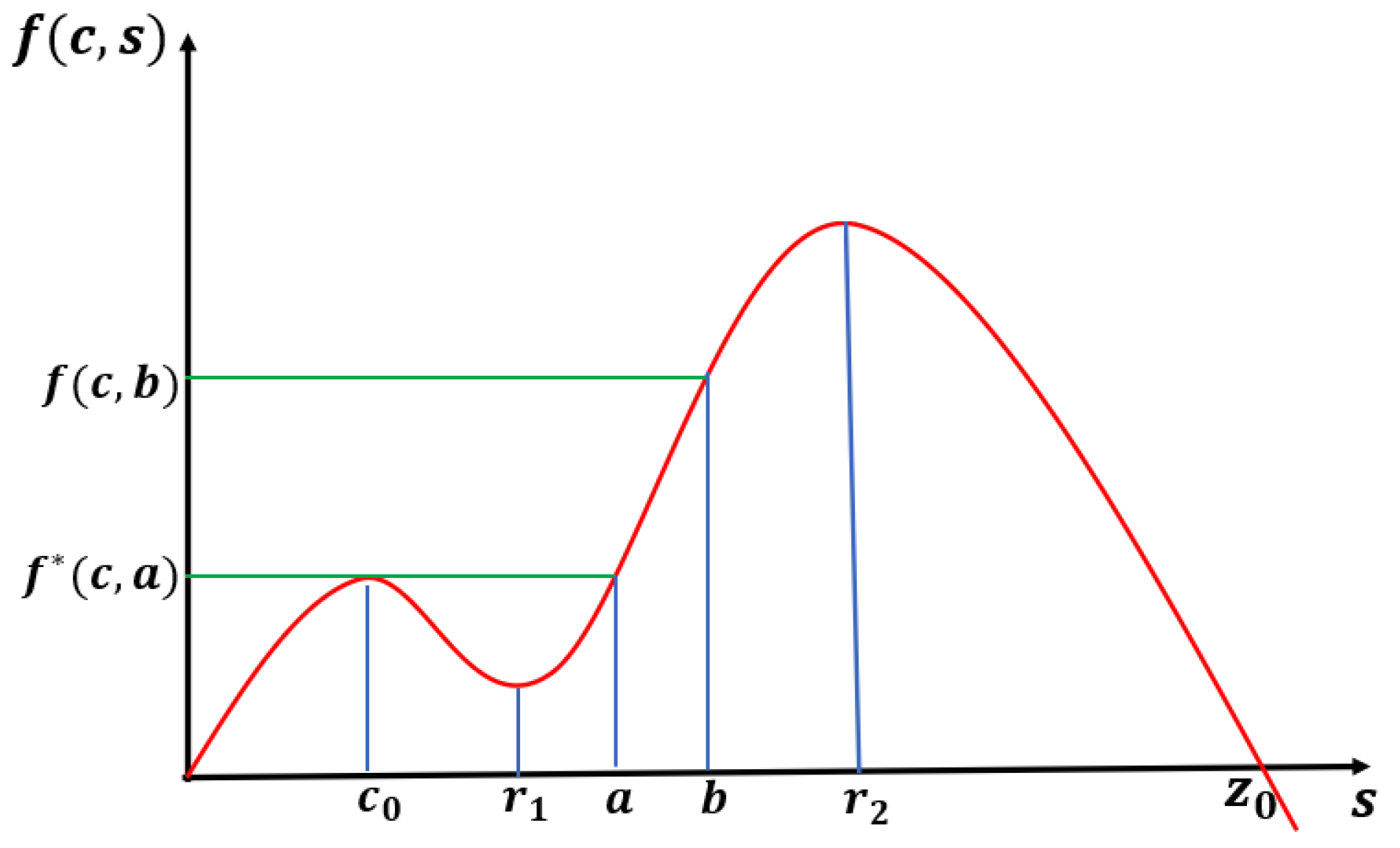
Notice that for
,
has two distinct zeros,
with
. We now state the results for this case with an expected bifurcation curve presented in
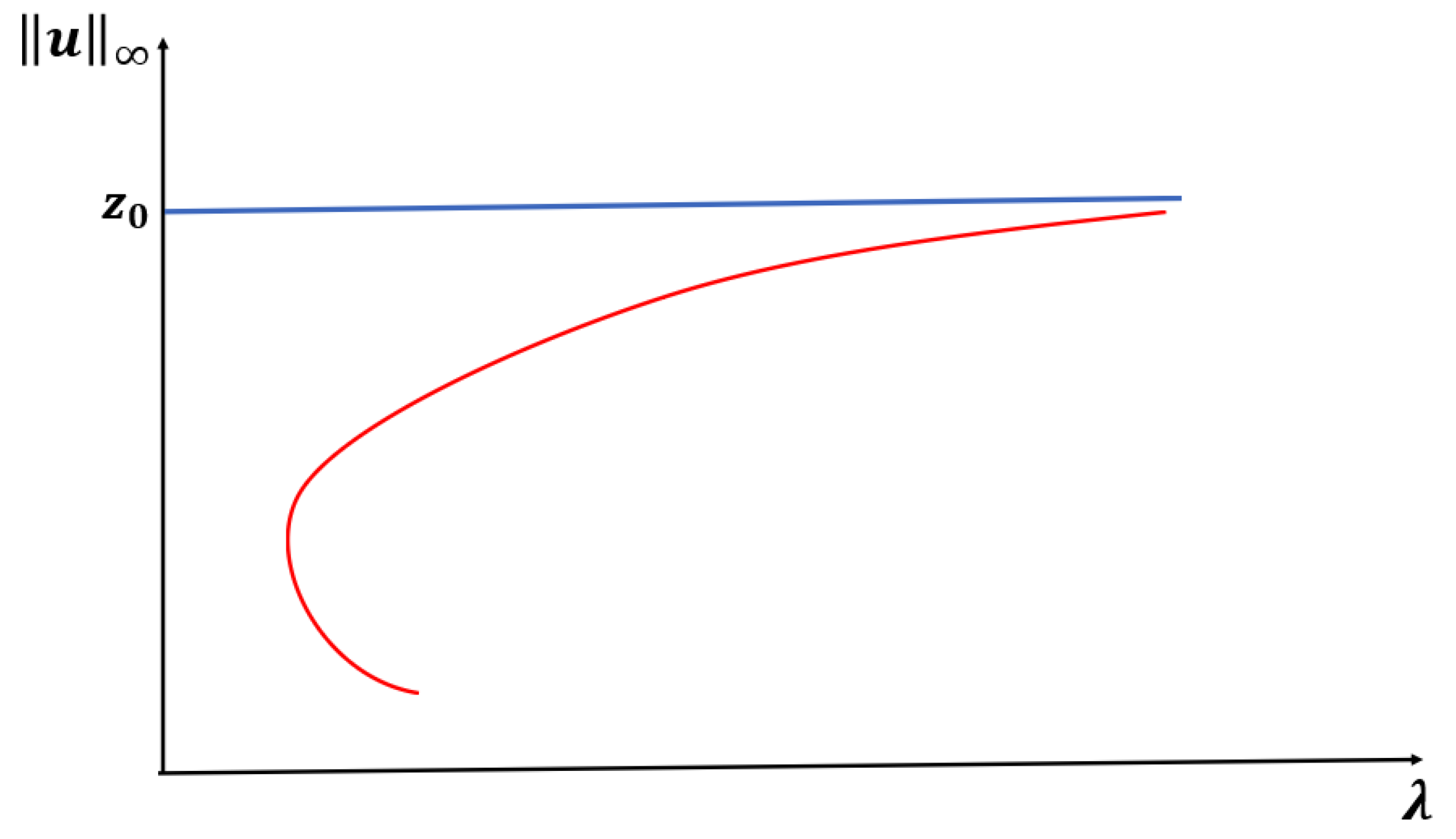
Figure 1.
Theorem 1. Let or and be fixed. Then, (1) has at least one positive solution for such that as (see Figure 1). Moreover, the time-dependent model predicts extinction for any when the initial density profile is below . For the next two cases (
and
), we also focus on exploring the existence of a patch-level Allee effect (PAE). In this scenario, the trivial solution and at least one other positive solution of (
1) are both stable, giving rise to a population density threshold that must be maintained to ensure the persistence of the population. A sufficient condition for a PAE is the existence of a range of
to the left of
where at least one positive solution exists (see
Figure 2 and [
32]). Here,
when
and
when
. Furthermore, when
, numerical evidence shows that there is also a range of
with multiple positive solutions. However, this is a well-known open problem to establish analytically (see the literature on semi-positone problems).
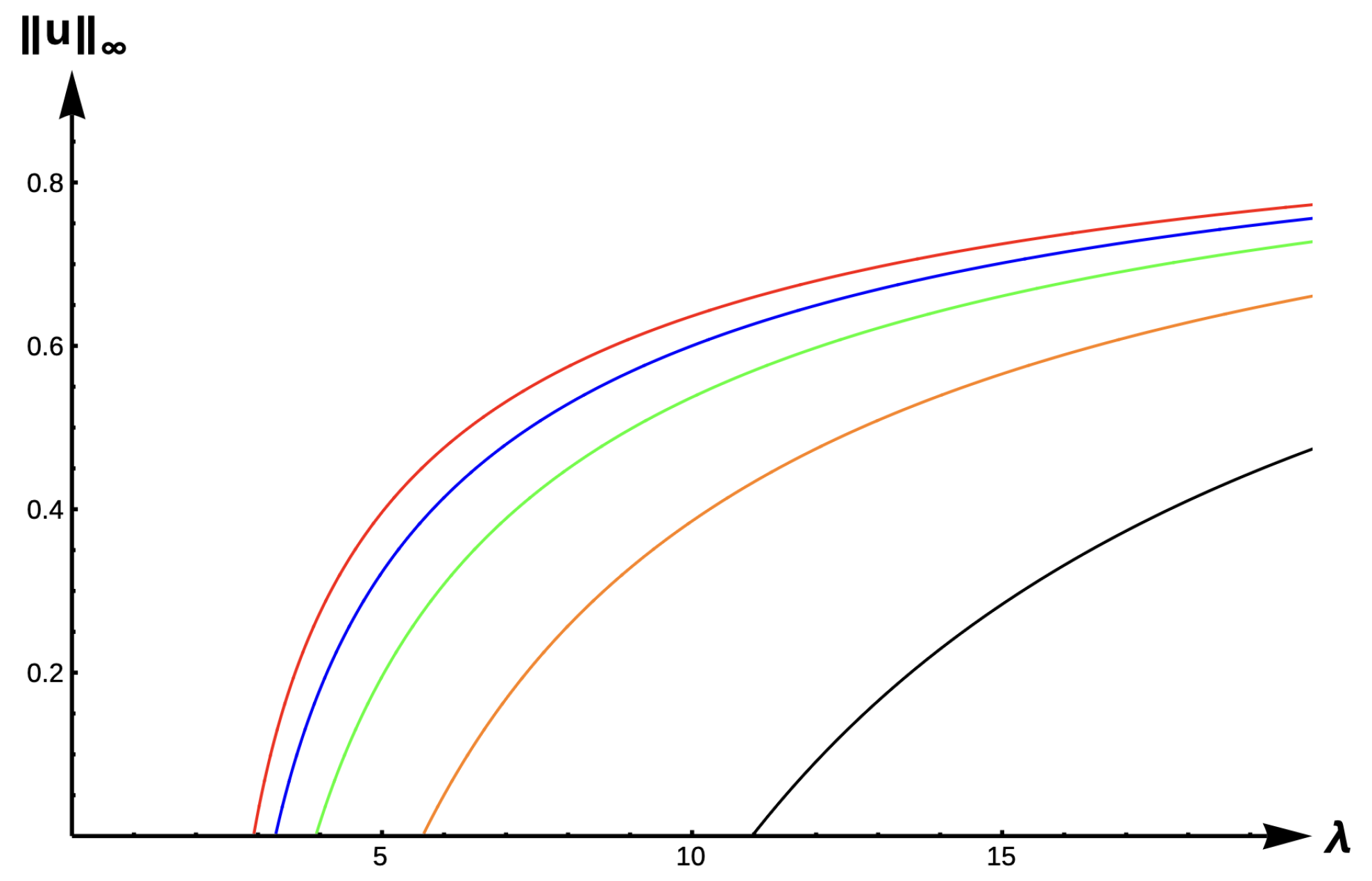
Results for Case B: Constant-Effort Predation:
In this case, the roots of are zero and . We now state the results for this case.
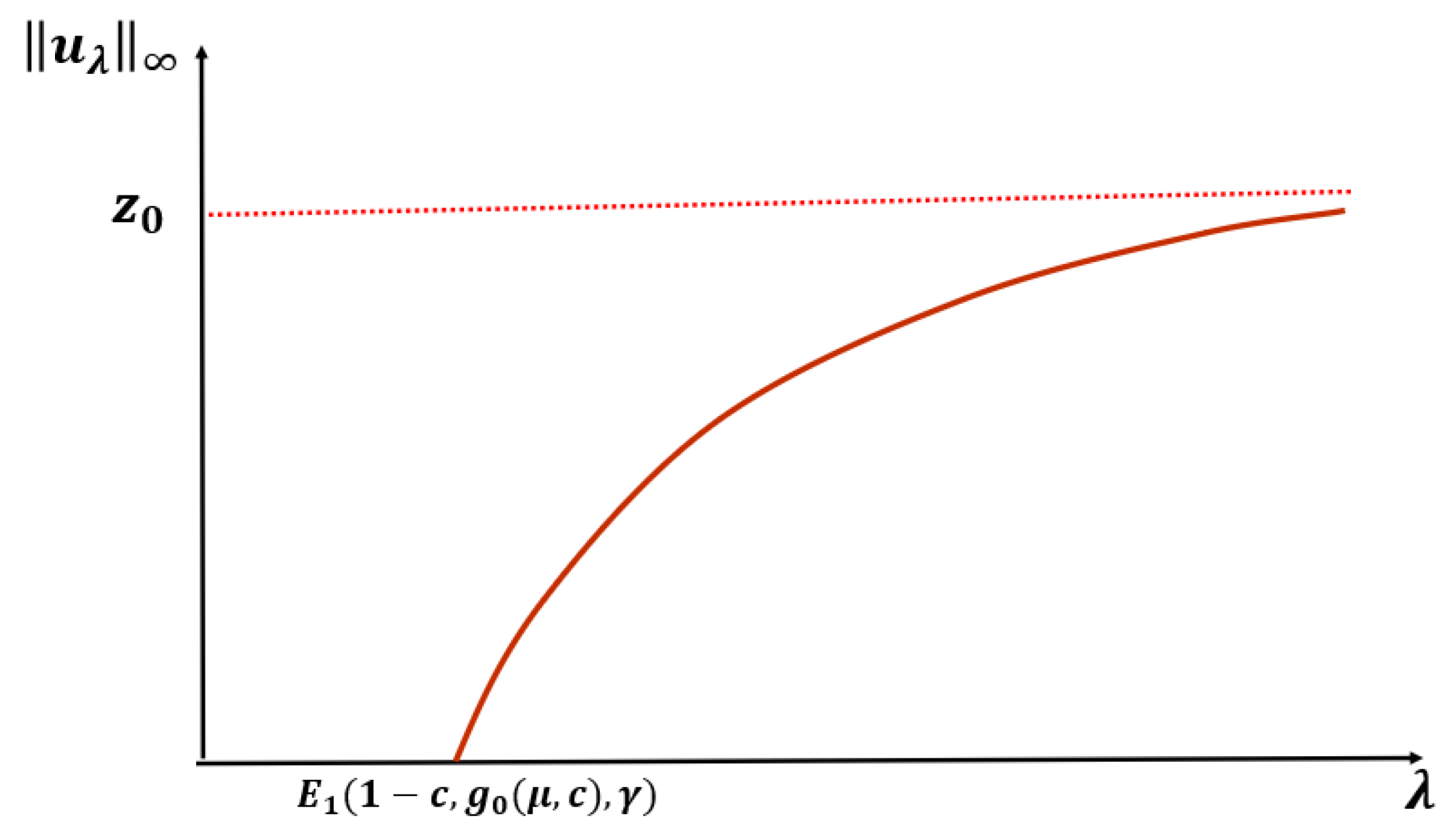
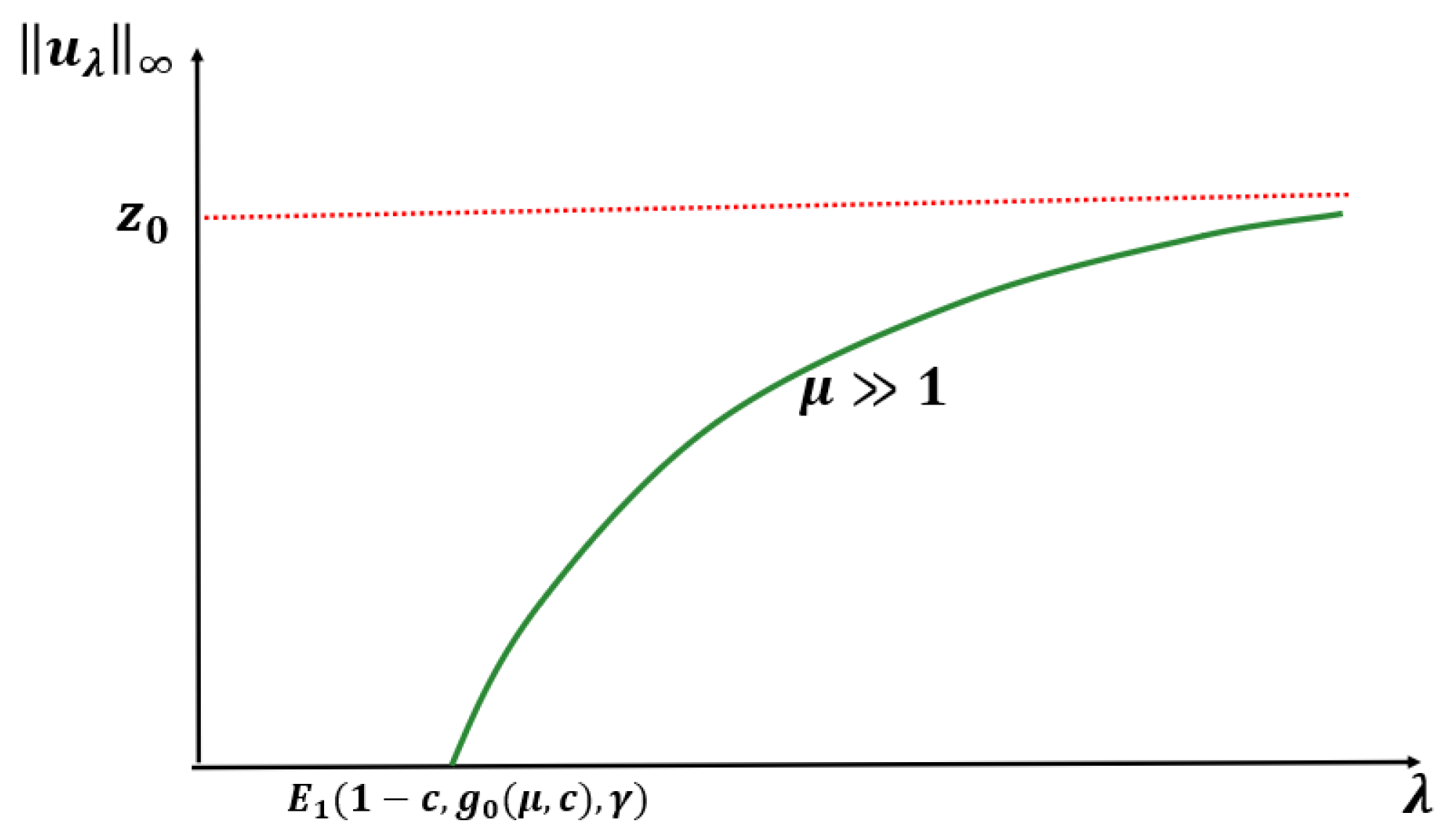
Theorem 2. Let or and be fixed. Then, (1) has a unique positive solution for and no positive solution for . Further, as and as (see Figure 3). Remark 1. Let or , , , and be the unique positive solutions of (1) when and , respectively, then (see Figure 4). Before we state our next theorem, we define
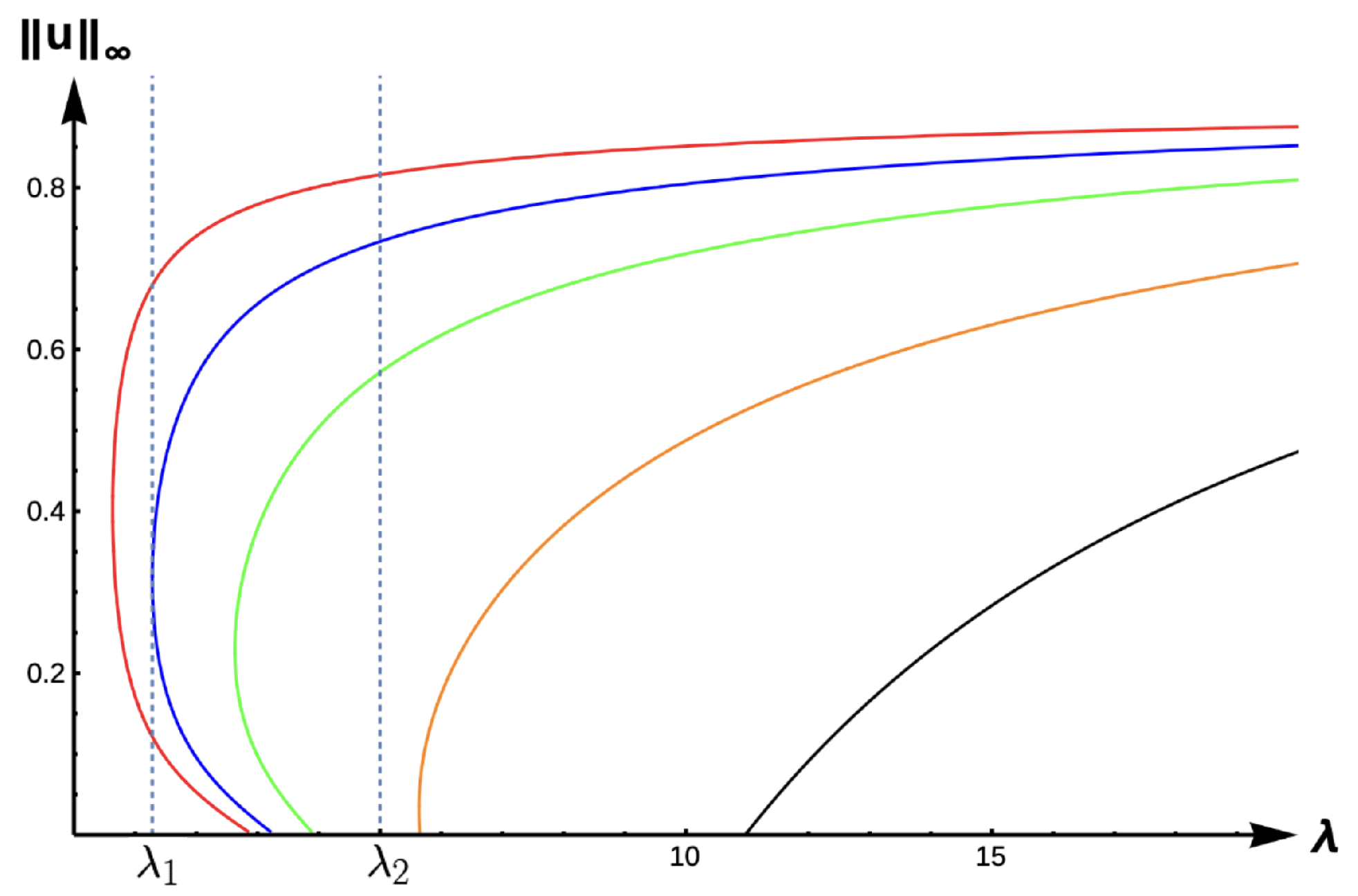
Theorem 3. Let . Then, for fixed and , there exists such that a PAE occurs for for . Further, if and are fixed, then there exists such that (1) has no positive solution for when , where is the principal eigenvalue of(see Figure 5). Next, we state the following conjecture based on our numerical results (see
Section 2).
Conjecture: For any
and fixed
, when
, (
1) has no positive solution for
, and (
1) has a unique positive solution
for
such that
as
and
as
(see
Figure 6).
Here, we state two hypotheses before we state our results. Let
be the radius of the largest ball that can be inscribed inside the domain
,
,
, and given a
, denote
as the unique solution of
with
and
.
Now, we state two hypotheses regarding f:
- (H1):
there exist such that and .
- (H2):
there exist and such that f is non-decreasing in .
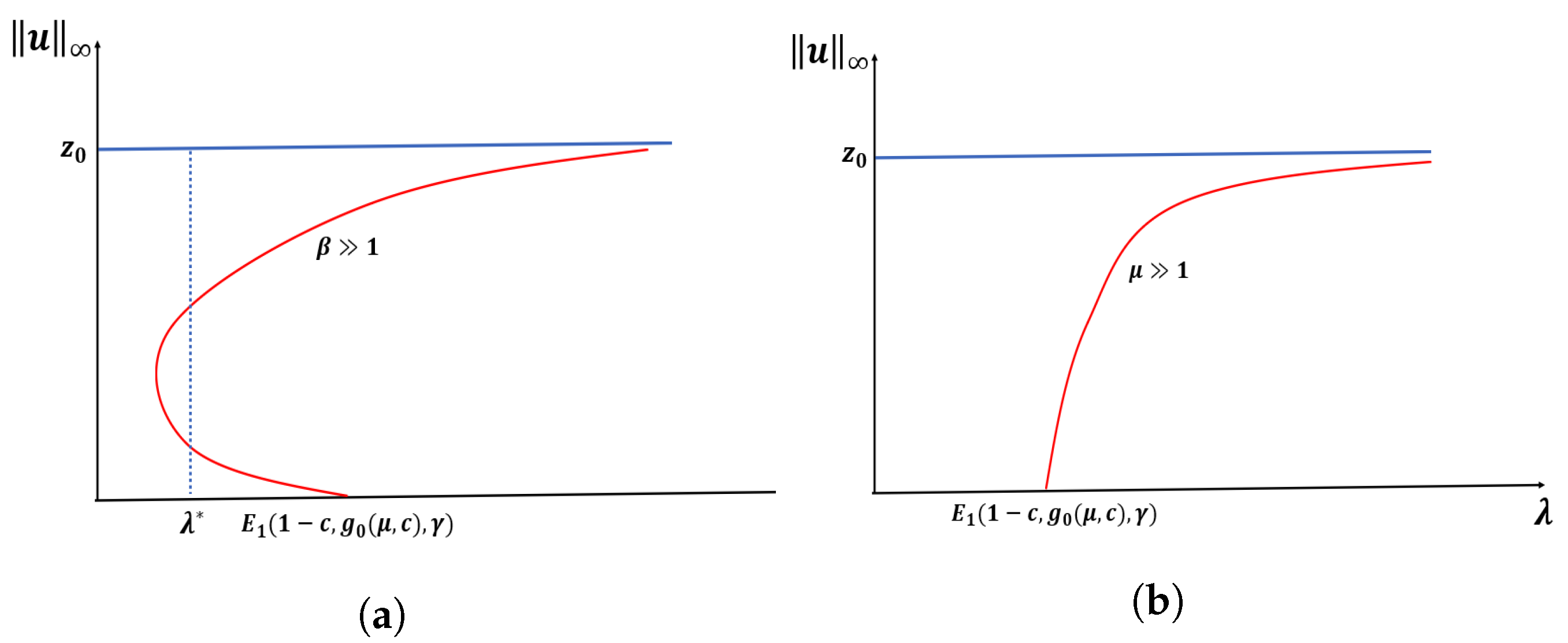
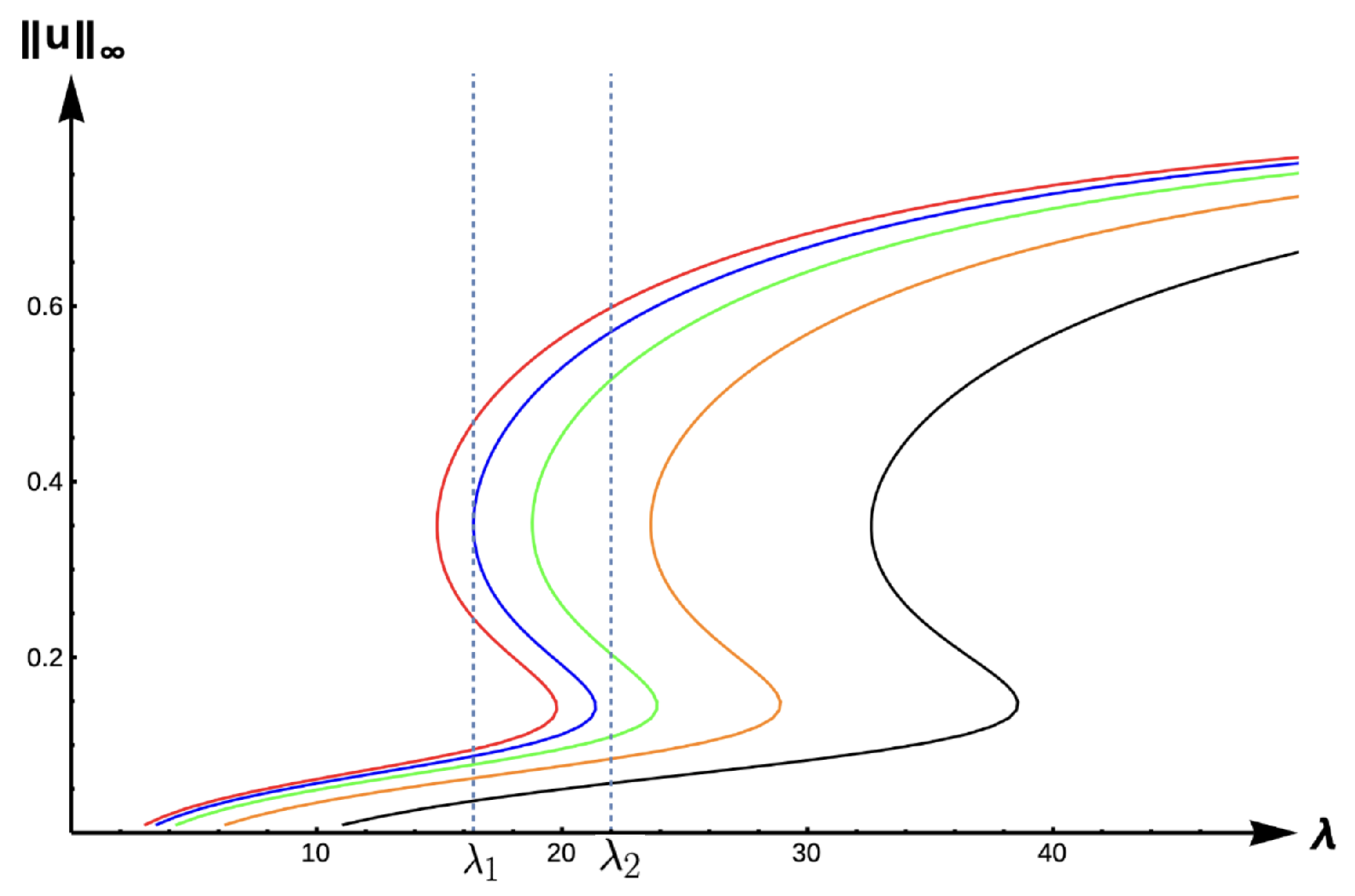
Theorem 4. Let . Then, for and fixed , there exists such that a PAE occurs for for . Further, if and are fixed, there exists such that (1) has no positive solution for when . Theorem 5. Let , , or with β be fixed. Let and hold. Then (1) has at least three positive solutions for (see Figure 7). Remark 2. In Section 4, when Ω is a ball of radius R, we prove that f and g satisfy the hypothesis of Theorem 5 for certain parameter values when μ is large. Theorem 6. Let or . Then, (1) has no positive solution for (see Figure 7). Remark 3. In Case A, (1) has no positive solutions for and one can easily adapt the techniques used in the literature of semi-positone problems with Dirichlet boundary conditions to prove this. For Cases B and C, we will later establish that (1) has no positive solutions for . In
Section 2, we numerically compute complete bifurcation diagrams of the positive solutions in the one-dimensional case where
and discuss their biological relevance. We provide some mathematical preliminaries in
Section 3, and in
Section 4 we construct several sub- and supersolutions that we will use to establish our analytical results. Finally, we prove Theorems 1–6 and Remark 1 in
Section 5.
2. One-Dimensional Results and Biological Conclusions
In this section, we present computationally generated results for the one-dimensional case when
. In this way, we obtain more detailed bifurcation diagrams and provide some biological conclusions of these results. Namely, we consider the steady-state model:
and study the structure of positive solutions via bifurcation curves (
vs.
curves) in order to contrast model predictions of the cases:
(when predation does not affect emigration probability) and
(when predation does influence the emigration probability). Here, we denote
as the maximum density of the positive solution of (
7). We obtain complete bifurcation diagrams via a quadrature method (presented in Lemma 3 of
Section 3 and numerical computation using Mathematica (Wolfram Research Inc., version 14.0). See [
32,
39], and [
40] for extensions of the original quadrature method developed in [
41] for the case of Dirichlet boundary conditions. For completeness, we prove the extension of the quadrature method employed here in
Appendix A. For simplicity of presentation, we choose
for
and
, and
and
for
. We note that a full exposition of the parameter space is outside the scope of this current work. Rather, we choose certain parameter ranges to provide prototypical model predictions. We also highlight several cases where predator-induced emigration and the strength of this response as measured in
plays a significant role on model predictions.
2.1. Results for Case A: Constant-Yield Predation
In this subsection, we present results for the constant-yield predation case, where we have fixed and .
We note that when
, our model has a semi-positone structure (see [
29]), and for Lemma 3 to hold, the
-value must be between
and
, where
is the first positive zero of
F and
is the falling positive zero of
f (see
Figure 8).
Before we state our results, we denote the -value associated with the minimum patch size as , namely, is proportional to the minimum size of the habitat that will allow unconditional population persistence.
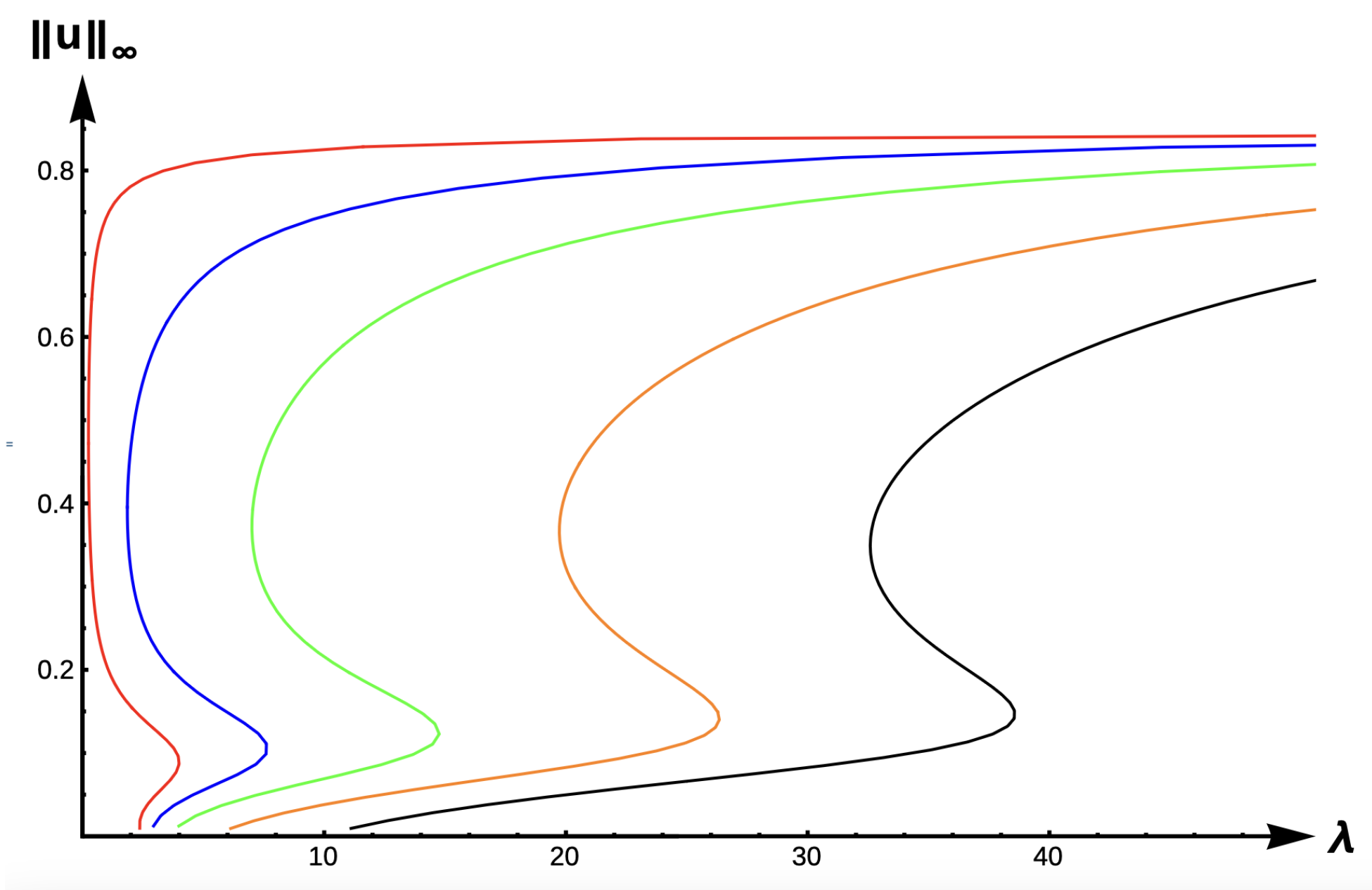
2.1.1. Bifurcation Diagrams for DIE:
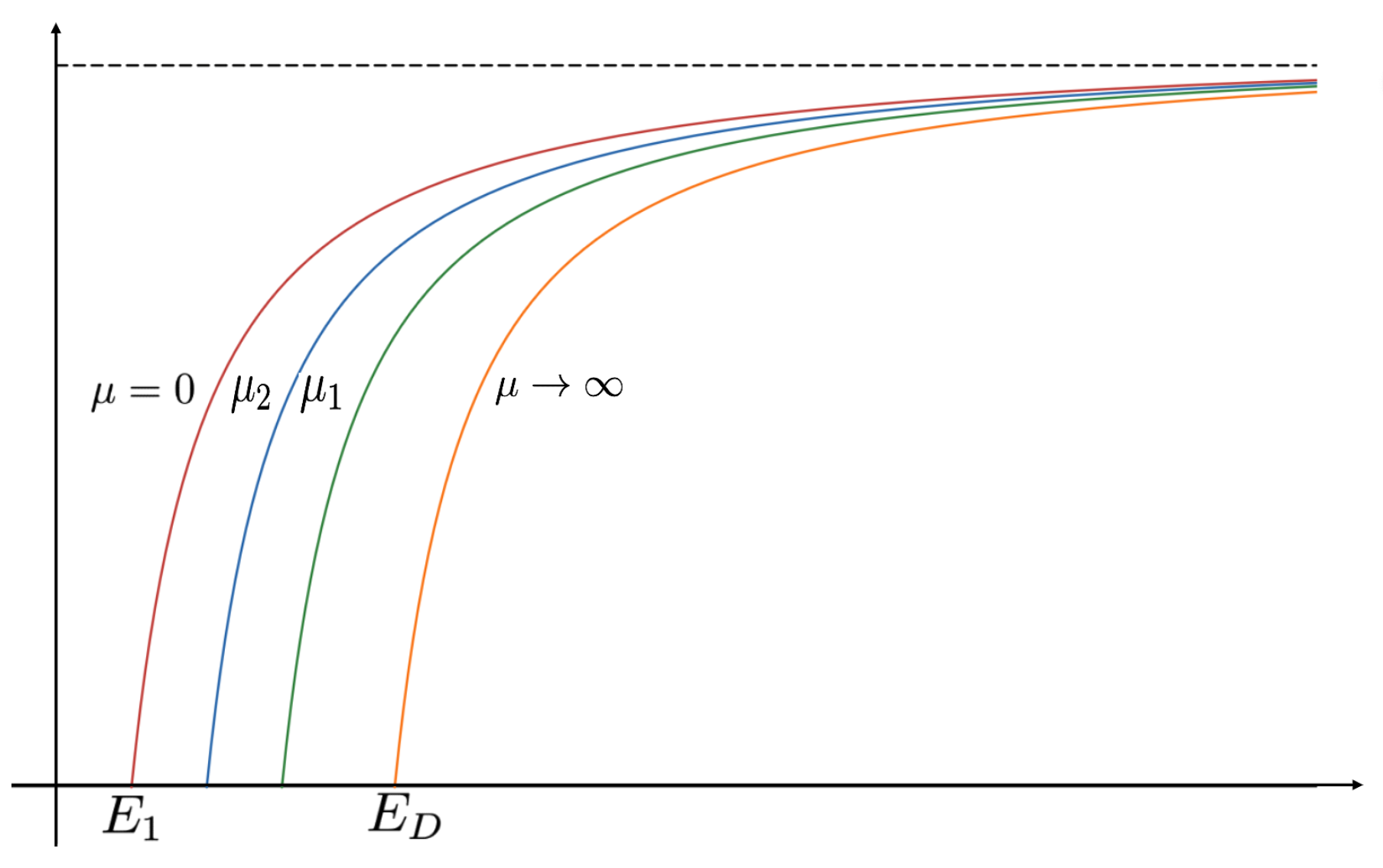
The evolution of bifurcation curves with respect to PIE strength
is shown in
Figure 9, while
Table 1 shows the evolution of
, the uniqueness region, and the multiplicity region. Notice that for all
, the smaller root of
is a supersolution of (
7), implying that the time-dependent problem would predict extinction for any size patch if the initial density distribution is too small. Here,
corresponds to the case where predation does not influence the emigration probability. We observe that the bifurcation curves shift from left to the right, the multiplicity region increases, and the uniqueness region decreases as
increases. The bifurcation curves are approaching the bifurcation curve corresponding to a case of a completely lethal matrix (Dirichlet boundary condition) as the PIE strength
. For a given
-value, we see that the number of positive steady states also varies as
changes. To highlight changes in model predictions with respect to the PIE strength, consider a patch with fixed matrix hostility and patch size yielding a
as shown in
Figure 9. In the absence of PIE (
), there is a unique positive solution of (
7),
and
show two positive solutions, and no positive solution is possible (extinction) when
for some
. Notice also that the red curve (
) and the black curve (
) define an envelope of possible bifurcation curves, where PIE would have an effect on the model predictions of the population’s persistence. In other words, only populations with patch sizes having corresponding
-values lying in this envelope could be adversely affected by the presence of PIE.
2.1.2. Bifurcation Diagrams for +DDE
In the case of +DDE with strength measured in the nondimensional parameter
, the bifurcation diagram given in
Figure 10 is very similar to that of the DIE case (
Figure 9), albeit shifted further to the right. The
, uniqueness region, and multiplicity region are given in
Table 2 for
, and in
Table 3 for
(a stronger +DDE response). Model predictions are identical for this case as the previous one, including our ability to identify
-values where the number of positive solutions changes from one to two, and then to none, as
increases in the respective envelope.
2.1.3. Bifurcation Diagrams for −DDE:
Figure 11 shows the evolution of bifurcation curves with respect to
, the −DDE case with
. Again, the bifurcation diagram is very similar to that of the DIE case (
Figure 9), albeit shifted further to the left. The
, uniqueness region, and multiplicity region are given in
Table 4 for
, and in
Table 5 for
(a stronger +DDE response). Model predictions are identical for this case as the previous one, including our ability to identify
-values where the number of positive solutions changes from one to two, and then to none, as
increases in the respective envelope.
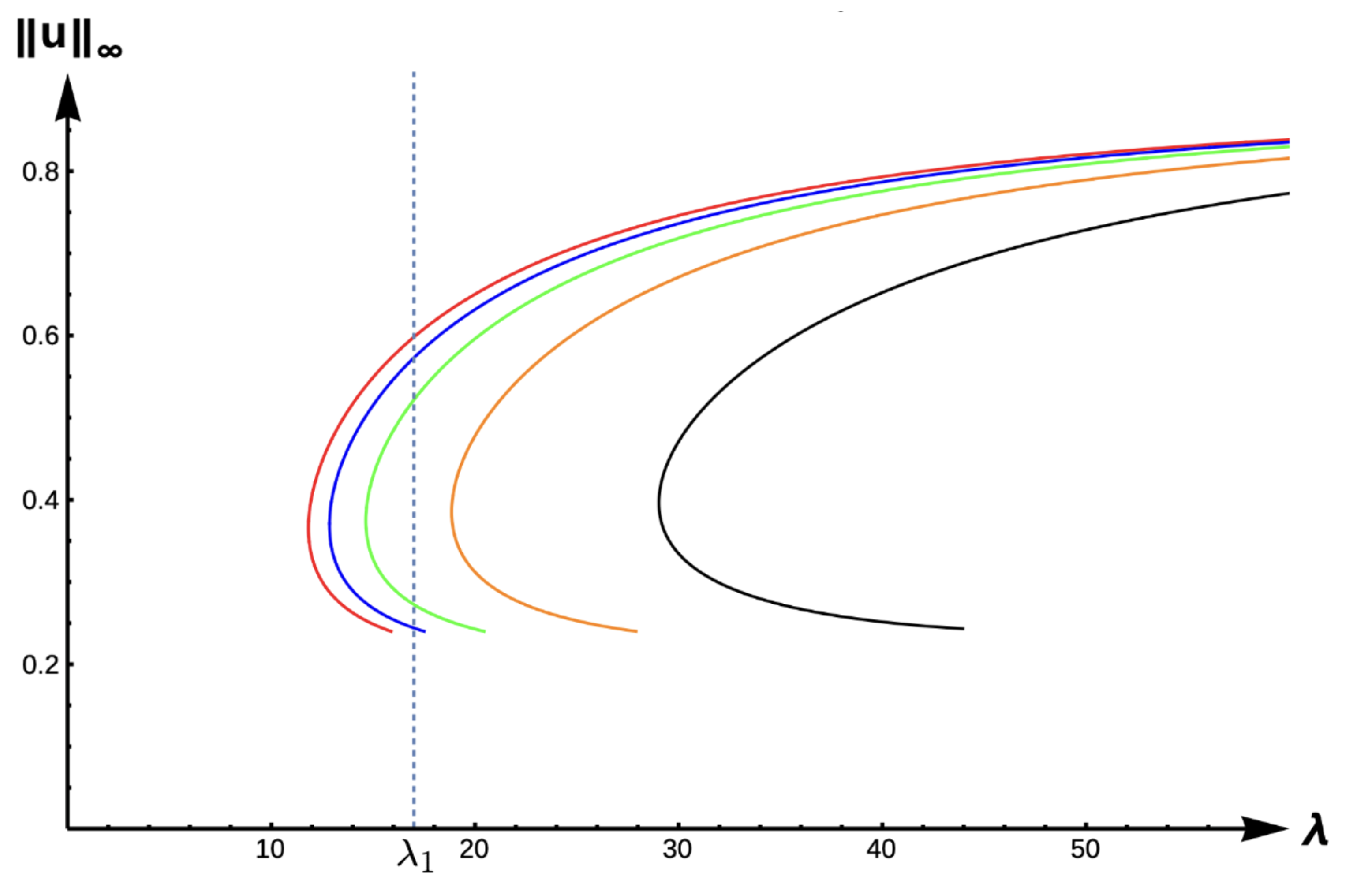
2.2. Results for Case B: Constant-Effort Predation:
In this subsection, we present results for the constant-effort predation case, where we have fixed
and
. We note that
has two zeros (0 and 0.9) when
. Here, Lemma 3 holds for
and the shapes of
and
are as in
Figure 12.
2.2.1. Bifurcation Diagrams for DIE:
For the DIE case,
Figure 13 displays the bifurcation diagram of the positive solutions for (
7), and
Table 6 gives the corresponding
-values for various
-values. We observe that the solution to (
7) is always unique with a prediction of extinction for
and unconditional persistence for
. The bifurcations curve translate to the right, and hence
-values increase as
increases (see
Table 6). The bifurcation curves are again approaching the bifurcation curve corresponding to the completely lethal matrix case (Dirichlet boundary condition) as
. We again highlight that model predictions are drastically affected by PIE strength as measured in
for patch sizes with a corresponding
-value inside the envelope determined by the red (
) and black (
) curves. For example, for a patch size with corresponding
,
Figure 13 shows model predictions of unconditional persistence when
, and extinction when
for some
.
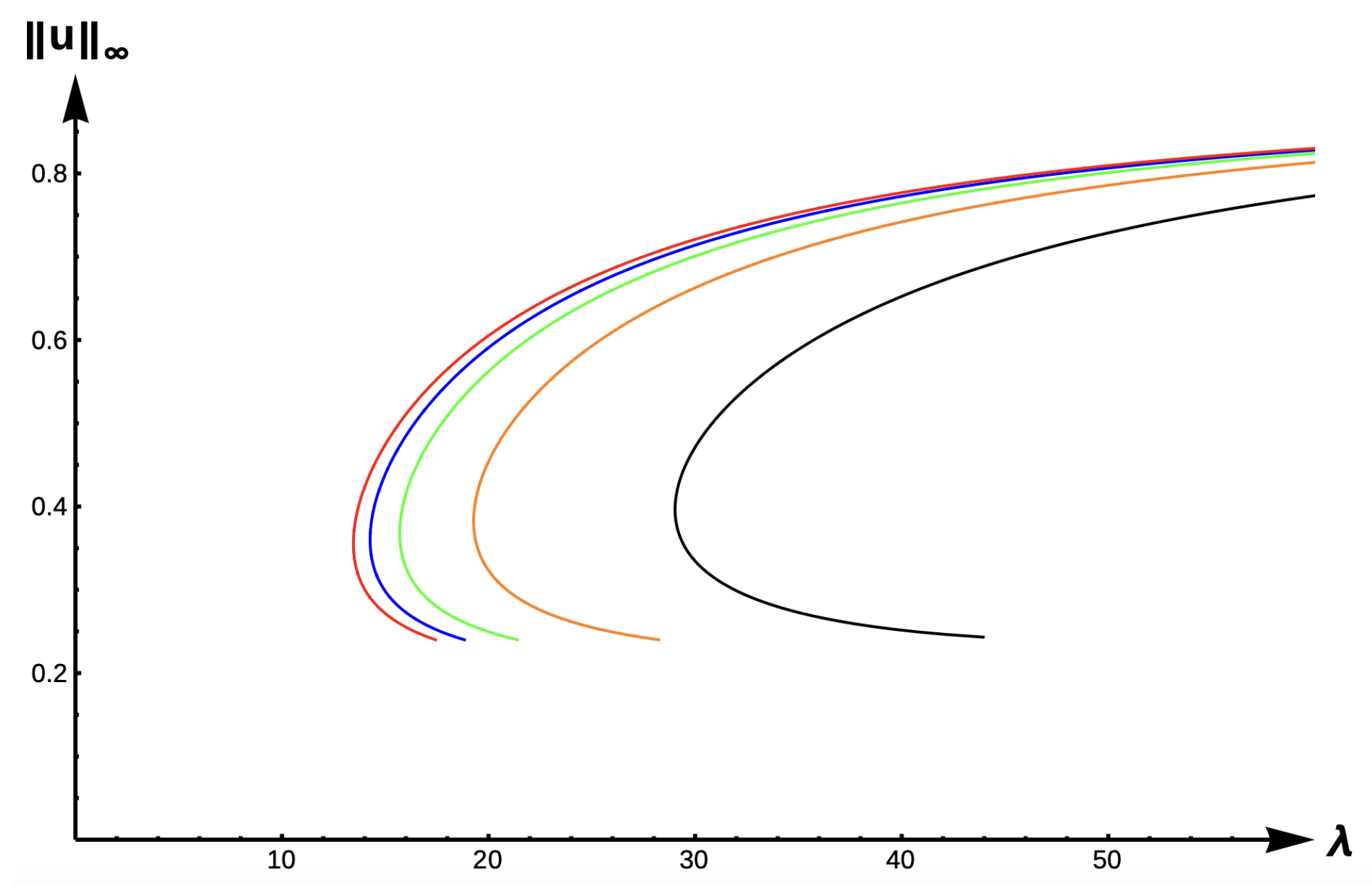
2.2.2. Bifurcation Diagrams for +DDE:
Figure 14 shows evolution of the bifurcation curves, and
Table 7 gives the corresponding
-values for the +DDE case with various
values. The bifurcation curves translate to the right and approach the bifurcation curve corresponding to the completely lethal matrix case (Dirichlet boundary condition) as
increases to
∞. Model predictions are identical to that of the DIE case.
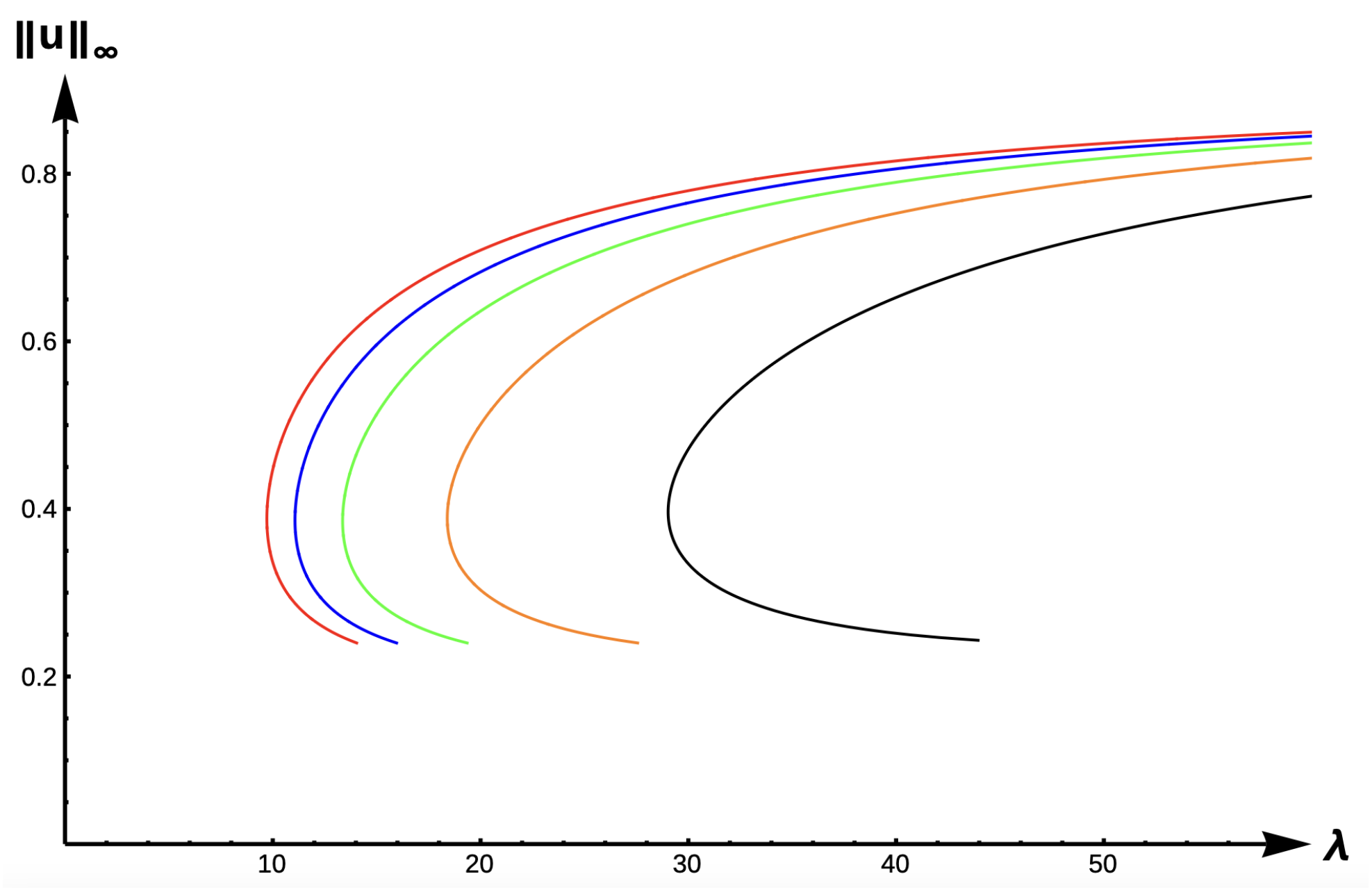
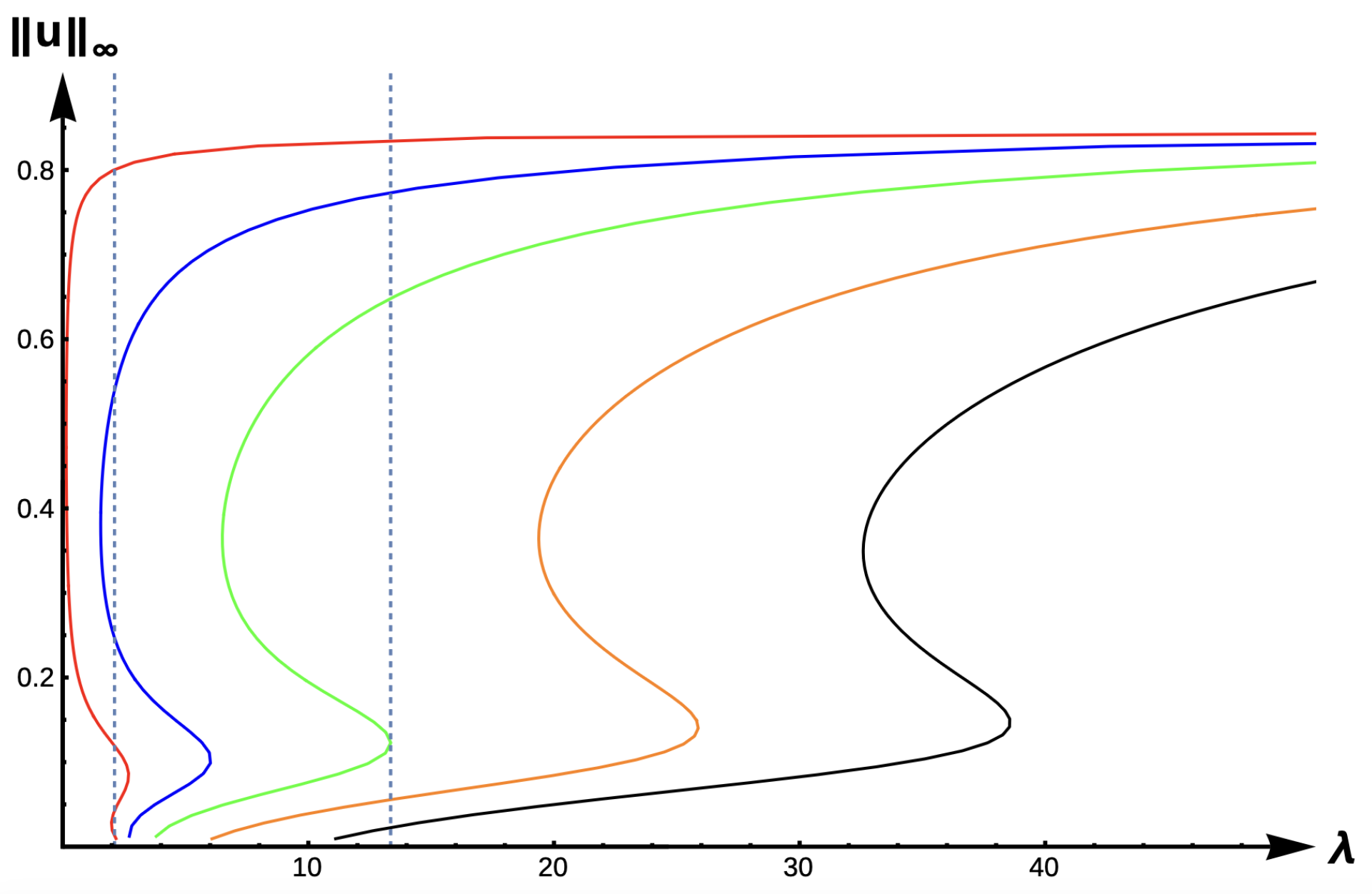
2.2.3. Bifurcation Diagrams for −DDE:
Figure 15 and
Figure 16 show the bifurcation curves for the −DDE case with strength
and
, respectively, for various PIE strength values
.
Table 8 and
Table 9 give
-values, uniqueness regions, and multiplicity regions for various values of
for
and
, respectively. First, we observe that for any fixed DDE strength
, the bifurcation curves translate to the right as
increases, approaching the bifurcation curve of the completely hostile matrix case (Dirichlet boundary condition) when
. For
, model predictions are again similar to those of the previous DIE and +DDE cases. However, when the DDE strength is sufficiently high (as in
Figure 16 with
), model predictions become much more interesting. For small PIE strength (
, red, blue, green, and orange in
Figure 16), there is a range of patch sizes where a patch-level Allee effect (PAE) is predicted by the model. For a patch with size corresponding to a
-value in this range, the model predicts that the population will need to remain above a certain threshold in order to persist. Since the growth term is logistic, this PAE arises solely from the −DDE relationship. Similar cases of PAE have been noted by previous authors for the case when
(see [
18,
32,
37,
42]). However, for higher PIE strength (
, black in
Figure 16), no such PAE is present and unconditional persistence is predicted by the model for patch sizes corresponding to
.
Notice the PIE envelope between the red and black curves in
Figure 16 provides a varied range of predictions crucially dependent upon the PIE strength. As an example, a patch with size corresponding to
would have predictions of a PAE for
and extinction for
. For a patch corresponding to
, the model predicts the population is not as sensitive to PIE strength with unconditional persistence predicted for low PIE strength, a PAE for medium levels of PIE, and extinction predicted for high sensitivity to predation. For patches with corresponding
outside of this envelope, predictions of unconditional persistence for
and extinction for
are unaffected by PIE altogether.
2.3. Results for Case C: Prey Switching Predation:
In this subsection, we present the results for the prey switching predation (Holling Type III) case, where we have fixed
and
. We note that
has two zeros (0 and 0.849) when
and
. Here, Lemma 3 holds when
and
,
have similar shapes to those in
Figure 12.
2.3.1. Bifurcation Diagrams for DIE:
For these results, we fix the effective matrix hostility at
.
Figure 17 shows the bifurcation curves for the DIE case for various
-values. Regardless of the PIE strength, the model predicts one of four outcomes: (1) extinction for
, (2) unconditional persistence at a low maximum density level, (3) multiple positive steady states (one at low max density and one at high density), or (4) unconditional persistence at a high maximum density level.
Table 10 shows the evolution of the uniqueness region as
varies and
Table 11 gives multiplicity regions. We also observe that there are three solutions for a certain interval of
between the uniqueness regions. The
interval where the multiplicity occurs translates to the right as
increases. The variation in
-values is given in
Table 12 and we observe that the bifurcation curves translate to the right, approaching the bifurcation curve of the completely lethal matrix case (Dirichlet boundary condition) when
.
As in previous cases, the envelope of
-values for which model predictions are especially sensitive to PIE strength is given between the red and black curves in
Figure 17. To highlight the effects of PIE, we first consider the example of a patch with size corresponding to
in
Figure 17. For a small PIE strength (
), there is a non-Allee type bi-stability as denoted in case (3) above; however, for large PIE strength, unconditional persistence at a low density level (as in case (2) above) is predicted. For a larger patch with size corresponding to
, cases (2)–(4) from above are possible as the PIE strength increases. Again, the sensitivity of the predation–emigration relationship causes vast population dynamical outcomes, even for the same patch size and matrix quality.
2.3.2. Bifurcation Diagrams for +DDE: ,
For the +DDE case, we again fix
.
Figure 18 shows how the bifurcation curves translate to the right as the PIE strength
increases. Corresponding
-values are given in
Table 12, while
Table 10 shows the variation in the uniqueness regions as
changes. The
-interval where the multiplicity occurs translates to the right as
increases. Model predictions are identical to those of the DIE case.
2.3.3. Bifurcation Diagrams for −DDE:
For the final case of −DDE,
Figure 19,
Figure 20 and
Figure 21 show the evolution of bifurcation curves for
, respectively, as
varies.
Table 13 gives uniqueness regions,
Table 14 gives multiplicity regions,
Table 15 gives patch level Alllee effect regions, and
Table 16 provide uniqueness regions and
-values, respectively, as
varies. The case of low DDE strength (
) gives a scenario that is identical to the DIE and +DDE cases but with a leftward translation in the curves as illustrated in
Figure 19. However, the envelope created by the red and black curves in
Figure 20 and
Figure 21 show a myriad of possible model predictions.
In the case of medium DDE strength (
in
Figure 20), potential model predictions include (1) extinction for
, (2) a PAE, (3) unconditional persistence at a low maximum density level, (4) non-Allee effect type bi-stability (one positive steady state at low max density and one at high density), or (5) unconditional persistence at a high maximum density level. For the high DDE strength case (
), all five of these dynamical outcomes are possible with the addition of a scenario as illustrated at
in the red curve of
Figure 21. For this
-value, a PAE occurs and persistence is possible at either a high maximum density or a small maximum density level. Increasing the PIE strength for this patch size will change model prediction to that of a PAE, followed by extinction for sufficiently high PIE strength. However, for a patch with size corresponding to
, outcomes (2)–(5) above are possible as the PIE sensitivity increases.
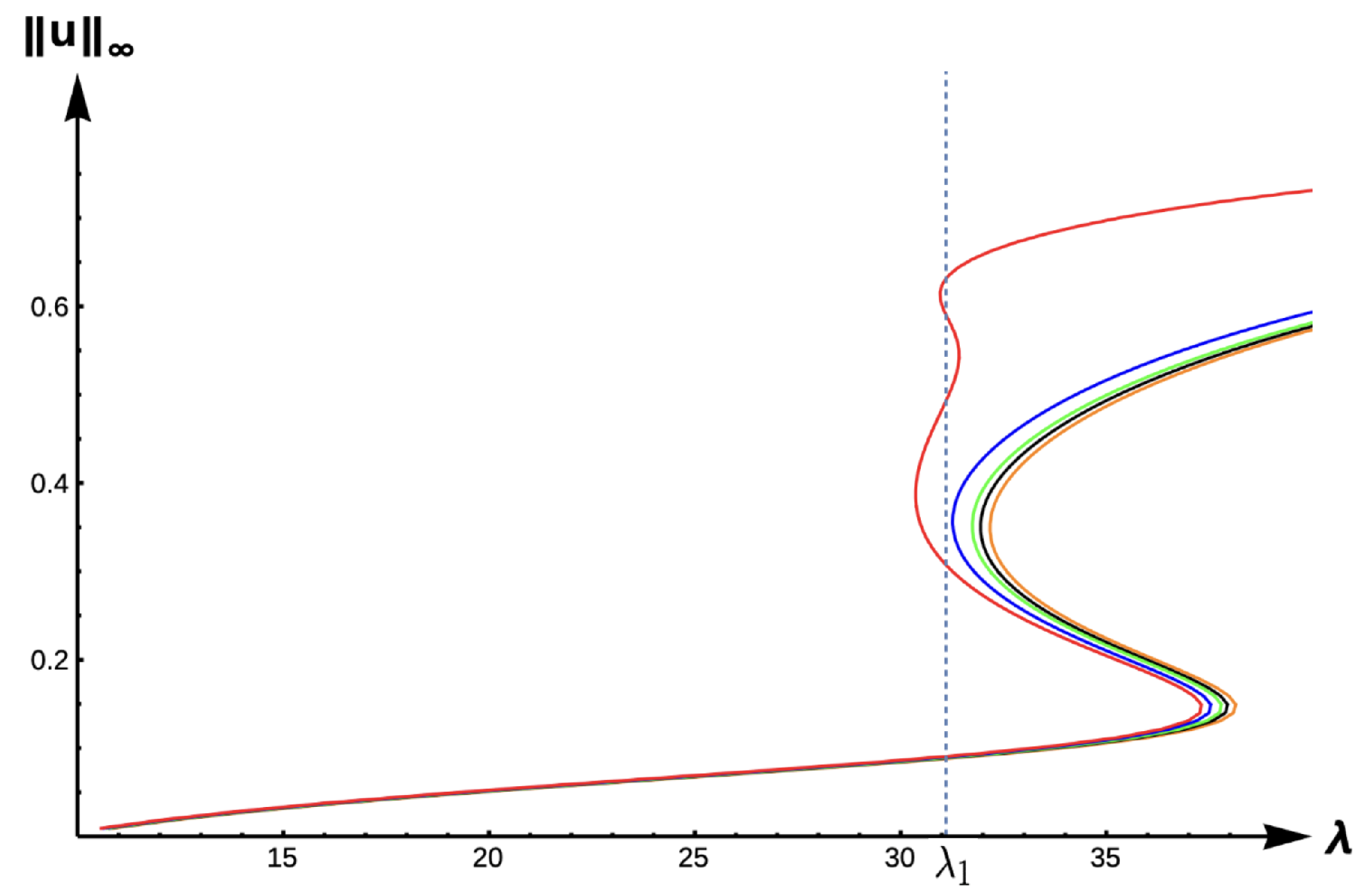
Next, we explore the effects of matrix hostility by considering the case with high DDE strength (
) and high matrix hostility (
).
Figure 22 illustrates the effect that extreme matrix hostility has on the model predictions, showing a new dynamical scenario at
. At this value, there are five positive solutions to the steady-state problem, showing a very complex situation. Here, persistence is possible but with a variety of different maximum density levels.
Figure 23 shows the evolution of bifurcation curves for
as the PIE strength increases.
3. Preliminaries
In this section, we introduce definitions of the (strict) subsolution and (strict) supersolution of (
1), state a sub-supersolution theorem that is used to prove the existence and multiplicity results of positive solutions, and state a lemma which we employ to numerically generate bifurcation curves.
By a subsolution of (
1), we mean
that satisfies
By a supersolution of (
1), we mean
that satisfies
By a strict subsolution (supersolution) of (
1), we mean a subsolution (supersolution) which is not a solution. Then, the following results hold (see [
43,
44,
45]):
Lemma 1. Let ψ and Z be a subsolution and a supersolution of (1), respectively, such that . Then, (1) has a solution such that . Lemma 2. Let and be a subsolution and a supersolution of (1), respectively, such that . Let and be a strict subsolution and a strict supersolution of (1), respectively, such that , and . Then, (1) has at least three solutions, , and where and . Let
u be a positive solution of (
7) when
. Since
is increasing for all
it follows that
u must be symmetric about
and has the shape as in
Figure 24 below (see [
32]). Let
and
.
Then the following result holds:
Lemma 3. (See [
32]).
For (7) has a positive solution u such that , , with if and only if λ, ρ, and q satisfyandwhere and . Remark 4. We provide the proof of Lemma 3 in the appendix for the convenience of the reader.
Next, we briefly explain how we obtain bifurcation curves using (
8)–(
9). Let
be the unique positive falling zero of
f and
for some
, where
is the smallest positive zero of
F when
and
when
or
. Letting
, we numerically solve the Equation (
9) for
q using the FindRoot command in Mathematica. The values of
q and
are substituted into (
8) to find the corresponding value of
. Repeating this procedure for
,
, we obtain
points for the bifurcation diagram.
We note that, in our study, when
, the branch of positive solutions bifurcates from
at
, where
is the principal (first positive) eigenvalue of
with an eigenfunction
. Here,
when
and
when
.
4. Construction of Subsolutions and Supersolutions to Prove Theorems 1–6
Here, we state a couple of eigenvalue problems which are crucial to our proofs and recall some properties of their respective principal eigenvalues. For
, let
be the principal eigenvalue,
be the corresponding normalized eigenfunction of
and let
be the principal eigenvalue and
be the corresponding normalized eigenfunction of
Note that the existence of both principle eigenvalues is standard (see [
15,
46]). For simplicity of notation, we denote
with corresponding normalized eigenfunction
for
.
The following lemma gives several useful properties of
and
(see [
15,
46,
47]).
Lemma 4. Let , denote the principal eigenvalue of (11), the principal eigenvalue of (12), and the principal eigenvalue of (2). Then, we have the following for : - (1)
for .
- (2)
for .
- (3)
is decreasing in M and increasing in b and γ.
- (4)
.
- (5)
is decreasing in M and increasing in b and γ.
Now, we present the construction of several crucial sub- and supersolutions for (
2).
Construction of a subsolution when for , or and (DIE), (+DDE) or (−DDE), where when and when .
For a fixed
, recall that
is the principal eigenvalue and
the corresponding normalized eigenfunction of (
12). We note that
for
. Let
for
and
Then, we have
and
since
. Therefore,
. This implies that
for
. We also have
for
since
and
for
Hence,
is a subsolution of (
1) for
.
Consider the following problem:
Let
be the unique positive solution of (
13) for
(see [
15]), where
is the principal eigenvalue of (
4).
We note that
as
Let
. Then, we have
Also,
by the Hopf maximum principle. Therefore,
is a subsolution of (
1) for
such that
as
.
Construction of a strict subsolution in when , , and when for some
Let
where
I is as in (
3). Choose
and
in (
11). Next, for a fixed
, let
be the principal eigenvalue and
be the corresponding normalized eigenfunction of (
11). We note that
when
. Define
for
. Then,
and
since
,
, and
. This implies that
for
. Let
be such that
Next, we define
where
. Observe that
, and
implies that
Let
and define
From (
14), we have
Next, since
, we have
Let
be such that
This implies that
Observe that
and
for
Now, for
, we have
by (
15). Hence,
is a strict subsolution of (
1) for
and
Construction of a strict subsolution when & hold for .
Let
be such that
is non-decreasing on
on
and
on
Then, the following boundary value problem
has a solution
such that
for
provided
and
are satisfied (see [
33]). Let
Since
on
and
on
by the Hopf maximum principle, it is easy to show that
is a strict subsolution of (
1) for
.
Construction of a strict supersolution for , , , , or , and any form of g.
is a global supersolution of (
1) for all
.
Construction of a strict supersolution for , for , or and any form of g where when and when .
For a fixed
, recall
is the principal eigenvalue and
the corresponding normalized eigenfunction of (
12). We note that
for
(see Lemma 4). Let
and
Since
and
, we have
and
giving that
for
. This implies that
and
for
since
Hence,
is a strict supersolution of (
1) for
and
.
Construction of a small supersolution for and when and (DIE) or (+DDE).
For a fixed
, recall
is the principal eigenvalue and
is the corresponding normalized eigenfunction of (
11) (here,
and
). We note that
for
(see Lemma 4). Define
, where
. Then, we have
Also, we have
since
when
(DIE) or
(+DDE). This implies that
is a supersolution of (
1) when
. Since
as
,
as
and hence,
as
.
Construction of a strict supersolution when & hold for
Let
where
is the unique positive solution of (
5). Thus, we have
since
and
Now, since
and
, we have
Hence,
is a strict supersolution of (
1) with
.
5. Proofs of Theorems 1–4
In this section, we provide proofs of our main results.
Proof of Theorem 1. We note that the problem
has a positive solution
for
such that
as
(see [
48]). Let
. Then, we have
Also,
by the Hopf maximum principle. Therefore,
is a subsolution of (
1) for
. Further,
is a global supersolution of (
1). Then, by Lemma 1, it follows that (
1) has at least one positive solution in
for
This implies that (
1) has at least one positive solution
for
such that
as
since
as
. □
Proof of Theorem 2. We first prove here the non-existence of a positive solution for
. Let
be the principal eigenvalue and
be the corresponding normalized eigenfunction of (
11). We note that
when
(see Lemma 4). Suppose
is a positive solution of (
1) for
. Then, by Green’s Second Identity, we have
On the other hand, we have
since
and
when
. This is a contradiction. Thus, (
1) has no positive solution for
We prove the existence of a positive solution, , for such that as
Recall the subsolution
for
and the supersolution
Since
, by Lemma 1, it follows that (
1) has a positive solution in
for
Also, recall the subsolution
for
Then, by Lemma 1, it follows that (
1) has a positive solution in
for
This implies that (
1) has a positive solution
for
such that
as
since
as
.
Next, we prove that
as
. Recall the subsolution
and supersolution
and choose
small enough such that
. Then, by Lemma 1, (
1) has a positive solution
such that
as
since
as
. But, the uniqueness of positive solutions of (
1) proved above implies that
. Hence, we have
as
Now, we prove the uniqueness of the positive solution for
. Suppose that (
1) has two distinct positive solutions,
, for
. Since
is a global supersolution, it follows that (
1) has a maximal solution. Thus, without loss of generality, we may assume that
Then, by Green’s Second Identity, we have
since
g is non-decreasing and
. We note that
is decreasing, and hence
since
. This is a contradiction. Hence, (
1) has at most one positive solution for
. □
Proof of Remark 1. Let
,
,
,
and
be the unique positive solutions of (
1) when
and
, respectively. Then, we have
It is clear that
is a strict subsolution of
Note that that
is a global supersolution of (
17). Hence, by Lemma 1, (
17) has a positive solution
. Since the solution of (
17) is unique by Theorem 2, we have
. Thus
. This completes the proof. □
Proof of Theorem 3. We note that
is a solution and hence a subsolution of (
1). Recall the strict subsolution
for
when
, strict supersolution
(with
) for
, and supersolution
for
We can also choose
to be small enough such that
. By Lemma 2, (
1) has at least two positive solutions,
and
, for
. Since
is a solution, Lemma 2 can only guarantee the existence of at least two positive solutions. Hence, there is a PAE for
.
We now show that if
and
are fixed, then there exists
such that (
1) has no positive solution for
when
. Choose
in (
11) such that
(see Lemma 4). For a given
, recall that
is the principal eigenvalue, and
is the corresponding normalized eigenfunction of (
11). We note that
for
(see Lemma 4). Now, suppose that (
1) has a positive solution
, for
. Then, we have
. This implies that
Clearly, there exists
such that
for
. Then, by Green’s Second Identity, we have
for
by (
18) and (
19). On the other hand, noting that
, we have
since
for
. This is a contradiction. Hence, (
1) has no positive solution for
when
. □
Proof of Theorem 4. The proof of this Theorem follows from the proof of Theorem 3. □
Proof of Theorem 5. Recall the subsolution for (here, ), strict subsolution for , supersolution for and strict supersolution for Since , we have By construction, we have Choosing , we have . Then, by Lemma 2, the result follows. □
Justification of the Remark 3:
Here, we prove that our growth term f and emigration term g satisfy the conditions in Theorem 5 when , and is a ball of radius R.
Recall that
. For simplicity of demonstration, we let
. Note that
and
has three positive solutions
, and
such that
(see
Figure 25). It can be shown that
, and
. Note that
increases on
and decreases on
with respect to
s. Hence
and
are the local maximum and
is a local minimum.
We choose
such that
, and
. Now, we will show that
which will imply
for
. By choosing
, we have
. This implies that
for
since
is an increasing function of
N. Then, we have
We note that the solution
of (
5) is given by
and
as
since
as
. Hence, we have
by (
20) for
. Hence, when
is large, we have
Thus, we have when is large for .
We note that the principal eigenvalue
(of (
4)) when
is a ball of radius
R is given by
where
is the first zero of the Bessel function of order
n. From [
33], we have
for
, and
when
. Then,
and
for
. On the other hand, we have
This implies that
when
is large for
since
as
. Thus, we have
and
holds when
. This completes the justification.
Proof of Theorem 6. Here, we prove the non-existence of a positive solution for
Note that
for all
. Let
be the principal eigenvalue and
be the corresponding normalized eigenfunction of (
11). We note that
when
(see Lemma 4). Suppose
is a positive solution of (
1) for
. Then, by Green’s Second Identity, we have
On the other hand, we have
since
and
for
. This is a contradiction. Thus, (
1) has no positive solution for
. □
Proof of Remark 3. Here, we prove non-existence for
in the Cases
A and
B. Recall that
for
. Assume
is a positive solution of (
1) for
. Then, by Green’s Second Identity, we have
since
. On the other hand, we have
This is a contradiction. Thus, (
1) has no positive solution for
. This completes the proof. □



 ,
, 










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}