
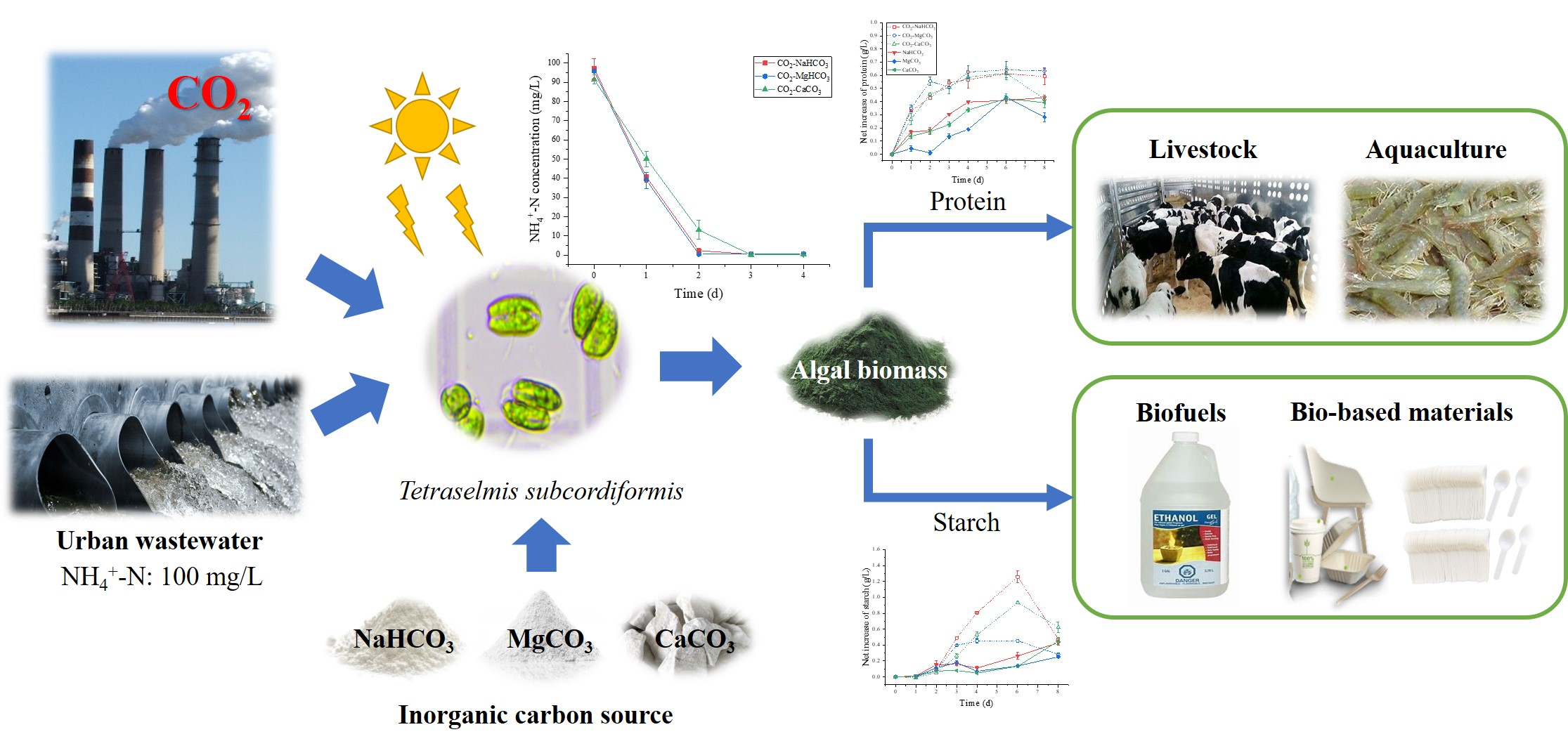
CO2-Inorganic Carbon Auto-Buffering System for Efficient Ammonium Reclamation Coupled with Valuable Biomass Production in a Euryhaline Microalga Tetraselmis subcordiformis

Abstract
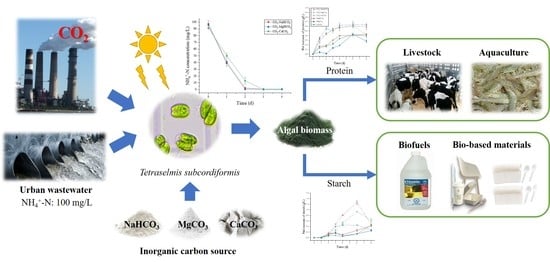
:
1. Introduction
2. Materials and Methods
2.1. Algal Strain and Culture Conditions
2.2. pH, Growth Measurement and Biochemical Component Analysis
2.3. Photosynthetic Performance Analysis
2.4. Ammonium-Nitrogen (NH4+-N) Analysis
2.5. Nitrogen Distribution and Total Organic Carbon (TOC) Analysis
2.6. Amino Acid Analysis
2.7. Statistical Analysis
3. Results and Discussion
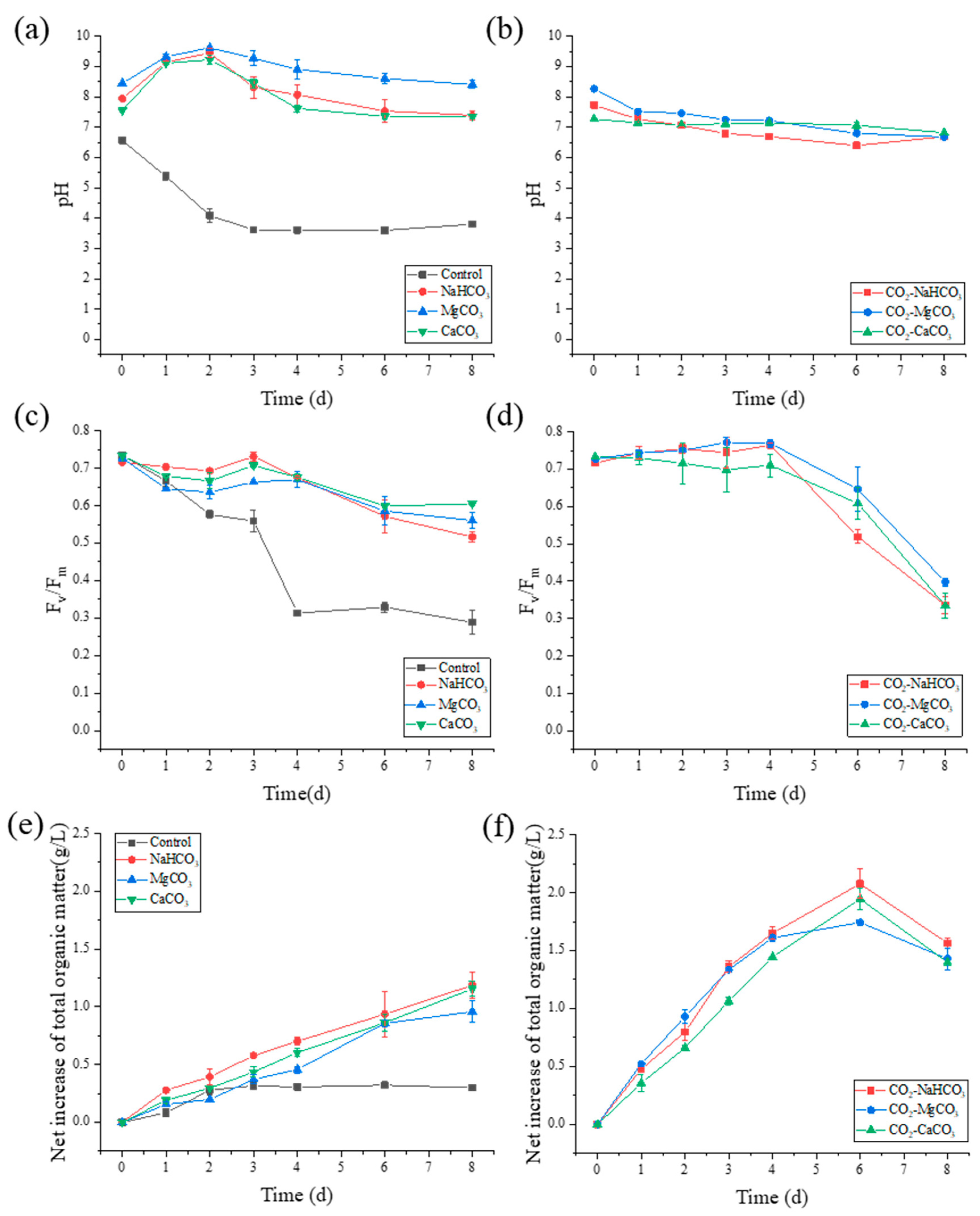
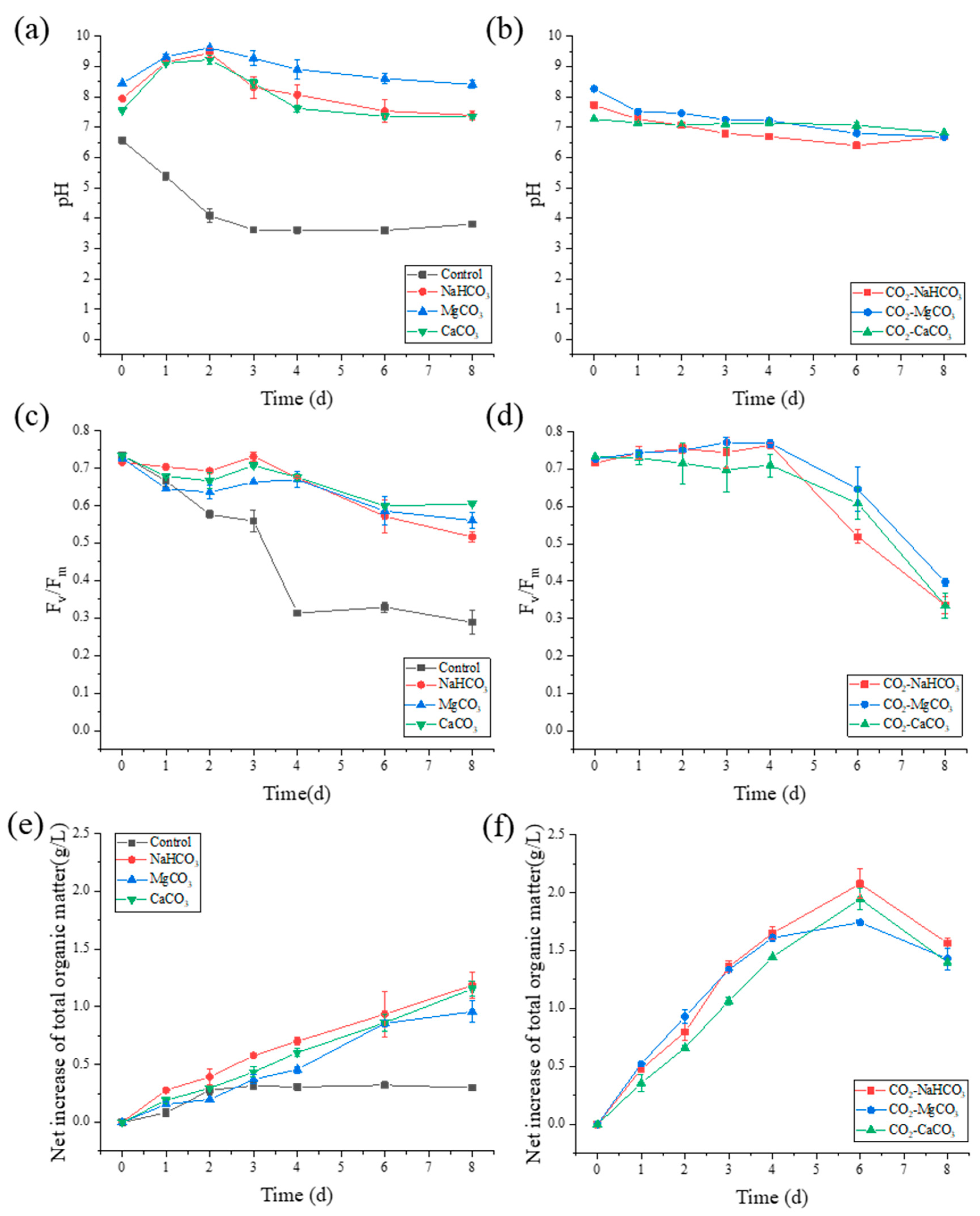
3.1. pH Variation, Photosynthetic Performance, and Cell Growth
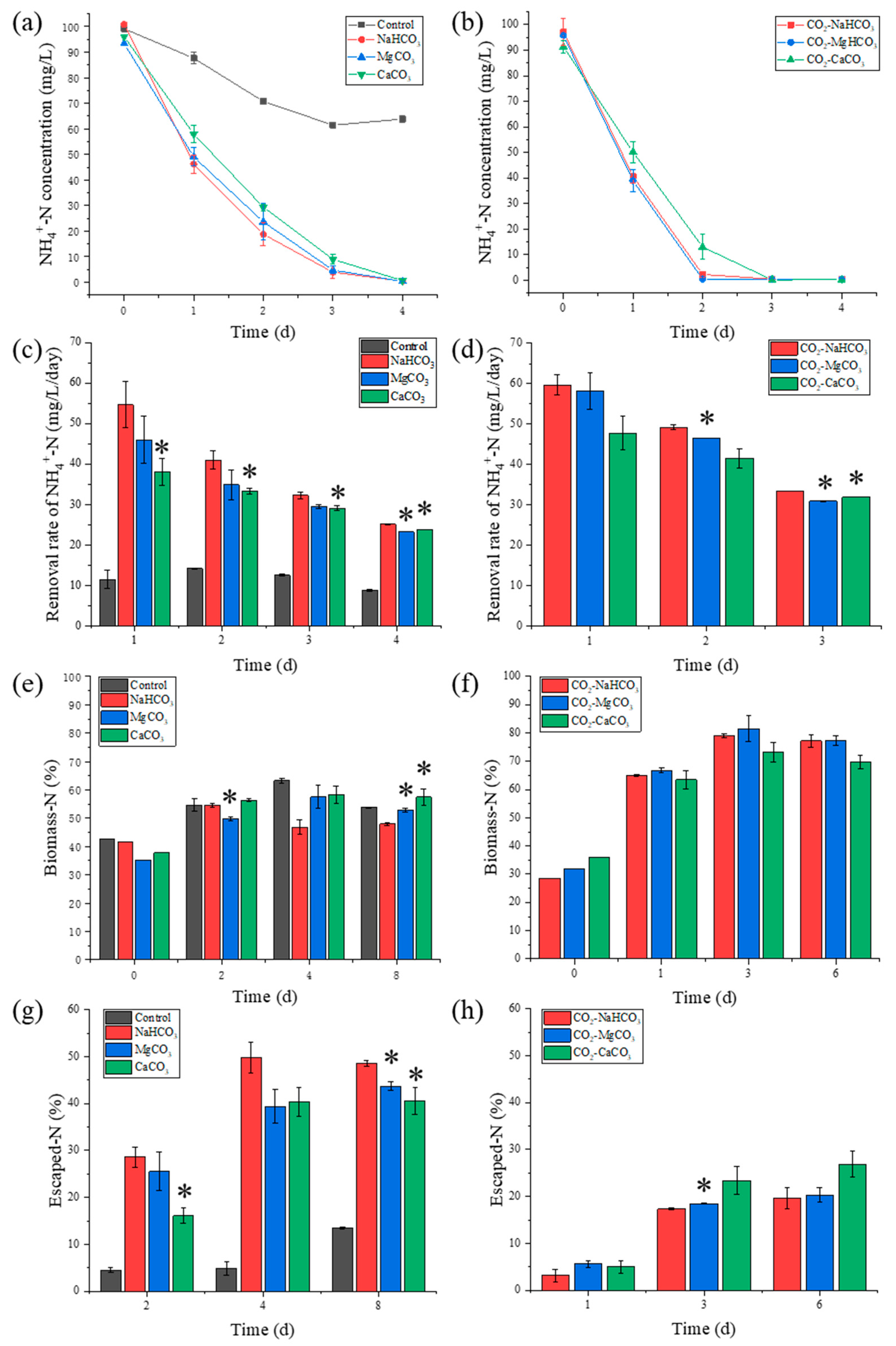
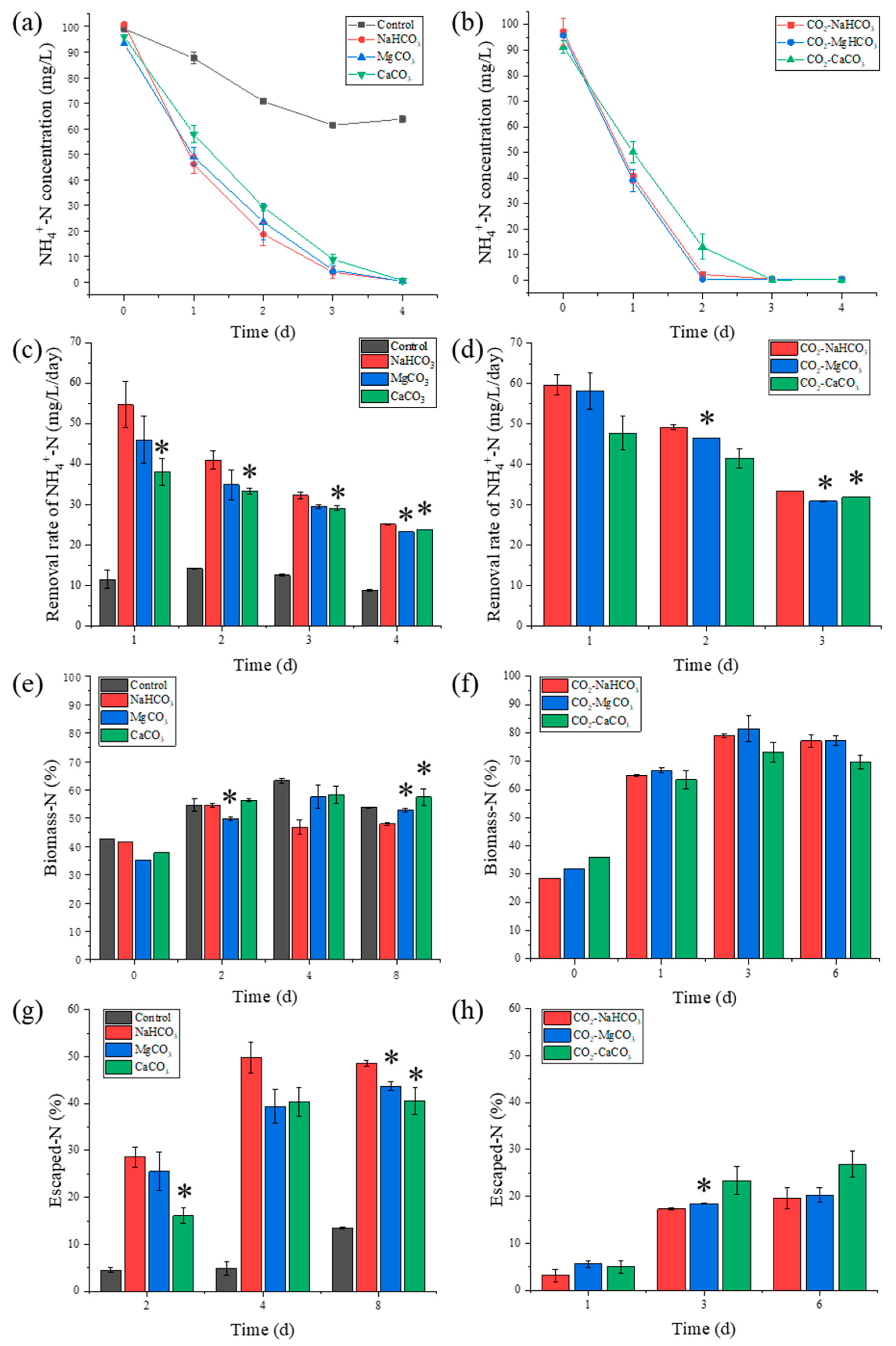
3.2. NH4+-N Removal and N Distribution
3.2.1. NH4+-N Removal
3.2.2. N Distribution
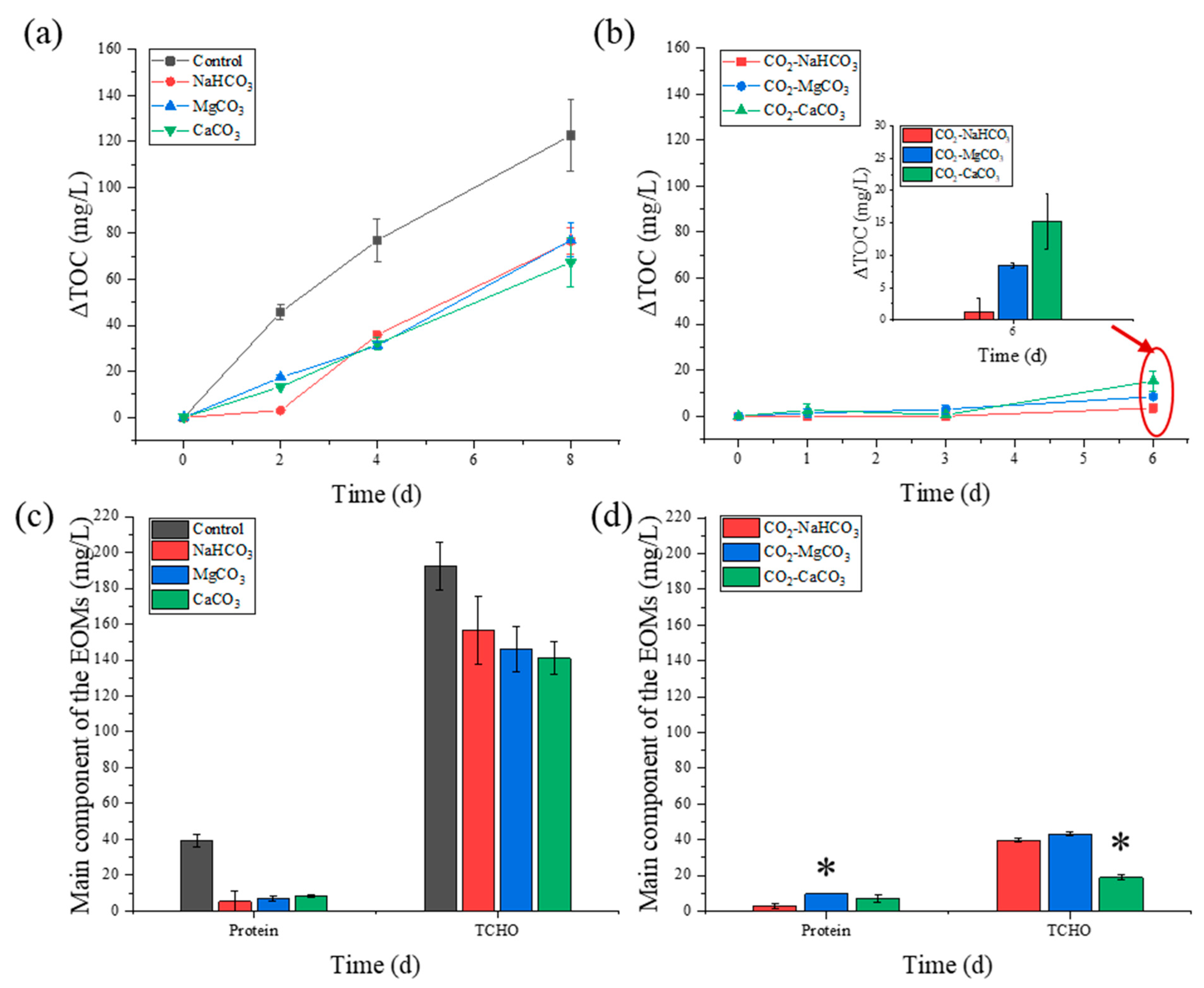
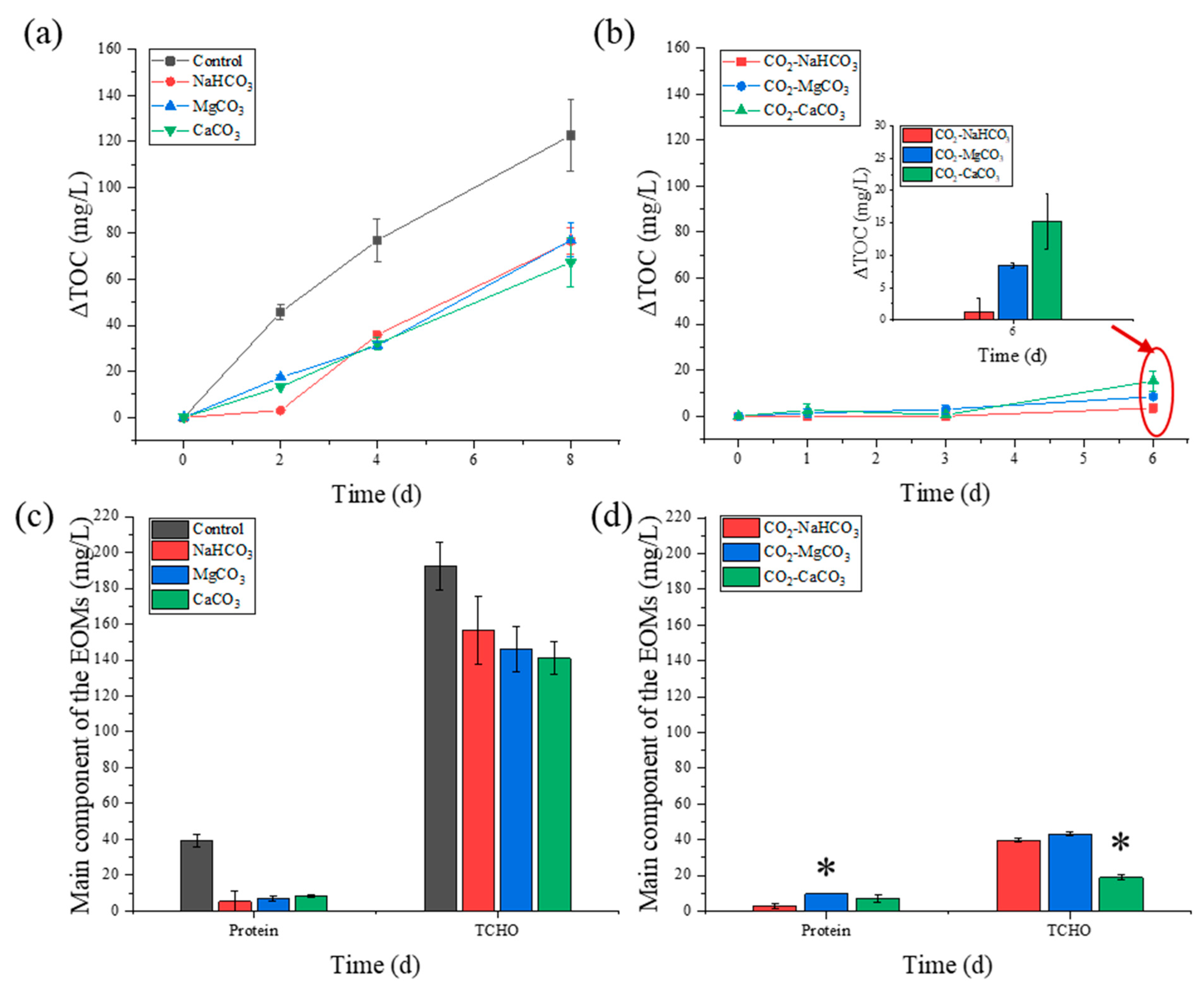
3.3. Extracellular Organic Matter Release during NH4+-N Removal
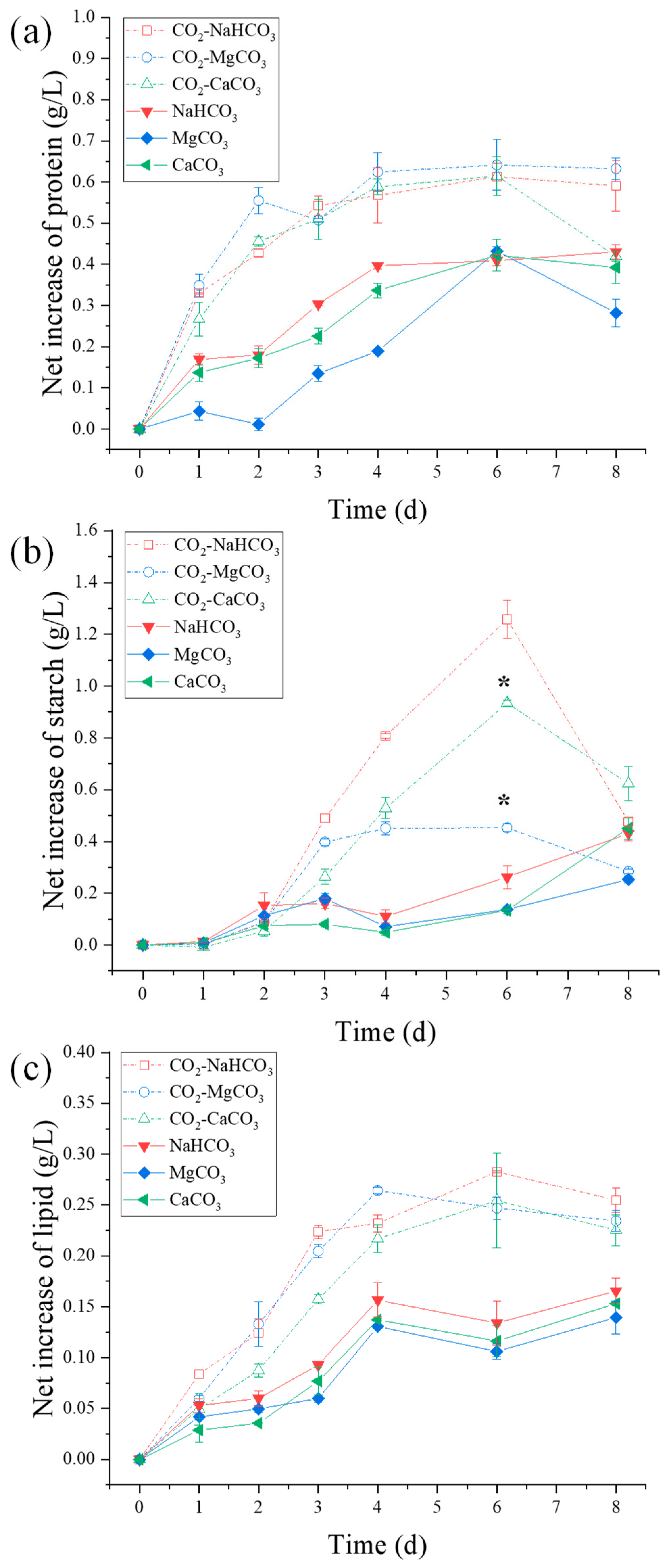
3.4. Biomass Component Production
3.5. Biomass Quality Evaluation
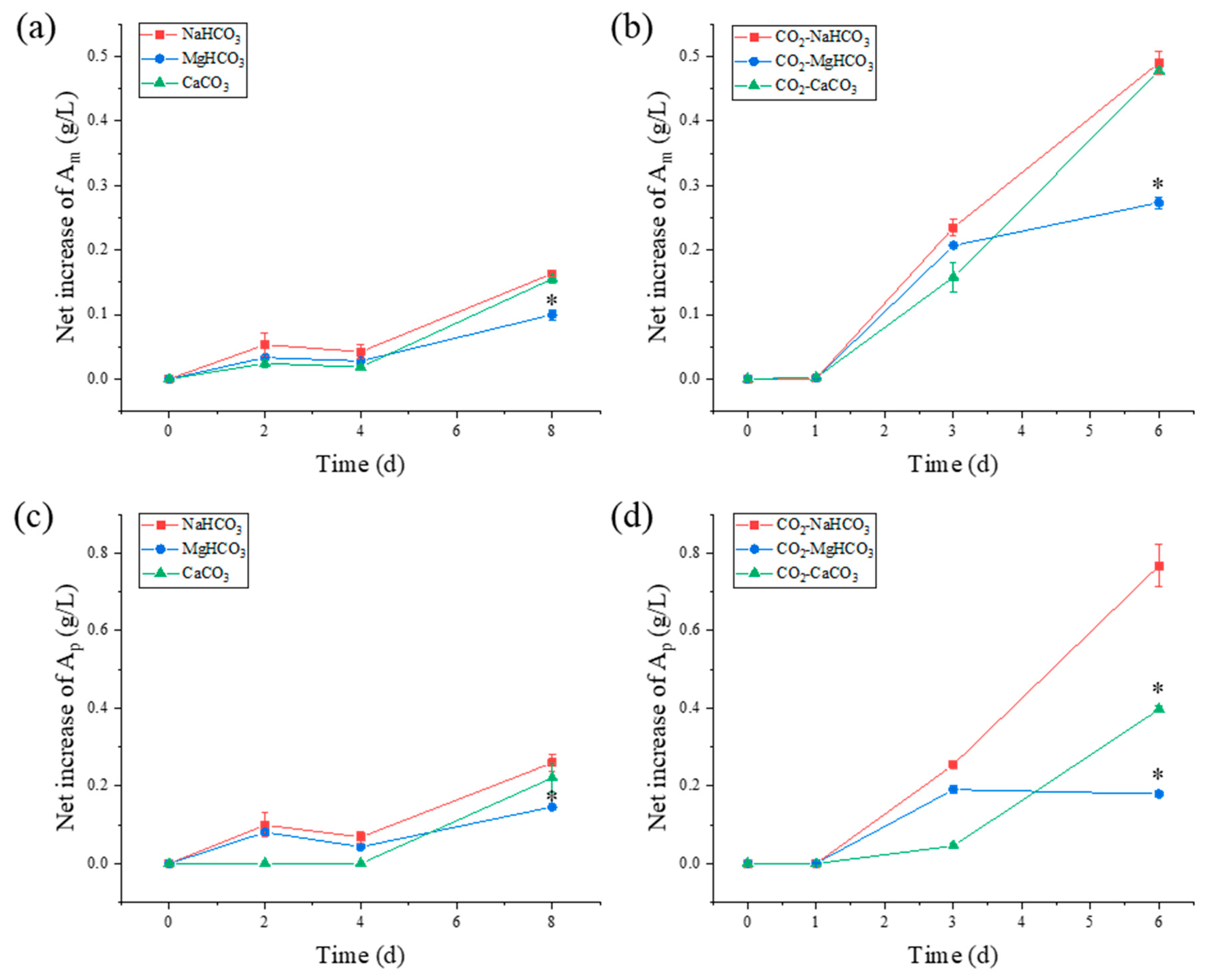
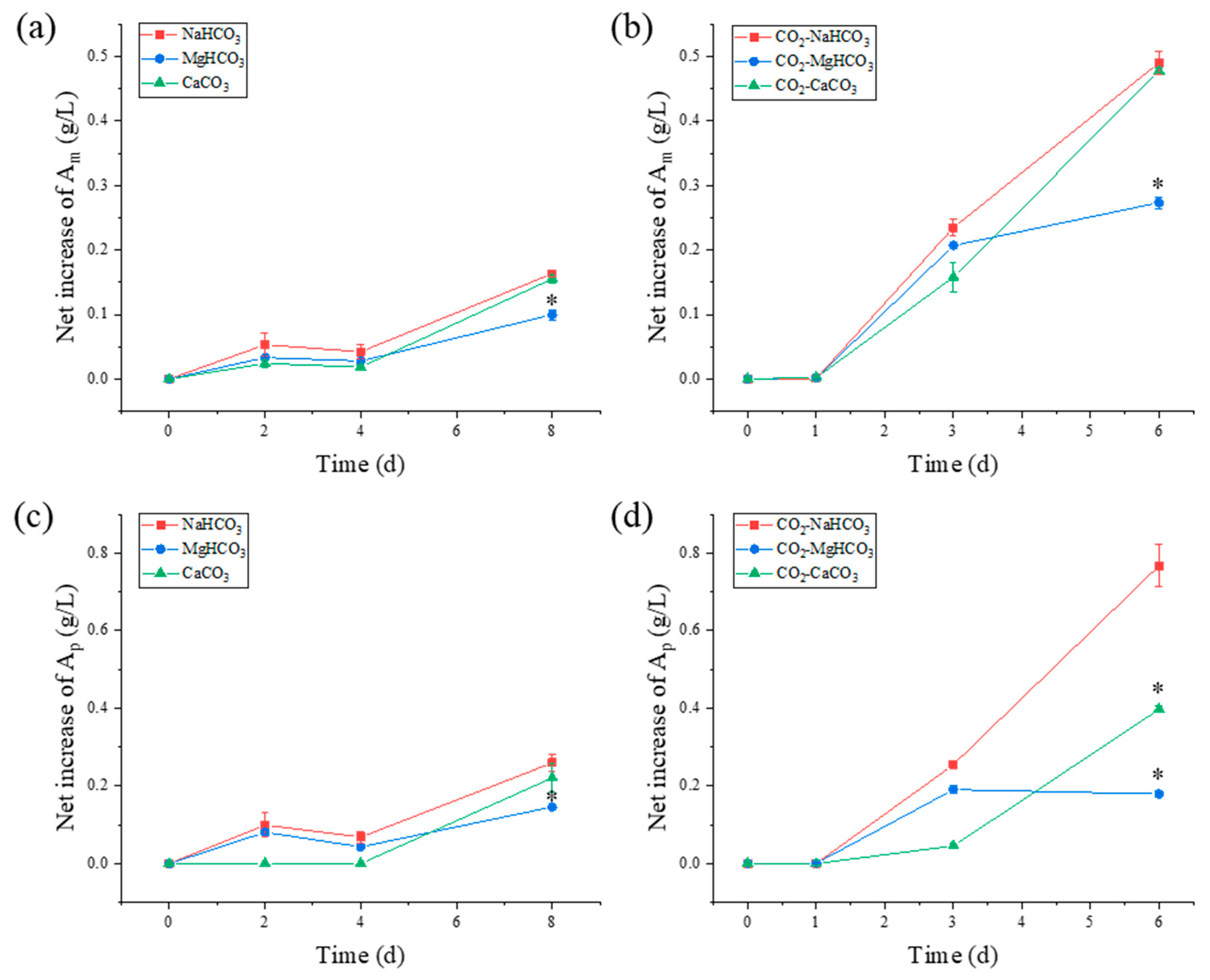
3.5.1. Starch Composition
3.5.2. Amino Acid Profile
3.6. Future Works
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Winkler, M.-K.H.; van Loosdrecht, M.C.M. Intensifying existing urban wastewater. Science 2022, 375, 377–378. [Google Scholar] [CrossRef] [PubMed]
- Selvaratnam, T.; Henkanatte-Gedera, S.M.; Muppaneni, T.; Nirmalakhandan, N.; Deng, S.; Lammers, P.J. Maximizing recovery of energy and nutrients from urban wastewaters. Energy 2016, 104, 16–23. [Google Scholar] [CrossRef]
- Rosemarin, A.; Macura, B.; Carolus, J.; Barquet, K.; Ek, F.; Järnberg, L.; Lorick, D.; Johannesdottir, S.; Pedersen, S.M.; Koskiaho, J.; et al. Circular nutrient solutions for agriculture and wastewater—A review of technologies and practices. Curr. Opin. Environ. Sustain. 2020, 45, 78–91. [Google Scholar] [CrossRef]
- Schulze, P.S.C.; Carvalho, C.F.M.; Pereira, H.; Gangadhar, K.N.; Schüler, L.M.; Santos, T.F.; Varela, J.C.S.; Barreira, L. Urban wastewater treatment by Tetraselmis sp. CTP4 (Chlorophyta). Bioresour. Technol. 2017, 223, 175–183. [Google Scholar] [CrossRef]
- Acién, F.G.; Gómez-Serrano, C.; Morales-Amaral, M.M.; Fernández-Sevilla, J.M.; Molina-Grima, E. Wastewater treatment using microalgae: How realistic a contribution might it be to significant urban wastewater treatment? Appl. Microbiol. Biotechnol. 2016, 100, 9013–9022. [Google Scholar] [CrossRef]
- Li, K.; Liu, Q.; Fang, F.; Luo, R.; Lu, Q.; Zhou, W.; Huo, S.; Cheng, P.; Liu, J.; Addy, M.; et al. Microalgae-based wastewater treatment for nutrients recovery: A review. Bioresour. Technol. 2019, 291, 121934. [Google Scholar] [CrossRef]
- Salbitani, G.; Carfagna, S. Ammonium Utilization in Microalgae: A Sustainable Method for Wastewater Treatment. Sustainability 2021, 13, 956. [Google Scholar] [CrossRef]
- Su, Y. Revisiting carbon, nitrogen, and phosphorus metabolisms in microalgae for wastewater treatment. Sci. Total Environ. 2021, 762, 144590. [Google Scholar] [CrossRef]
- You, N.; Deng, S.; Wang, C.; Ngo, H.H.; Wang, X.; Yu, H.; Tang, L.; Han, J. Review and Opinions on the Research, Development and Application of Microalgae Culture Technologies for Resource Recovery from Wastewater. Water 2023, 15, 1192. [Google Scholar] [CrossRef]
- Yu, H.; Kim, J.; Rhee, C.; Shin, J.; Shin, S.G.; Lee, C. Effects of Different pH Control Strategies on Microalgae Cultivation and Nutrient Removal from Anaerobic Digestion Effluent. Microorganisms 2022, 10, 357. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, W.; Chen, H.; Zhan, J.; He, C.; Wang, Q. Ammonium Nitrogen Tolerant Chlorella Strain Screening and Its Damaging Effects on Photosynthesis. Front. Microbiol. 2019, 9, 3250. [Google Scholar] [CrossRef]
- Xie, Y.; Zhao, X.; Chen, J.; Yang, X.; Ho, S.-H.; Wang, B.; Chang, J.-S.; Shen, Y. Enhancing cell growth and lutein productivity of Desmodesmus sp. F51 by optimal utilization of inorganic carbon sources and ammonium salt. Bioresour. Technol. 2017, 244, 664–671. [Google Scholar] [CrossRef]
- Farahin, A.W.; Natrah, I.; Nagao, N.; Yusoff, F.M.; Shariff, M.; Banerjee, S.; Katayama, T.; Nakakuni, M.; Koyama, M.; Nakasaki, K.; et al. Tolerance of Tetraselmis tetrathele to High Ammonium Nitrogen and Its Effect on Growth Rate, Carotenoid, and Fatty Acids Productivity. Front. Bioeng. Biotechnol. 2021, 9, 568776. [Google Scholar] [CrossRef]
- Nakamura, H.; Shiozaki, T.; Gonda, N.; Furuya, K.; Matsunaga, S.; Okada, S. Utilization of ammonium by the hydrocarbon-producing microalga, Botryococcus braunii Showa. Algal Res. 2017, 25, 445–451. [Google Scholar] [CrossRef]
- Ma, R.; Tao, X.; Chua, E.T.; Ho, S.-H.; Shi, X.; Liu, L.; Xie, Y.; Chen, J. Enhancing astaxanthin production in Haematococcus pluvialis QLD by a pH steady NaHCO3-CO2-C/NH4Cl-N culture system. Algal Res. 2022, 64, 102697. [Google Scholar] [CrossRef]
- Collos, Y.; Harrison, P.J. Acclimation and toxicity of high ammonium concentrations to unicellular algae. Mar. Pollut. Bull. 2014, 80, 8–23. [Google Scholar] [CrossRef]
- Miura, R.; Furuhashi, K.; Hasegawa, F.; Kaizu, Y.; Imou, K. Calcium carbonate prevents Botryococcus braunii growth inhibition caused by medium acidification. J. Appl. Phycol. 2022, 34, 177–183. [Google Scholar] [CrossRef]
- Zhou, Y.; He, Y.; Xiao, X.; Liang, Z.; Dai, J.; Wang, M.; Chen, B. A novel and efficient strategy mediated with calcium carbonate-rich sources to remove ammonium sulfate from rare earth wastewater by heterotrophic Chlorella species. Bioresour. Technol. 2022, 343, 125994. [Google Scholar] [CrossRef]
- Tang, C.-C.; Zhang, X.-Y.; Wang, R.; Wang, T.-Y.; He, Z.-W.; Wang, X.C. Calcium ions-effect on performance, growth and extracellular nature of microalgal-bacterial symbiosis system treating wastewater. Environ. Res. 2022, 207, 112228. [Google Scholar] [CrossRef]
- Yao, C.-H.; Ai, J.-N.; Cao, X.-P.; Xue, S. Salinity manipulation as an effective method for enhanced starch production in the marine microalga Tetraselmis subcordiformis. Bioresour. Technol. 2013, 146, 663–671. [Google Scholar] [CrossRef]
- Rodriguez-Sanchez, A.; Leyva-Diaz, J.C.; Gonzalez-Lopez, J.; Poyatos, J.M. Membrane bioreactor and hybrid moving bed biofilm reactor-membrane bioreactor for the treatment of variable salinity wastewater: Influence of biomass concentration and hydraulic retention time. Chem. Eng. J. 2018, 336, 102–111. [Google Scholar] [CrossRef]
- Goswami, R.K.; Agrawal, K.; Mehariya, S.; Verma, P. Current perspective on wastewater treatment using photobioreactor for Tetraselmis sp.: An emerging and foreseeable sustainable approach. Environ. Sci. Pollut. Res. 2022, 29, 61905–61937. [Google Scholar] [CrossRef] [PubMed]
- Patrinou, V.; Daskalaki, A.; Kampantais, D.; Kanakis, D.C.; Economou, C.N.; Bokas, D.; Kotzamanis, Y.; Aggelis, G.; Vayenas, D.V.; Tekerlekopoulou, A.G. Optimization of Cultivation Conditions for Tetraselmis striata and Biomass Quality Evaluation for Fish Feed Production. Water 2022, 14, 3162. [Google Scholar] [CrossRef]
- Moheimani, N.R. Inorganic carbon and pH effect on growth and lipid productivity of Tetraselmis suecica and Chlorella sp (Chlorophyta) grown outdoors in bag photobioreactors. J. Appl. Phycol. 2013, 25, 387–398. [Google Scholar] [CrossRef]
- Song, C.; Qiu, Y.; Li, S.; Liu, Z.; Chen, G.; Sun, L.; Wang, K.; Kitamura, Y. A novel concept of bicarbonate-carbon utilization via an absorption-microalgae hybrid process assisted with nutrient recycling from soybean wastewater. J. Clean. Prod. 2019, 237, 117864. [Google Scholar] [CrossRef]
- Song, C.; Qiu, Y.; Xie, M.; Qi, Y.; Li, S.; Kitamura, Y. Novel Bio-regeneration Concept via Using Rich Solution as Nutrition Resource for Microalgae Cultivation: Effect of pH and Feeding Modes. ACS Sustain. Chem. Eng. 2019, 7, 14471–14478. [Google Scholar] [CrossRef]
- Song, C.; Liu, J.; Qiu, Y.; Xie, M.; Sun, J.; Qi, Y.; Li, S.; Kitamura, Y. Bio-regeneration of different rich CO2 absorption solvent via microalgae cultivation. Bioresour. Technol. 2019, 290, 121781. [Google Scholar] [CrossRef]
- Song, C.; Liu, J.; Xie, M.; Qiu, Y.; Chen, G.; Qi, Y.; Kitamura, Y. Intensification of a novel absorption-microalgae hybrid CO2 utilization process via fed-batch mode optimization. Int. J. Greenh. Gas Control 2019, 82, 1–7. [Google Scholar] [CrossRef]
- Song, C.; Xie, M.; Qiu, Y.; Liu, Q.; Sun, L.; Wang, K.; Kansha, Y. Integration of CO2 absorption with biological transformation via using rich ammonia solution as a nutrient source for microalgae cultivation. Energy 2019, 179, 618–627. [Google Scholar] [CrossRef]
- Jiang, R.; Qin, L.; Feng, S.; Huang, D.; Wang, Z.; Zhu, S. The joint effect of ammonium and pH on the growth of Chlorella vulgaris and ammonium removal in artificial liquid digestate. Bioresour. Technol. 2021, 325, 124690. [Google Scholar] [CrossRef]
- Markou, G.; Vandamme, D.; Muylaert, K. Ammonia inhibition on Arthrospira platensis in relation to the initial biomass density and pH. Bioresour. Technol. 2014, 166, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-Y.; Pan, L.-Y.; Hong, M.-J.; Lee, A.-C. The effects of temperature on the growth of and ammonia uptake by marine microalgae. Bot. Stud. 2012, 53, 125–133. [Google Scholar]
- Khatoon, H.; Penz Penz, K.; Banerjee, S.; Redwanur Rahman, M.; Mahmud Minhaz, T.; Islam, Z.; Ara Mukta, F.; Nayma, Z.; Sultana, R.; Islam Amira, K. Immobilized Tetraselmis sp. for reducing nitrogenous and phosphorous compounds from aquaculture wastewater. Bioresour. Technol. 2021, 338, 125529. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Ai, J.; Cao, X.; Xue, S.; Zhang, W. Enhancing starch production of a marine green microalga Tetraselmis subcordiformis through nutrient limitation. Bioresour. Technol. 2012, 118, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Jiang, J.; Cao, X.; Liu, Y.; Xue, S.; Zhang, Y. Phosphorus Enhances Photosynthetic Storage Starch Production in a Green Microalga (Chlorophyta) Tetraselmis subcordiformis in Nitrogen Starvation Conditions. J. Agric. Food Chem. 2018, 66, 10777–10787. [Google Scholar] [CrossRef]
- Rausch, T. The estimation of micro-algal protein content and its meaning to the evaluation of algal biomass I. Comparison of methods for extracting protein. Hydrobiologia 1981, 78, 237–251. [Google Scholar] [CrossRef]
- Yao, C.; Wu, P.; Pan, Y.; Lu, H.; Chi, L.; Meng, Y.; Cao, X.; Xue, S.; Yang, X. Evaluation of the integrated hydrothermal carbonization-algal cultivation process for enhanced nitrogen utilization in Arthrospira platensis production. Bioresour. Technol. 2016, 216, 381–390. [Google Scholar] [CrossRef]
- Mishra, S.K.; Suh, W.I.; Farooq, W.; Moon, M.; Shrivastav, A.; Park, M.S.; Yang, J.-W. Rapid quantification of microalgal lipids in aqueous medium by a simple colorimetric method. Bioresour. Technol. 2014, 155, 330–333. [Google Scholar] [CrossRef]
- Pan, Y.; Shen, Y.; Zhang, H.; Ran, X.; Xie, T.; Zhang, Y.; Yao, C. Fine-tuned regulation of photosynthetic performance via γ-aminobutyric acid (GABA) supply coupled with high initial cell density culture for economic starch production in microalgae. Bioresour. Bioprocess. 2022, 9, 52. [Google Scholar] [CrossRef]
- Hovenkamp-Hermelink, J.; De Vries, J.; Adamse, P.; Jacobsen, E.; Witholt, B.; Feenstra, W. Rapid estimation of the amylose/amylopectin ratio in small amounts of tuber and leaf tissue of the potato. Potato Res. 1988, 31, 241–246. [Google Scholar] [CrossRef]
- Strasserf, R.J.; Srivastava, A. Polyphasic Chlorophyll a Fluorescence Transient in Plants and Cyanobacteria. Photochem. Photobiol. 2008, 61, 32–42. [Google Scholar] [CrossRef]
- Liemann, F. Contribution to the microdetermination of ammonia, urea and residual nitrogen with the indophenol blue reactions. Z. Fur Die Gesamte Exp. Med. 1964, 138, 191. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Xiang, Q.; Wei, X.; Yang, Z.; Xie, T.; Zhang, Y.; Li, D.; Pan, X.; Liu, X.; Zhang, X.; Yao, C. Acclimation to a broad range of nitrate strength on a euryhaline marine microalga Tetraselmis subcordiformis for photosynthetic nitrate removal and high-quality biomass production. Sci. Total Environ. 2021, 781, 146687. [Google Scholar] [CrossRef]
- Zhang, T.; Chi, Z.; Sheng, J. A Highly Thermosensitive and Permeable Mutant of the Marine Yeast Cryptococcus aureus G7a Potentially Useful for Single-Cell Protein Production and its Nutritive Components. Mar. Biotechnol. 2009, 11, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef]
- Qi, M.; Yao, C.; Sun, B.; Cao, X.; Fei, Q.; Liang, B.; Ran, W.; Xiang, Q.; Zhang, Y.; Lan, X. Application of an in situ CO2–bicarbonate system under nitrogen depletion to improve photosynthetic biomass and starch production and regulate amylose accumulation in a marine green microalga Tetraselmis subcordiformis. Biotechnol. Biofuels 2019, 12, 184. [Google Scholar] [CrossRef]
- Khatoon, H.; Abdu Rahman, N.; Banerjee, S.; Harun, N.; Suleiman, S.S.; Zakaria, N.H.; Lananan, F.; Abdul Hamid, S.H.; Endut, A. Effects of different salinities and pH on the growth and proximate composition of Nannochloropsis sp. and Tetraselmis sp. isolated from South China Sea cultured under control and natural condition. Int. Biodeterior. Biodegrad. 2014, 95, 11–18. [Google Scholar] [CrossRef]
- Esakkimuthu, S.; Krishnamurthy, V.; Govindarajan, R.; Swaminathan, K. Augmentation and starvation of calcium, magnesium, phosphate on lipid production of Scenedesmus obliquus. Biomass Bioenergy 2016, 88, 126–134. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Yang, W.N.; Ngo, H.H.; Guo, W.S.; Jin, P.K.; Dzakpasu, M.; Yang, S.J.; Wang, Q.; Wang, X.C.; Ao, D. Current status of urban wastewater treatment plants in China. Environ. Int. 2016, 92–93, 11–22. [Google Scholar] [CrossRef]
- Khoo, K.H.; Culberson, C.H.; Bates, R.G. Thermodynamics of the dissociation of ammonium ion in seawater from 5 to 40 °C. J. Solut. Chem. 1977, 6, 281–290. [Google Scholar] [CrossRef]
- Ran, S.; Liu, B.; Jiang, W.; Sun, Z.; Liang, J. Transcriptome analysis of Enterococcus faecalis in response to alkaline stress. Front. Microbiol. 2015, 6, 795. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Elsayed, N.; Rezaei, N.; Guo, T.; Mohebbi, S.; Zhang, Q. Wastewater-based resource recovery technologies across scale: A review. Resour. Conserv. Recycl. 2019, 145, 94–112. [Google Scholar] [CrossRef]
- Wu, Y.-H.; Yu, Y.; Hu, H.-Y.; Zhuang, L.-L. Effects of cultivation conditions on the production of soluble algal products (SAPs) of Scenedesmus sp. LX1. Algal Res. 2016, 16, 376–382. [Google Scholar] [CrossRef]
- Baroni, É.; Cao, B.; Webley, P.A.; Scales, P.J.; Martin, G.J.O. Nitrogen Availability and the Nature of Extracellular Organic Matter of Microalgae. Ind. Eng. Chem. Res. 2020, 59, 6795–6805. [Google Scholar] [CrossRef]
- González-Camejo, J.; Pachés, M.; Marín, A.; Jiménez-Benítez, A.; Seco, A.; Barat, R. Production of microalgal external organic matter in a Chlorella-dominated culture: Influence of temperature and stress factors. Environ. Sci. Water Res. Technol. 2020, 6, 1828–1841. [Google Scholar] [CrossRef]
- Zhuang, L.-L.; Wu, Y.-H.; Espinosa, V.M.D.; Zhang, T.-Y.; Dao, G.-H.; Hu, H.-Y. Soluble Algal Products (SAPs) in large scale cultivation of microalgae for biomass/bioenergy production: A review. Renew. Sustain. Energy Rev. 2016, 59, 141–148. [Google Scholar] [CrossRef]
- Chai, W.S.; Chew, C.H.; Munawaroh, H.S.H.; Ashokkumar, V.; Cheng, C.K.; Park, Y.-K.; Show, P.L. Microalgae and ammonia: A review on inter-relationship. Fuel 2021, 303, 121303. [Google Scholar] [CrossRef]
- Paliwal, C.; Mitra, M.; Bhayani, K.; Bharadwaj, S.V.V.; Ghosh, T.; Dubey, S.; Mishra, S. Abiotic stresses as tools for metabolites in microalgae. Bioresour. Technol. 2017, 244, 1216–1226. [Google Scholar] [CrossRef]
- Ran, W.; Wang, H.; Liu, Y.; Qi, M.; Xiang, Q.; Yao, C.; Zhang, Y.; Lan, X. Storage of starch and lipids in microalgae: Biosynthesis and manipulation by nutrients. Bioresour. Technol. 2019, 291, 121894. [Google Scholar] [CrossRef]
- Brányiková, I.; Maršálková, B.; Doucha, J.; Brányik, T.; Bišová, K.; Zachleder, V.; Vítová, M. Microalgae—Novel highly efficient starch producers. Biotechnol. Bioeng. 2011, 108, 766–776. [Google Scholar] [CrossRef]
- Sajjadi, B.; Chen, W.-Y.; Raman, A.A.A.; Ibrahim, S. Microalgae lipid and biomass for biofuel production: A comprehensive review on lipid enhancement strategies and their effects on fatty acid composition. Renew. Sustain. Energy Rev. 2018, 97, 200–232. [Google Scholar] [CrossRef]
- Shi, Q.; Chen, C.; He, T.; Fan, J. Circadian rhythm promotes the biomass and amylose hyperaccumulation by mixotrophic cultivation of marine microalga Platymonas helgolandica. Biotechnol. Biofuels Bioprod. 2022, 15, 75. [Google Scholar] [CrossRef]
- Zhong, Y.; Tai, L.; Blennow, A.; Ding, L.; Herburger, K.; Qu, J.; Xin, A.; Guo, D.; Hebelstrup, K.H.; Liu, X. High-amylose starch: Structure, functionality and applications. Crit. Rev. Food Sci. Nutr. 2022, 1–23, ahead-of-print. [Google Scholar] [CrossRef]
- Aikawa, S.; Ho, S.-H.; Nakanishi, A.; Chang, J.-S.; Hasunuma, T.; Kondo, A. Improving polyglucan production in cyanobacteria and microalgae via cultivation design and metabolic engineering. Biotechnol. J. 2015, 10, 886–898. [Google Scholar] [CrossRef]
- Tanadul, O.-u.-m.; VanderGheynst, J.S.; Beckles, D.M.; Powell, A.L.T.; Labavitch, J.M. The impact of elevated CO2 concentration on the quality of algal starch as a potential biofuel feedstock. Biotechnol. Bioeng. 2014, 111, 1323–1331. [Google Scholar] [CrossRef]
- Thanigaivel, S.; Priya, A.K.; Dutta, K.; Rajendran, S.; Vasseghian, Y. Engineering strategies and opportunities of next generation biofuel from microalgae: A perspective review on the potential bioenergy feedstock. Fuel 2022, 312, 122827. [Google Scholar] [CrossRef]
- Li, Q.; Deng, F.; Zeng, Y.; Li, B.; He, C.; Zhu, Y.; Zhou, X.; Zhang, Z.; Wang, L.; Tao, Y.; et al. Low Light Stress Increases Chalkiness by Disturbing Starch Synthesis and Grain Filling of Rice. Int. J. Mol. Sci. 2022, 23, 9153. [Google Scholar] [CrossRef]
- Li, H.; Gidley, M.J.; Dhital, S. High-Amylose Starches to Bridge the “Fiber Gap”: Development, Structure, and Nutritional Functionality. Compr. Rev. Food Sci. Food Saf. 2019, 18, 362–379. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Sang, C.; Turchini, G.M.; Wang, A.; Zhang, J.; Chen, N. Starch in aquafeeds: The benefits of a high amylose to amylopectin ratio and resistant starch content in diets for the carnivorous fish, largemouth bass (Micropterus salmoides). Br. J. Nutr. 2020, 124, 1145–1155. [Google Scholar] [CrossRef]
- Pereira, H.; Silva, J.; Santos, T.; Gangadhar, K.N.; Raposo, A.; Nunes, C.; Coimbra, M.A.; Gouveia, L.; Barreira, L.; Varela, J. Nutritional Potential and Toxicological Evaluation of Tetraselmis sp. CTP4 Microalgal Biomass Produced in Industrial Photobioreactors. Molecules 2019, 24, 3192. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Levine, R.L. Methionine in proteins defends against oxidative stress. FASEB J. 2009, 23, 464–472. [Google Scholar] [CrossRef]
- Mukherjee, R.; Chakraborty, R.; Dutta, A. Role of Fermentation in Improving Nutritional Quality of Soybean Meal—A Review. Asian-Australas. J. Anim. Sci. 2016, 29, 1523–1529. [Google Scholar] [CrossRef]
- Peñaflorida, V.D. An evaluation of indigenous protein sources as potential component in the diet formulation for tiger prawn, Penaeus monodon, using essential amino acid index (EAAI). Aquaculture 1989, 83, 319–330. [Google Scholar] [CrossRef]
- Wang, M.; Kuo-Dahab, W.C.; Dolan, S.; Park, C. Kinetics of nutrient removal and expression of extracellular polymeric substances of the microalgae, Chlorella sp. and Micractinium sp., in wastewater treatment. Bioresour. Technol. 2014, 154, 131–137. [Google Scholar] [CrossRef]
- Molaei, S.; Moussavi, G.; Talebbeydokhti, N.; Shekoohiyan, S. Biodegradation of the petroleum hydrocarbons using an anoxic packed-bed biofilm reactor with in-situ biosurfactant-producing bacteria. J. Hazard. Mater. 2022, 421, 126699. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae Strain | Water Source | NH4+-N (mg/L) | Carbon Source | Air + CO2 | NH4+-N Removal Efficiency (%) (Day) | Maximum NH4+-N Removal Capacity (mg N/L/Day) | NH3 Escaped (%) | Extra pH Control | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Chlorella sp. L38 | Freshwater | 248 | NH4HCO3 | Air | 80 (24) | 8.67 | NA a | No | [25] |
| 370 | NH4HCO3 | Air | 55 (27) | 10 | 19 b | Acid/base | [26] | ||
| 280 | NH4HCO3 | Air | 45 (15) | 8.31 | 21.43 | Acid/base | [27] | ||
| 248 | NH4HCO3 | Air | 44 (27) | 12 | 24.2 b | Acid/base | [28] | ||
| Chlorella sp. L166 | Freshwater | 247 | NH4HCO3 | Air | 87 (36) | 13 | 76 | No | [29] |
| 247 | NH4HCO3 | Air + 5% | 52 (18) | 10 | 42 | No | |||
| 247 | NH4HCO3 | Air | 84 (36) | 9 | 73 b | No | |||
| Chlorella vulgaris | Freshwater | 120 | NH4HCO3 | Air + 1% | 100 (3) | 53.4 | NA | Acid/base | [30] |
| Chlorella strains | Freshwater | 50 | Na2CO3 | Air | 100 (4) | 12.5 | NA | No | [11] |
| Haematococcus pluvialis QLD | Freshwater | 63 | NaHCO3 | Air + 1% | 95.6 (5) | 15.75 | NA | No | [15] |
| Desmodesmus sp. F51 | Freshwater | 60.2 | NaHCO3 | Air + 2.5% | 100 (1.1) | 55.2 | NA | No | [12] |
| 60.2 | No | Air + 2.5% | 100 (1.3) | 50.9 | NA | Acid/base | |||
| 60.2 | NaHCO3 | Air | 25.25 (2) | 20 | NA | No | |||
| Botryococcus braunii | Freshwater | 83.15 | CaCO3 | Air | 68.55 (20) | 2.85 | NA | No | [17] |
| Arthrospira platensis | Semi-seawater | 100 | NaHCO3 | Air | 100 (2) | 50 | 23~40 | No | [31] |
| Tetraselmis chui | Seawater | 13 | NaHCO3 | Air | 73 (1) | 9.5 | NA | No | [32] |
| Nannochloropsis oculata | Seawater | 13 | NaHCO3 | Air | 32 (1) | 4.2 | NA | No | [32] |
| Tetraselmis sp. | Seawater | 3.8 | No | Air | 100 (1) | 3.8 c | NA | No | [33] |
| Tetraselmis subcordiformis | Seawater | 100 | NaHCO3 | Air | 99.6 (4) | 49.27 | 48.60 | No | This study |
| 100 | MgCO3 | Air | 99.6 (4) | 44.00 | 43.70 | No | |||
| 100 | CaCO3 | Air | 99.3 (4) | 31.69 | 38.93 | No | |||
| 100 | NaHCO3 | Air + 2% | 99.5 (3) | 60.13 | 19.65 | No | |||
| 100 | MgCO3 | Air + 2% | 99.5 (2) | 54.38 | 20.33 | No | |||
| 100 | CaCO3 | Air + 2% | 99.8 (3) | 45.95 | 26.86 | No |
| Culture System | Bicarbonate/Carbonate Supply | Am (% of Total Starch) | Ap (% of Total Starch) | Am/Ap |
|---|---|---|---|---|
| IC | NaHCO3 | 39.46 ± 0.01 | 60.54 ± 0.01 | 0.65 ± 0.03 |
| MgCO3 | 41.76 ± 0.01 | 58.24 ± 0.01 | 0.72 ± 0.03 | |
| CaCO3 | 37.10 ± 0.02 | 62.90 ± 0.02 | 0.59 ± 0.05 | |
| CO2-IC | NaHCO3 | 38.90 ± 0.01 | 61.10 ± 0.01 | 0.64 ± 0.02 |
| MgCO3 | 59.57 ± 0.00 * | 40.43 ± 0.00 * | 1.47 ± 0.00 * | |
| CaCO3 | 50.54 ± 0.00 * | 49.46 ± 0.00 * | 1.02 ± 0.01 * |
| Animo Acids | IC | CO2-IC | Soybean Meal c | Fish Meal c | Penaeus monodon Juvenile c | Ruminant Diets c | Ideal Protein for Dairy Cow c | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| NaHCO3 | MgCO3 | CaCO3 | NaHCO3 | MgCO3 | CaCO3 | ||||||
| Essential AA (EAA) | |||||||||||
| Arginine | 4.65 ± 0.21 | 4.35 ± 0.12 | 4.92 ± 0.65 | 4.99 ± 0.26 | 4.50 ± 0.05 | 4.94 ± 0.17 | 8.30 | 6.75 | 8.00 | 14.8 | |
| Histidine | 1.69 ± 0.06 | 1.94 ± 0.03 | 1.75 ± 0.02 | 2.23 ± 0.19 | 1.88 ± 0.04 | 2.03 ± 0.15 | 3.04 | 2.54 | 2.48 | 4.8 | |
| Isoleucine | 4.22 ± 0.30 | 4.32 ± 0.09 | 4.39 ± 0.17 | 4.34 ± 0.12 | 4.49 ± 0.24 | 4.30 ± 0.01 | 5.22 | 4.32 | 4.46 | 3.6 | 4.8 |
| Leucine | 9.12 ± 0.23 | 9.16 ± 0.08 | 9.13 ± 0.27 | 9.16 ± 0.05 | 9.40 ± 0.23 | 8.91 ± 0.33 | 8.30 | 8.06 | 7.66 | 6.0 | 8.6 |
| Lysine | 7.39 ± 0.13 | 7.15 ± 0.01 | 7.45 ± 0.06 | 8.54 ± 0.57 | 8.31 ± 0.35 | 8.56 ± 0.28 | 6.95 | 8.76 | 7.58 | 7.6 | 6.7 |
| Methionine | 1.05 ± 0.62 | 2.14 ± 0.01 | 1.19 ± 0.07 | 0.92 ± 0.14 | 0.43 ± 0.20 | 1.45 ± 0.25 | 1.97 | 3.23 | 2.85 | 1.3 | 2.0 |
| Phenylalanine | 5.94 ± 0.01 | 5.86 ± 0.05 | 5.72 ± 0.12 | 5.71 ± 0.08 | 6.10 ± 0.23 | 5.75 ± 0.17 | 4.88 | 4.21 | 4.20 | 2.9 | 4.4 |
| Threonine | 6.66 ± 0.04 | 6.78 ± 0.33 | 6.33 ± 0.06 | 6.14 ± 0.06 | 6.15 ± 0.29 | 6.08 ± 0.04 | 4.09 | 5.12 | 3.96 | 3.8 | 5.2 |
| Tryptophan a | - | - | - | - | - | - | |||||
| Valine | 6.56 ± 0.18 | 6.67 ± 0.02 | 6.63 ± 0.04 | 6.25 ± 0.67 | 6.71 ± 0.12 | 6.30 ± 0.03 | 3.85 | 5.07 | 5.16 | 5.1 | 5.3 |
| TOTAL EAA | 47.27 | 48.39 | 47.51 | 48.05 | 48.54 | 48.69 | 46.60 | 48.06 | 46.34 | ||
| Non-essential AA (NEAA) | |||||||||||
| Alanine | 8.16 ± 0.03 | 7.89 ± 0.04 | 8.15 ± 0.08 | 7.31 ± 0.00 | 7.29 ± 0.45 | 7.75 ± 0.23 | 4.61 | 6.45 | 6.11 | ||
| Aspartic acid b | 11.40 ± 0.34 | 11.24 ± 0.56 | 11.17 ± 0.39 | 11.85 ± 0.47 | 11.09 ± 1.32 | 11.15 ± 0.26 | 11.91 | 10.63 | 10.20 | ||
| Cysteine | 0.60 ± 0.06 | 0.63 ± 0.16 | 0.91 ± 0.11 | 0.60 ± 0.10 | 0.91 ± 0.23 | 0.63 ± 0.10 | 2.20 | 1.08 | 1.03 | ||
| Glutamic acid b | 13.74 ± 0.45 | 12.92 ± 0.30 | 13.94 ± 0.09 | 13.52 ± 0.22 | 13.18 ± 1.27 | 13.00 ± 0.33 | 18.07 | 15.38 | 16.07 | ||
| Glycine | 5.48 ± 00.35 | 5.51 ± 0.03 | 5.33 ± 0.06 | 5.23 ± 0.20 | 6.07 ± 0.54 | 5.70 ± 0.10 | 3.99 | 5.68 | 8.12 | ||
| Proline | 4.96 ± 0.15 | 5.00 ± 0.03 | 4.98 ± 0.21 | 4.83 ± 0.08 | 4.97 ± 0.59 | 5.02 ± 0.32 | 4.37 | 4.21 | 4.02 | ||
| Serine | 5.38 ± 0.13 | 5.45 ± 0.33 | 5.13 ± 0.23 | 5.16 ± 0.24 | 5.04 ± 0.11 | 5.18 ± 0.07 | 5.04 | 5.10 | 4.16 | ||
| Tyrosine | 3.00 ± 0.14 | 2.97 ± 0.02 | 2.87 ± 0.14 | 3.21 ± 0.15 | 3.46 ± 0.35 | 3.25 ± 0.08 | 3.22 | 3.41 | 3.94 | ||
| EAAI for prawn | 0.90 | 0.97 | 0.92 | 0.91 | 0.83 | 0.94 | 0.98 | 1.01 | |||
| EAAI for ruminant | 1.01 | 1.09 | 1.03 | 1.02 | 0.93 | 1.06 | 0.97 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, Y.; Liao, L.; Wu, W.; Zhang, H.; Ran, X.; Xie, T.; Zhang, Y.; Yao, C. CO2-Inorganic Carbon Auto-Buffering System for Efficient Ammonium Reclamation Coupled with Valuable Biomass Production in a Euryhaline Microalga Tetraselmis subcordiformis. Water 2023, 15, 1671. https://doi.org/10.3390/w15091671
Shen Y, Liao L, Wu W, Zhang H, Ran X, Xie T, Zhang Y, Yao C. CO2-Inorganic Carbon Auto-Buffering System for Efficient Ammonium Reclamation Coupled with Valuable Biomass Production in a Euryhaline Microalga Tetraselmis subcordiformis. Water. 2023; 15(9):1671. https://doi.org/10.3390/w15091671
Chicago/Turabian StyleShen, Yuhan, Longren Liao, Weidong Wu, Haoyu Zhang, Xiuyuan Ran, Tonghui Xie, Yongkui Zhang, and Changhong Yao. 2023. "CO2-Inorganic Carbon Auto-Buffering System for Efficient Ammonium Reclamation Coupled with Valuable Biomass Production in a Euryhaline Microalga Tetraselmis subcordiformis" Water 15, no. 9: 1671. https://doi.org/10.3390/w15091671