
Establishment of Axenic Cultures for Two Marine Microalgae and the Effects of Antibiotics on Their Growth


Abstract
1. Introduction
2. Materials and Methods
2.1. Microalgal Strains and Culture Conditions
2.2. Antibiotics and Working Concentrations
2.3. Isolation and Identification of Phycospheric Bacteria
2.4. Antibiotic Sensitivity Testing of Phycospheric Bacteria
2.5. Establishment of a Sterile System
2.6. Sterility Testing and Antibiotic Removal
2.7. Effects of Antibiotics on Microalgae Growth
2.8. Data Analysis
3. Results
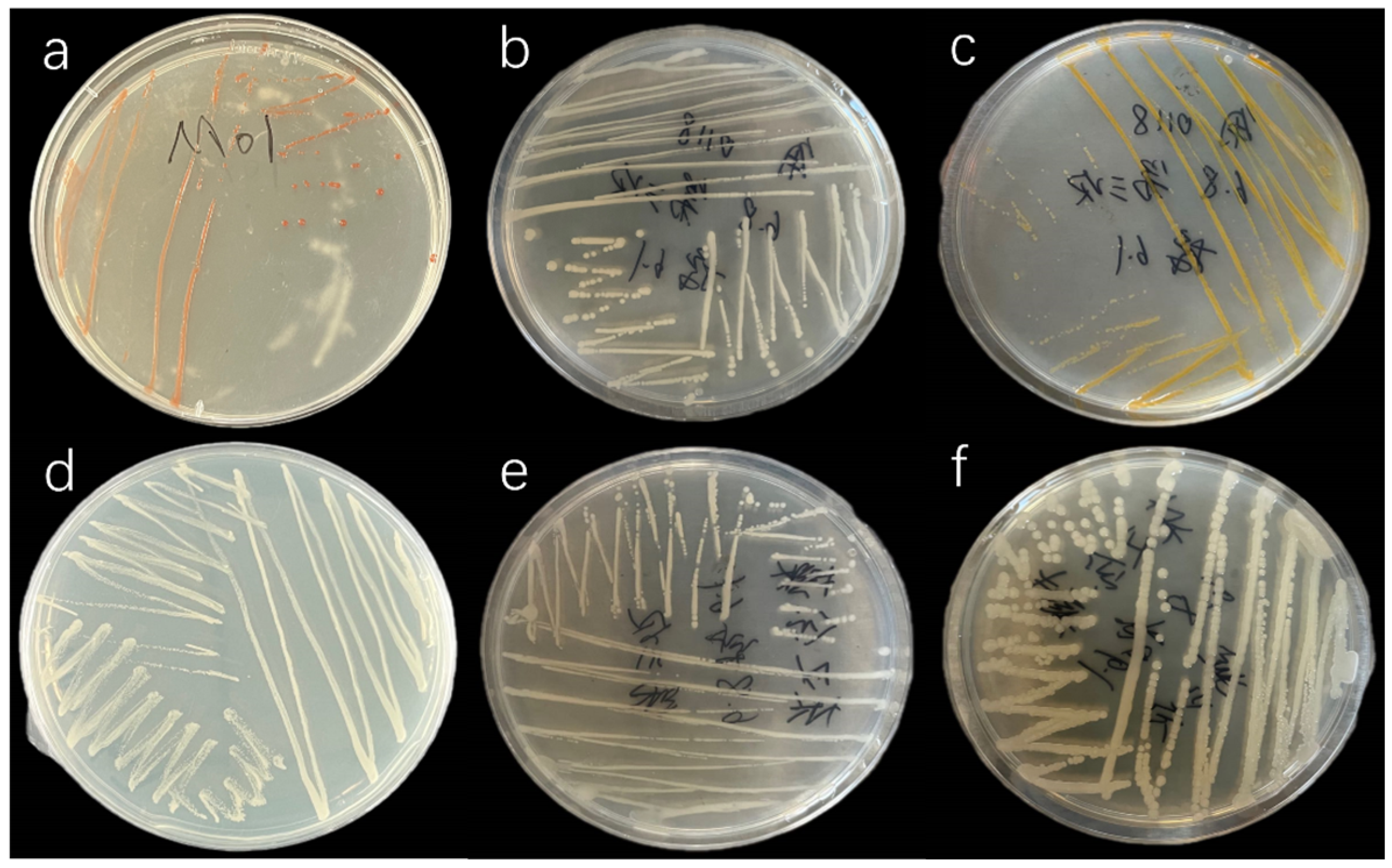
3.1. Isolation of Phycospheric Bacteria
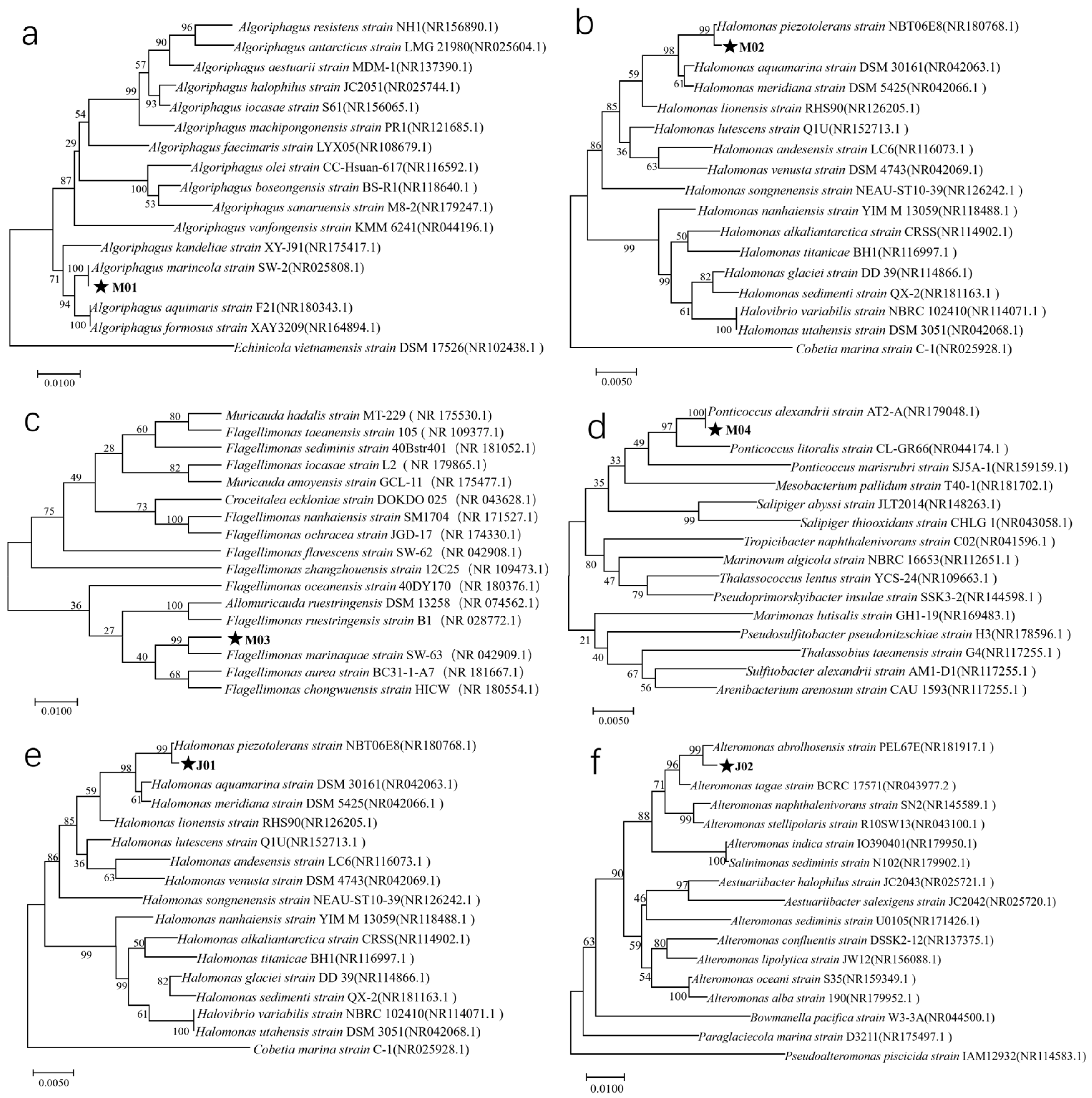
3.2. Identification of Phycospheric Bacteria Through 16S rDNA Sequencing
3.3. Sensitivity of Phycospheric Bacteria to Antibiotics
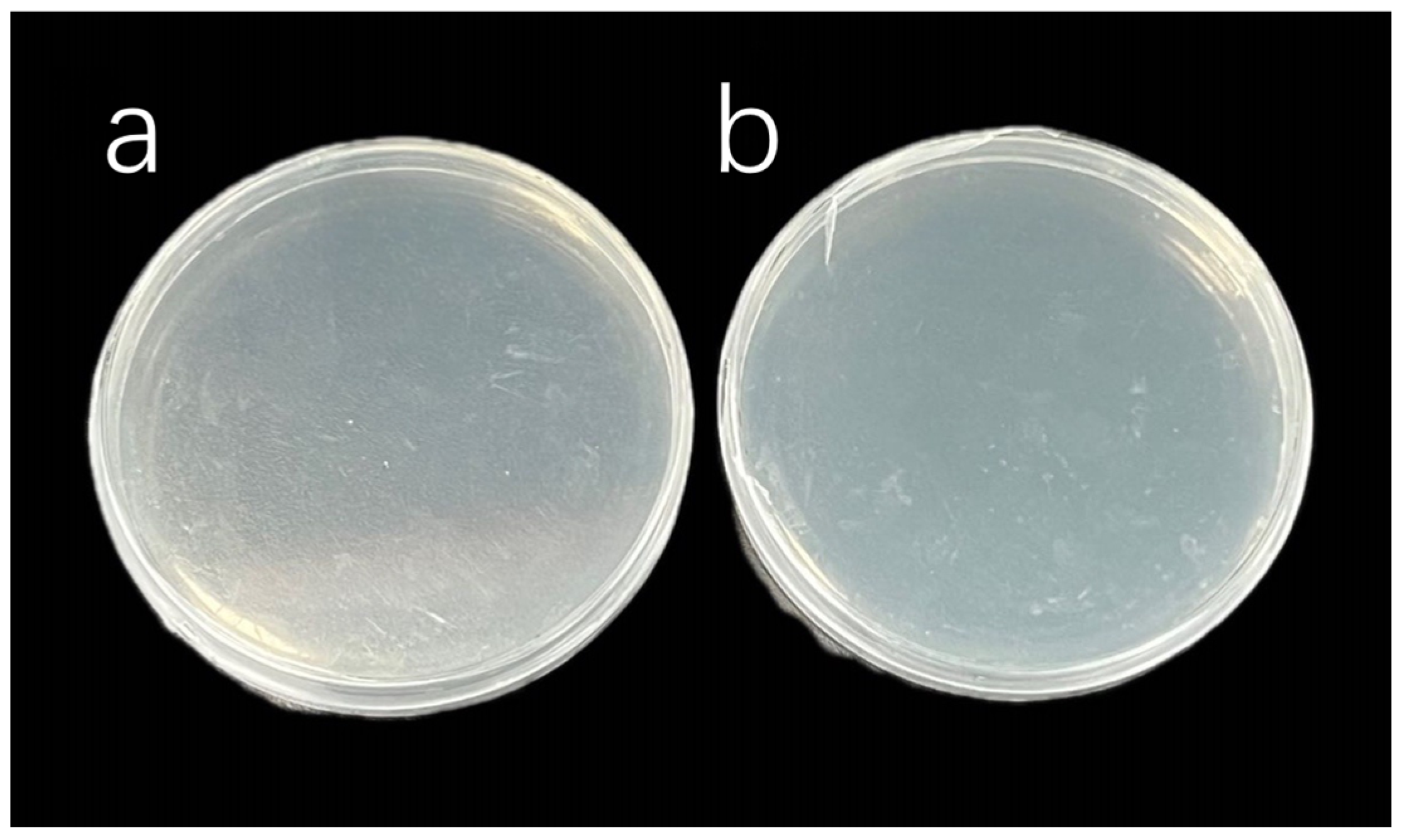
3.4. Algal Sterilization Treatment Results
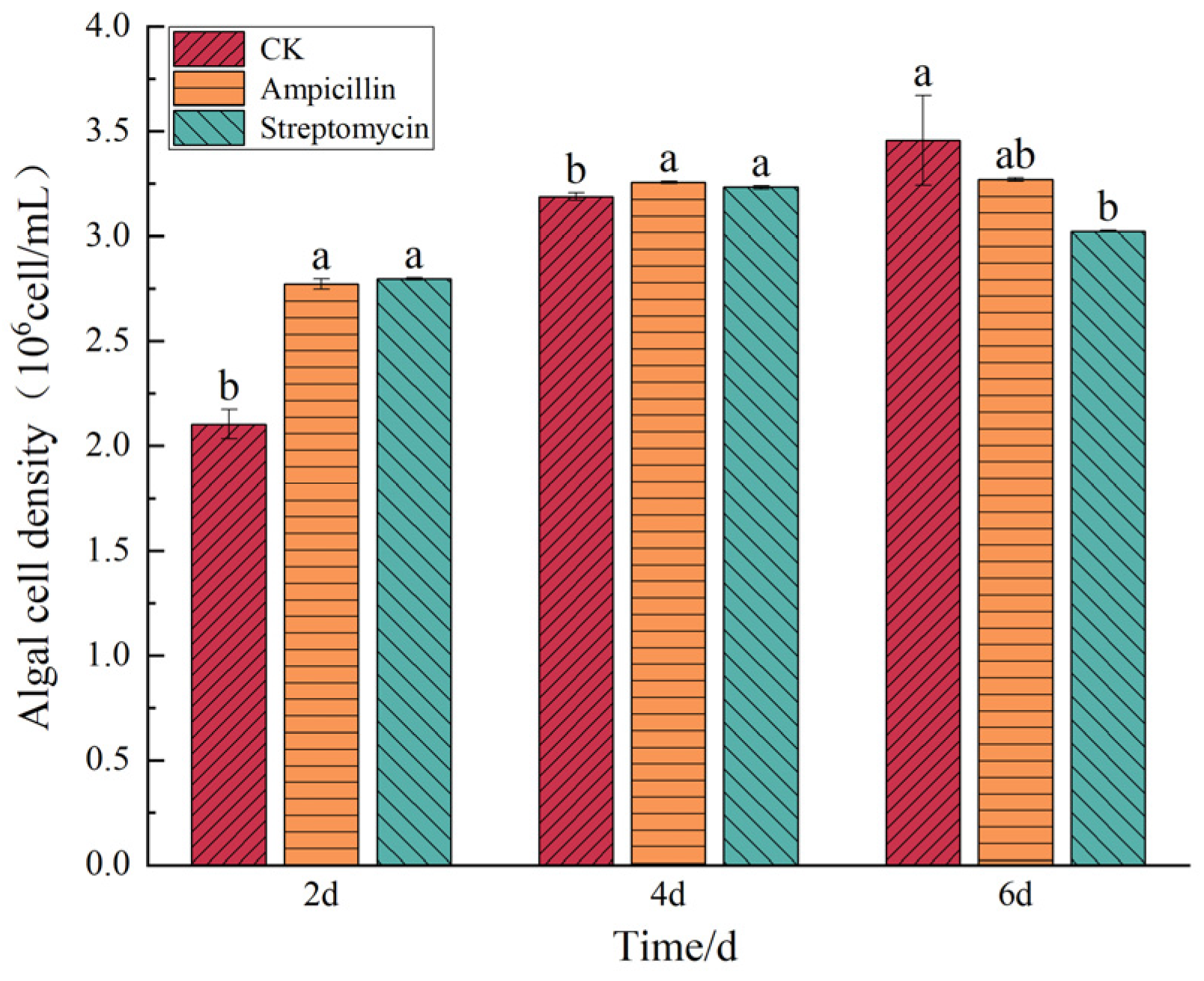
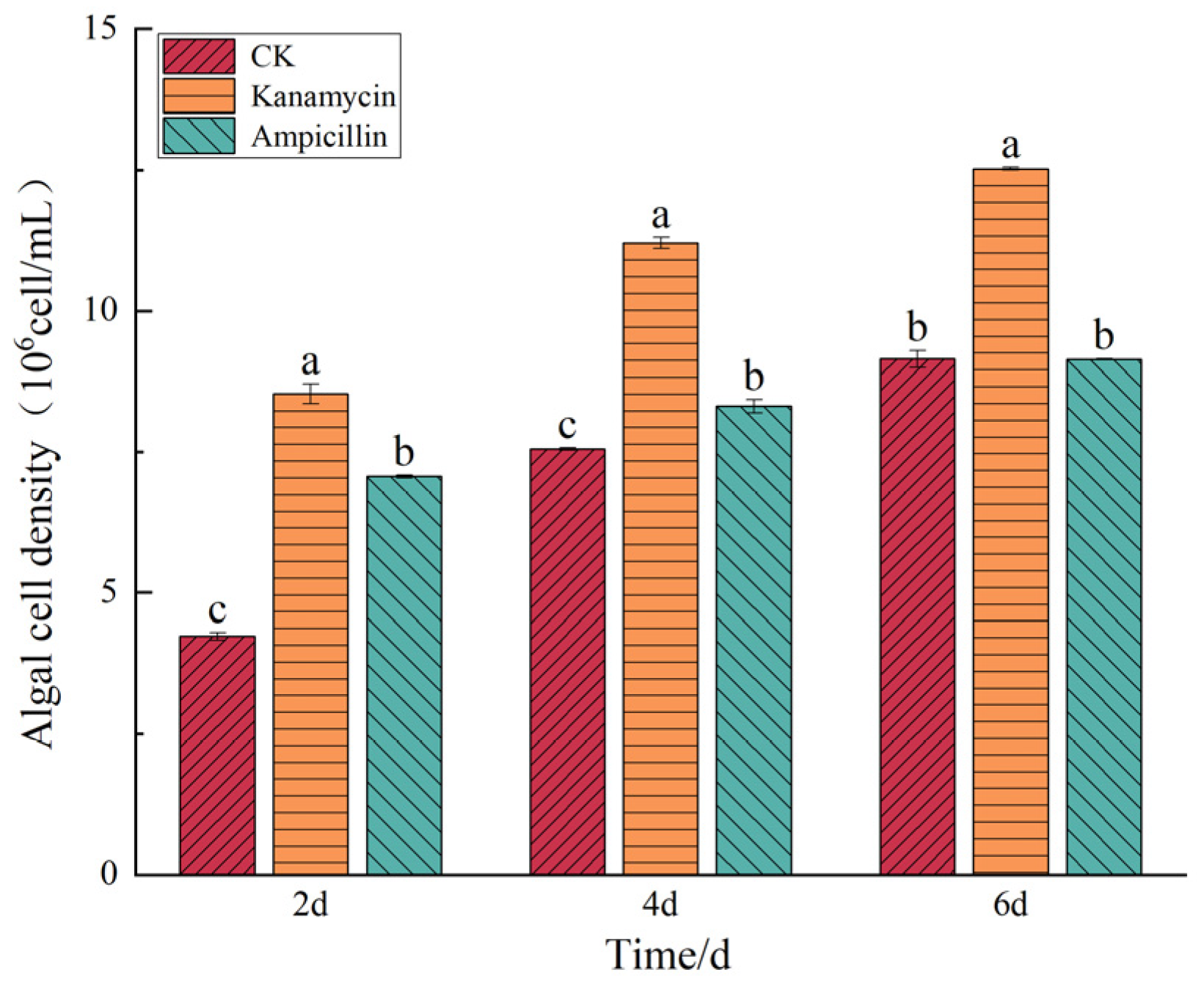
3.5. Results of the Effects of Antibiotics on Microalgae Growth
4. Discussion
4.1. Composition of Phycospheric Bacteria in Two Microalgal Species
4.2. Sterilization of Two Microalgal Species
4.3. Effects of Antibiotics on the Cell Density of the Two Microalgal Species
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhu, J.; Rong, J.; Zong, B. Factors in Mass Cultivation of Microalgae for Biodiesel. Chin. J. Catal. 2013, 34, 80–100. [Google Scholar] [CrossRef]
- Syafiuddin, A.; Boopathy, R. Effect of Algal Cells on Water Pollution Control. Curr. Pollut. Rep. 2021, 7, 213–226. [Google Scholar] [CrossRef]
- Jiang, X.; Han, Q.; Gao, X.; Gao, G. Conditions Optimising on the Yield of Biomass, Total Lipid, and Valuable Fatty Acids in Two Strains of Skeletonema menzelii. Food Chem. 2016, 194, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Gui, L.; Xu, L.; Liu, Z.; Zhou, Z.; Sun, Z. Carotenoid-Rich Microalgae Promote Growth and Health Conditions of Artemia nauplii. Aquaculture 2022, 546, 737289. [Google Scholar] [CrossRef]
- Shah, M.R.; Lutzu, G.A.; Alam, A.; Sarker, P.; Kabir Chowdhury, M.A.; Parsaeimehr, A.; Liang, Y.; Daroch, M. Microalgae in Aquafeeds for a Sustainable Aquaculture Industry. J. Appl. Phycol. 2018, 30, 197–213. [Google Scholar] [CrossRef]
- da Silva Vaz, B.; Moreira, J.B.; de Morais, M.G.; Costa, J.A.V. Microalgae as a New Source of Bioactive Compounds in Food Supplements. Curr. Opin. Food Sci. 2016, 7, 73–77. [Google Scholar] [CrossRef]
- Zhang, Z.; Tang, L.; Zhang, Y. Algae-Bacteria Interactions and Their Ecological Functions in the Ocean. Microbiol. China 2018, 45, 2043–2053. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Y.; He, J.; Deng, X. Isolation of bacteria in phycospher from Chaetoceros muelleri and its effect on growth and metabolism of algae. Feed Res. 2022, 45, 91–96. [Google Scholar] [CrossRef]
- Su, N.; Zhang, Y.; Yang, L.; Chen, H.; Gao, Q.; Chen, C.; Yang, Y. Effects of four microalgae on the survival and growth of Acanthaster spp. larvae. J. Appl. Oceanogr. 2023, 42, 633–642. [Google Scholar]
- Zhong, J.; Lv, X.; Li, G.; Fang, S.; Wang, Y. Mass Cultivation of Feed Organism for Amphioxus in Laboratory. Chin. Wild Plant Resour. 2006, 25, 44–47, 57. [Google Scholar]
- Huang, Z.; Liu, X.; Hu, Z.; Duan, S. Effects of antibiotics on the growth of Isochrysis zhangjiangensis and axenic culture. Ecol. Sci. 2007, 26, 120–121, 125. [Google Scholar]
- Wang, L.; Liu, X.; Ma, G.; Wang, L. Axenation of Dunaliella salina and the Effects of Axenic Cultivation on the Growth of the Strain. Anhui Agric. Sci. Bull. 2019, 25, 17–21. [Google Scholar] [CrossRef]
- Wang, W.; Sheng, Y. Gene Expression Concerning Fatty Acid and Amino Acid Metabolism in Chlorella vulgaris Cultured with Antibiotics. Appl. Microbiol. Biotechnol. 2020, 104, 8025–8036. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Choi, J.S.; Kong, I.S.; Park, S.I.; Kerr, R.G.; Hong, Y.K. A Procedure for Axenic Isolation of the Marine Microalga Isochrysis galbana from Heavily Contaminated Mass Cultures. J. Appl. Phycol. 2002, 14, 385–390. [Google Scholar] [CrossRef]
- Han, J.; Wang, S.; Zhang, L.; Yang, G.; Zhao, L.; Pan, K. A Method of Batch-Purifying Microalgae with Multiple Antibiotics at Extremely High Concentrations. Chin. J. Oceanol. Limnol. 2016, 34, 79–85. [Google Scholar] [CrossRef]
- Joo, H.-N.; Lee, C.-G. Antibiotics Addition as an Alternative Sterilization Method for Axenic Cultures in Haematococcus pluvialis. J. Ind. Eng. Chem. 2007, 13, 110–115. [Google Scholar]
- Chen, J.; Xu, J.; Zhang, S.; Liu, F.; Peng, J.; Peng, Y.; Wu, J. Nitrogen Removal Characteristics of a Novel Heterotrophic Nitrification and Aerobic Denitrification Bacteria, Alcaligenes faecalis Strain WT14. J. Environ. Manag. 2021, 282, 111961. [Google Scholar] [CrossRef]
- Kim, B.-H.; Ramanan, R.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Role of Rhizobium, a Plant Growth Promoting Bacterium, in Enhancing Algal Biomass through Mutualistic Interaction. Biomass Bioenergy 2014, 69, 95–105. [Google Scholar] [CrossRef]
- Munoz, R.; Guieysse, B. Algal-Bacterial Processes for the Treatment of Hazardous Contaminants: A Review. Water Res. 2006, 40, 2799–2815. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Cao, J.; Wu, M.; Xu, Y.; Wu, Y.; Shang, K.; Ma, B.; Zhang, L.; Chen, D.; Liu, X.; et al. Enhancing the Thermotolerance of Isochrysis zhangjiangensis Through Co-Culturing with Algoriphagus marincola. Mar. Biotechnol. 2023, 25, 463–472. [Google Scholar] [CrossRef]
- Wang, J.; Xie, Z.; Liu, Y.; Yan, F.; Cao, J.; Liu, R.; Wang, L.; Wei, Y.; Fang, J. Complete Genome Sequence of a Multiple-Stress-Tolerant Bacterium Halomonas piezotolerans NBT06E8T Isolated from a Deep-Sea Sediment Sample of the New Britain Trench. 3 Biotech 2022, 12, 236. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.; Kim, H.; Cha, I.; Joh, K. Flagellimonas maritima sp. Nov., Isolated from Surface Seawater. Int. J. Syst. Evol. Microbiol. 2020, 70, 187–192. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, X.; Li, L.; Zhang, R.; Feng, L.; Mu, J. Ponticoccus alexandrii sp. Nov., a Novel Bacterium Isolated from the Marine Toxigenic Dinoflagellate Alexandrium minutum. Antonie Van Leeuwenhoek 2018, 111, 995–1000. [Google Scholar] [CrossRef]
- Nóbrega, M.S.; Silva, B.S.; Leomil, L.; Tschoeke, D.A.; Campeão, M.E.; Garcia, G.D.; Dias, G.A.; Vieira, V.V.; Thompson, C.C.; Thompson, F.L. Description of Alteromonas abrolhosensis sp. Nov., Isolated from Sea Water of Abrolhos Bank, Brazil. Antonie Van Leeuwenhoek 2018, 111, 1131–1138. [Google Scholar] [CrossRef]
- Subashchandrabose, S.R.; Ramakrishnan, B.; Megharaj, M.; Venkateswarlu, K.; Naidu, R. Consortia of Cyanobacteria/Microalgae and Bacteria: Biotechnological Potential. Biotechnol. Adv. 2011, 29, 896–907. [Google Scholar] [CrossRef]
- Liu, J.; Sun, Y.; Zhang, L.; Li, X.; He, Z.; Zhou, C.; Han, J. Screening of Antibiotics to Obtain Axenic Cell Cultures of a Marine Microalga Chrysotila roscoffensis. Front. Bioeng. Biotechnol. 2023, 11, 1218031. [Google Scholar] [CrossRef] [PubMed]
- Kiki, C.; Rashid, A.; Wang, Y.; Li, Y.; Zeng, Q.; Yu, C.-P.; Sun, Q. Dissipation of Antibiotics by Microalgae: Kinetics, Identification of Transformation Products and Pathways. J. Hazard. Mater. 2020, 387, 121985. [Google Scholar] [CrossRef]
- Daneshvar, E.; Zarrinmehr, M.J.; Hashtjin, A.M.; Farhadian, O.; Bhatnagar, A. Versatile Applications of Freshwater and Marine Water Microalgae in Dairy Wastewater Treatment, Lipid Extraction and Tetracycline Biosorption. Bioresour. Technol. 2018, 268, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Backhaus, T.; Scholze, M.; Grimme, L.H. The Single Substance and Mixture Toxicity of Quinolones to the Bioluminescent Bacterium Vibrio fischeri. Aquat. Toxicol. 2000, 49, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Faust, M.; Altenburger, R.; Backhaus, T.; Bödeker, W.; Scholze, M.; Grimme, L.H. Predictive Assessment of the Aquatic Toxicity of Multiple Chemical Mixtures. J. Environ. Qual. 2000, 29, 1063–1068. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, Y.; Xiao, H.; Wang, R.; Qu, L.; Tang, X. Sensitivity of Several Marine Microalga to Antibiotics. J. Wuhan Univ. Nat. Sci. Ed. 2007, 53, 249–254. [Google Scholar]
- Huang, J.; Gong, X.; Tang, X.; Li, Y. Hormesis of streptomycin on 6 species of marine microalga. J. Ocean. Univ. Qingdao 2000, 30, 642–644. [Google Scholar]
- Zhou, J.; Zhang, L.; An, M.; Duan, S. Hormesis Effect of Organophosphorus Pesticide Glyphostate-Isopropylammonium on Heterosigma akashiwo. Ecol. Sci. 2012, 31, 396–400. [Google Scholar]
- Wang, H.; Cheng, M.; Yang, Y.; Li, S.; Yan, B. The Responses of Algae Pavlova viridis and Karenia mikimotoi to Three Conventional Antibiotics Stress. Fish Sci. 2012, 31, 329–332. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, X.; Zhang, J.; Gao, B. Hormesis Effects of Amoxicillin on Growth and Cellular Biosynthesis of Microcystis aeruginosa at Different Nitrogen Levels. Microb. Ecol. 2015, 69, 608–617. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Bacterial Characteristics | Gram Staining |
|---|---|---|
| M01 | Orange, round, medium to large, transparent | Positive |
| M02 | White, round, small, opaque | Positive |
| M03 | Yellow, round, small, opaque | Positive |
| M04 | White, round, small, opaque | Positive |
| J01 | White, round, small, opaque | Positive |
| J02 | White, round, small, slightly transparent | Positive |
| Inhibition Zone Diameter (mm) | Strain | |||
|---|---|---|---|---|
| Ampicillin (5 mg/mL) | Streptomycin (5 mg/mL) | Kanamycin (5 mg/mL) | Gentamicin Sulfate (5 mg/mL) | |
| ≥20 (High) | M01, M03 | M01, M03, M04 | M01, M04, J01/M02 | J01/M02, J02 |
| 11–19 (Medium) | J02 | J01/M02, J02 | J02 | M03, M04 |
| <10 (Low) | M04 | M03 | M01 | |
| Processing Methods | Group A: C. muelleri | Group B: C. muelleri | Group C: I. zhanjiangensis | Group D: I. zhanjiangensis |
|---|---|---|---|---|
| Antibiotic Addition | 10 mg/mL Ampicillin (7 mL) | 25 mg/mL Streptomycin (7 mL) | 25 mg/mL Kanamycin (7 mL) | 10 mg/mL Ampicillin (7 mL) |
| Inhibition Effect | M02, M04 not inhibited | M02, M03, M04 not inhibited | Not inhibited | J01 not inhibited |
| Additional Antibiotic Addition | 25 mg/mL Streptomycin (5 mL) | 10 mg/mL Ampicillin (5.5 mL) | 10 mg/mL Ampicillin (5 mL) | 25 mg/mL Kanamycin (5 mL) |
| Inhibition Effect | Not inhibited | Not inhibited | J01 not inhibited | Not inhibited and J02 appeared |
| Antibiotic Total Concentration in the Initial Culture System | Ampicillin: 0.331 mg/mL; Streptomycin: 0.590 mg/mL | Streptomycin: 0.824 mg/mL; Ampicillin: 0.259 mg/mL | Kanamycin: 0.824 mg/mL; Ampicillin: 0.259 mg/mL | Ampicillin: 0.331 mg/mL; Kanamycin: 0.590 mg/mL |
| Subsequent Addition of Antibiotics (First Generation) | 25 mg/mL Gentamycin Sulfate (10 mL) | 25 mg/mL Kanamycin, (6.5 mL) | None | 25 mg/mL Streptomycin, (10 mL) |
| Inhibition Effect | M02 not inhibited | Not inhibited | J01 not inhibited | J01 not inhibited |
| Antibiotic Addition | 25 mg/mL Streptomycin (7 mL) | 10 mg/mL Ampicillin (5.5 mL) | 25 mg/mL Streptomycin (12 mL) and 25 mg/mL Kanamycin (11 mL) | 25 mg/mL Gentamycin Sulfate (10 mL) |
| Antibiotic Total Concentration in First Generation | Gentamycin Sulfate: 1.152 mg/mL; Streptomycin 0.806 mg/mL | Kanamycin: 0.766 mg/mL; Ampicillin: 0.259 mg/mL | Streptomycin: 1.345 mg/mL; Kanamycin: 1.234 mg/mL | Streptomycin: 1.136 mg/mL; Gentamycin Sulfate: 1.136 mg/mL |
| Subsequent Addition of Antibiotics (Second Generation) | 10 mg/mL Ampicillin (10 mL) | 10 mg/mL Ampicillin (10 mL) | 10 mg/mL Ampicillin (10 mL) | 10 mg/mL Ampicillin (10 mL) |
| Inhibition Effect | Sterile | Not inhibited | Sterile | Not inhibited |
| Antibiotic Total Concentration in Second Generation | Ampicillin: 0.476 mg/mL | Ampicillin: 0.476 mg/mL | Ampicillin: 0.476 mg/mL | Ampicillin: 0.476 mg/mL |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Liang, S.; Liu, H.; Li, Y.; Wang, X.; Guo, Y.; Liang, J. Establishment of Axenic Cultures for Two Marine Microalgae and the Effects of Antibiotics on Their Growth. Water 2025, 17, 275. https://doi.org/10.3390/w17020275
Huang J, Liang S, Liu H, Li Y, Wang X, Guo Y, Liang J. Establishment of Axenic Cultures for Two Marine Microalgae and the Effects of Antibiotics on Their Growth. Water. 2025; 17(2):275. https://doi.org/10.3390/w17020275
Chicago/Turabian StyleHuang, Jin, Shuang Liang, Huiru Liu, Yongren Li, Xiaoyu Wang, Yongjun Guo, and Jian Liang. 2025. "Establishment of Axenic Cultures for Two Marine Microalgae and the Effects of Antibiotics on Their Growth" Water 17, no. 2: 275. https://doi.org/10.3390/w17020275
APA StyleHuang, J., Liang, S., Liu, H., Li, Y., Wang, X., Guo, Y., & Liang, J. (2025). Establishment of Axenic Cultures for Two Marine Microalgae and the Effects of Antibiotics on Their Growth. Water, 17(2), 275. https://doi.org/10.3390/w17020275