
Elemental Fractionation in Sabellariidae (Polychaeta) Biocement and Comparison with Seawater Pattern: A New Environmental Proxy in a High-Biodiversity Ecosystem?


 ,
, 
 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
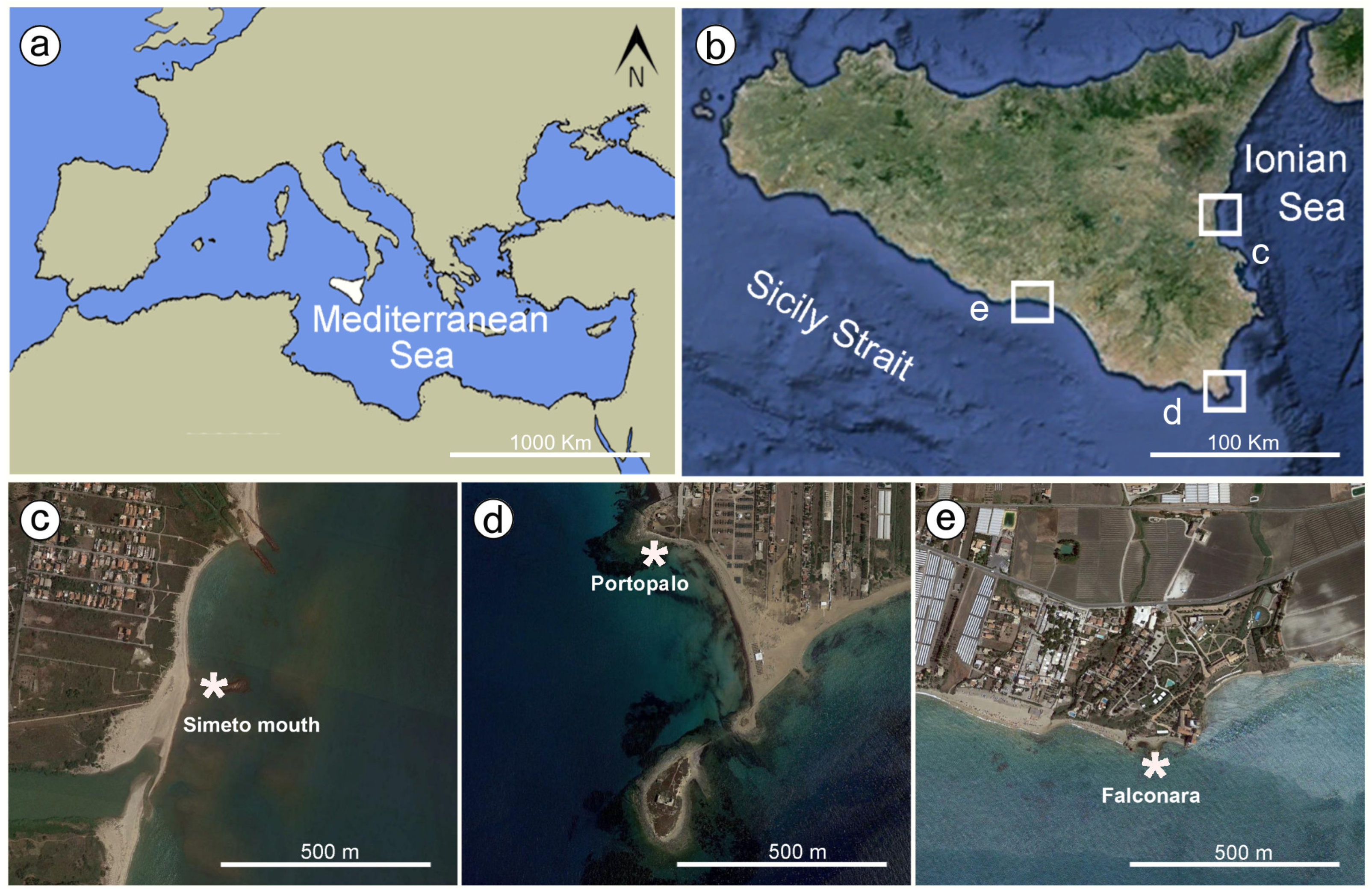
2. Study Areas
3. Materials and Methods
4. Results and Discussion
4.1. Biocement: Morphological and Chemical Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | N | O | Na | Mg | Al | Si | P | S | Cl | K | Ca | Fe | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| wt% | wt% | wt% | wt% | wt% | wt% | wt% | wt% | wt% | wt% | wt% | wt% | wt% | |
| Portopalo | 42.80 | 12.18 | 21.70 | 3.25 | 3.10 | 0.26 | 0.38 | 2.86 | 0.80 | 4.90 | 0.20 | 7.12 | 0.06 |
| Simeto | 44.70 | 13.08 | 22.14 | 1.52 | 3.22 | 0.11 | 0.31 | 6.34 | 0.79 | 2.79 | 0.12 | 4.70 | 0.02 |
| Falconara | 38.31 | 10.97 | 28.62 | 1.95 | 1.99 | 1.41 | 3.08 | 4.03 | 0.66 | 2.17 | 0.27 | 5.46 | 0.74 |
| B | Sr | Cu | Zn | U | Cr | Ni | |
|---|---|---|---|---|---|---|---|
| wt% | wt% | wt% | wt% | wt% | wt% | wt% | |
| Portopalo | 0.299 | 0.016 | 0.006 | 0.018 | 0.038 | 0.002 | 0.012 |
| Simeto | 0.103 | 0.018 | 0.010 | 0.014 | 0.023 | 0.004 | 0.003 |
| Falconara | 0.308 | 0.011 | 0.009 | 0.008 | 0.010 | 0.005 | 0.011 |
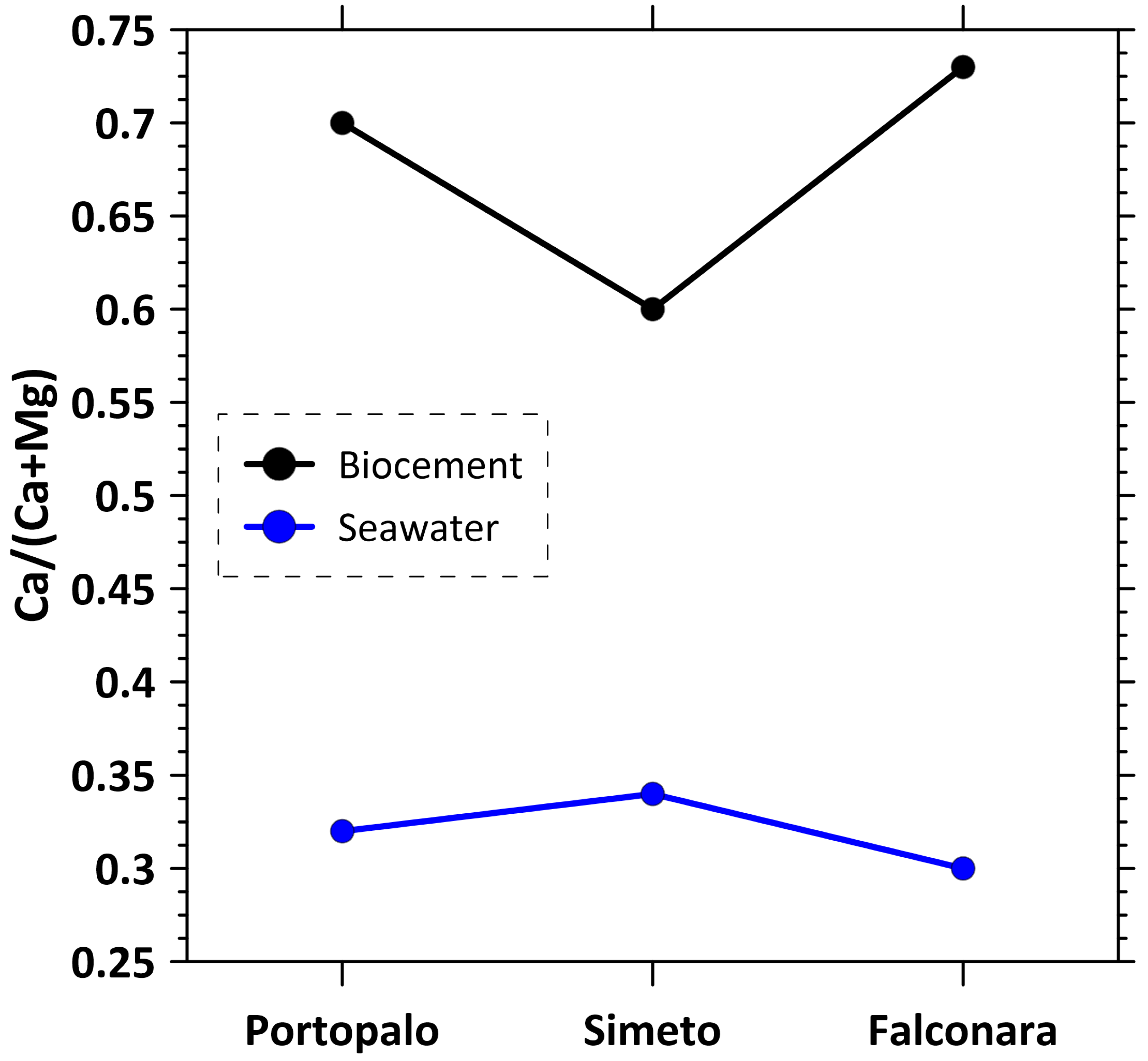
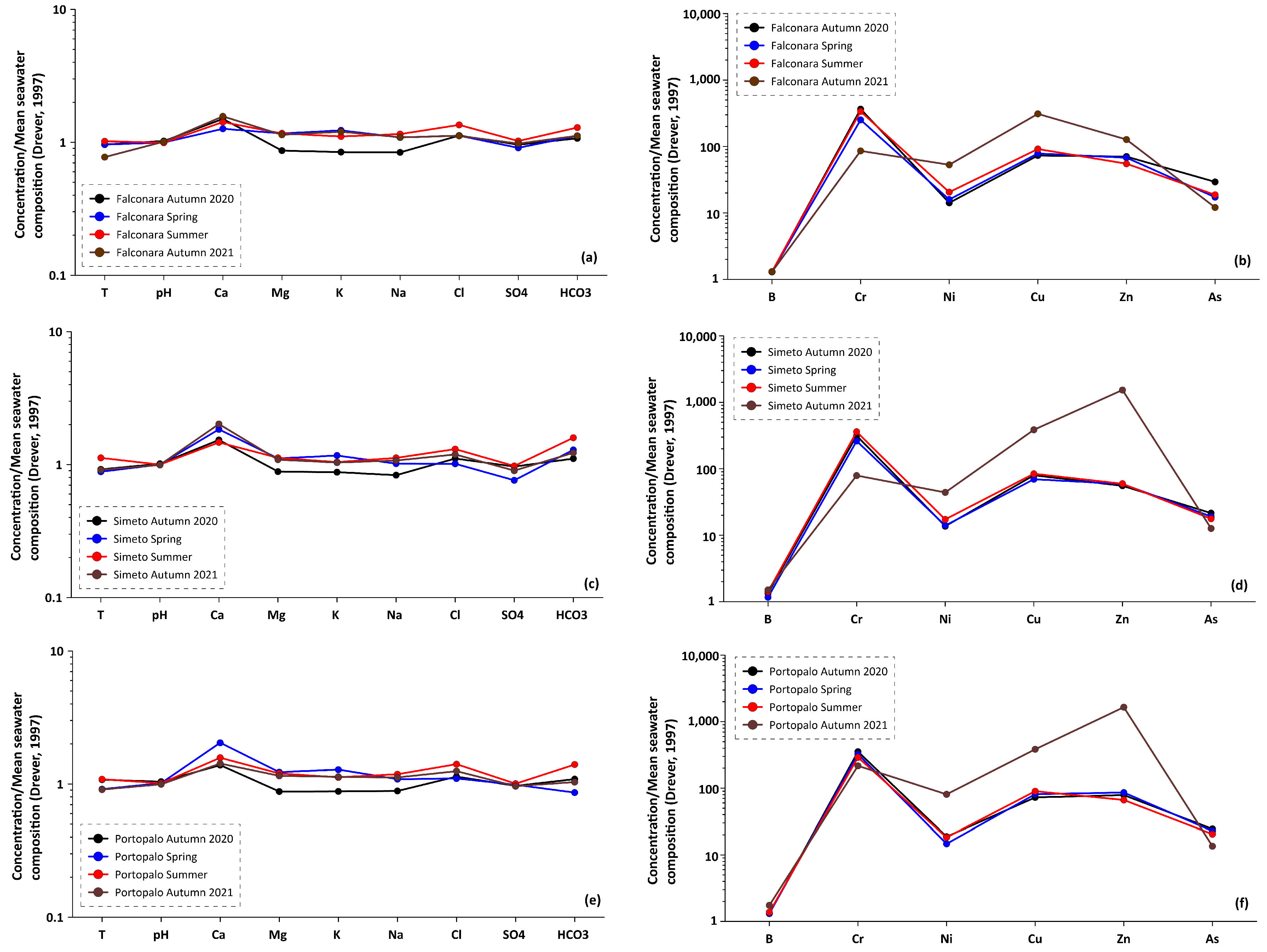
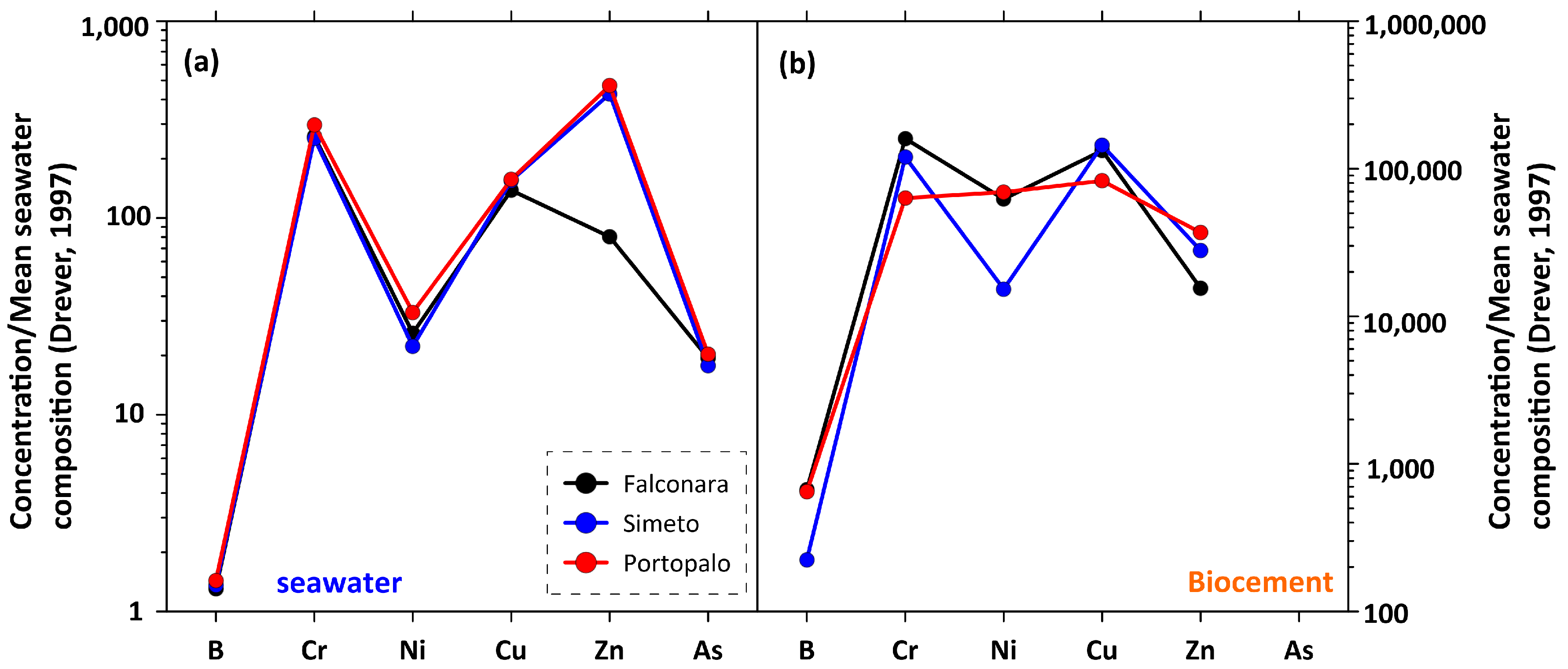
4.2. Local Seawater Composition and Comparison with Biocement
4.3. Sabellariid Biocement as a New Environmental Proxy
4.4. Ecosystem Biodiversity
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linnaeus, C. Systema Naturae, per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species, Cum Caracteribus, Differentiis, Synonymis, Locis; Editio duodecima, reformata; L. Salvii: Holmiae, Sweden, 1767; Volume I, Part 2; pp. 533–1327. [Google Scholar]
- Vovelle, J. Le tube de Sabellaria alveolata (L.) Annelide Polychete Hermellidae et son ciment. Etude ecologique, experimentale, histologique et histochimique. Arch. Zool. Exp. Gen. 1965, 106, 1–187. [Google Scholar]
- Gruet, Y.; Vovelle, J.; Grasset, M. Composante biominerale du ciment du tube ches Sabellaria alveolata (L.) Annelide Polychete. Can. J. Zool. 1987, 65, 837–842. [Google Scholar] [CrossRef]
- Waite, J.H.; Jensen, R.A.; Morse, D.E. Cement precursor proteins of the reef–building polychaete Phragmatopoma californica (Fewkes). Biochemistry 1992, 31, 5733–5738. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Sun, C.; Stewart, J.; Waite, J. Cement proteins of the tube–building polychaete Phragmatopoma californica. J. Biol. Chem. 2005, 280, 42938–42944. [Google Scholar] [CrossRef]
- Gruet, Y. Aspects morphologiques et dynamiques de constructions de l’Annelide polychete Sabellaria alveolata (Linnaeus). Rev. Trav. Inst. Peches Marit. 1972, 36, 131–161. [Google Scholar]
- Gruet, Y. Recherches sur l’écologie des” récifs” d’hermelles édifiés par l’Annélide Polychète Sabellaria alveolata (Linnaeus). Ph.D. Thesis, Université de Nantes, Nantes, France, 1982. [Google Scholar]
- Naylor, L.A.; Viles, H.A. A temperate reef builder: An evaluation of the growth, morphology and composition of Sabellaria alveolata (L.) colonies on carbonate platforms in South Wales. Geol. Soc. London Spéc. Publ. 2000, 178, 9–19. [Google Scholar] [CrossRef]
- Delbono, I.; Bianchi, C.N.; Morri, C. Le biocostruzioni di Sabellaria alveolata come indicatori ambientali: Area costiera fra Chiavari e Sestri Levante. In Studi per la Creazione di Strumenti di Gestione costiera: Golfo del Tigullio; Ferretti, O., Ed.; ENEA, Centro Ricerche Ambiente Marino: La Spezia, Italy, 2003; pp. 130–140. [Google Scholar]
- Wilson, D. Sabellaria colonies at Duckpool, North Cornwall, 1961–1970. J. Mar. Biol. Assoc. U. K. 1971, 51, 509–580. [Google Scholar] [CrossRef]
- Cunningham, P.N.; Hawkins, S.J.; Jones, H.D.; Burrows, M.T. The geographical distribution of Sabellaria alveolata (L.) in England, Wales and Scotland, with investigations into the community structure of, and the effects of trampling on Sabellaria alveolata colonies. In Report to the Nature Conservancy Council from the Department of Zoology; Manchester University: Manchester, UK, 1984; pp. 1–38. [Google Scholar]
- Dias, A.S.; Paula, J. Associated fauna of Sabellaria alveolata colonies on the central coast of Portugal. J. Mar. Biolog. Assoc. 2001, 80, 169–170. [Google Scholar] [CrossRef]
- Dubois, S.; Retière, C.; Olivier, F. Biodiversity associated with Sabellaria alveolata (Polychaeta: Sabellariidae) reefs: Effects of human disturbances. J. Mar. Biol. Assoc. U. K. 2002, 82, 817–826. [Google Scholar] [CrossRef]
- Dubois, S.; Barille, L.; Retiere, C. Efficiency of particle retention and clearance rate in the polychaete Sabellaria alveolata L. Comptes Rendus Biol. 2003, 326, 413–421. [Google Scholar] [CrossRef]
- Dubois, S.; Commito, J.A.; Olivier, F.; Retière, C. Effects of epibionts on Sabellaria alveolata (L.) biogenic reefs and their associated fauna in the Bay of Mont Saint–Michel. Estuar. Coast. Shelf Sci. 2006, 68, 635–646. [Google Scholar] [CrossRef]
- Firth, L.B.; Mieszkowska, N.; Grant, L.M.; Bush, L.E.; Davies, A.J.; Frost, M.T.; Hawkins, S.J. Historical comparisons reveal multiple drivers of decadal change of an ecosystem engineer at the range edge. Ecol. Evol. 2015, 5, 3210–3222. [Google Scholar] [CrossRef]
- Schlund, E.; Basuyaux, O.; Lecornu, B.; Pezy, J.P.; Baffreau, A.; Dauvin, J.C. Macrofauna associated with temporary Sabellaria alveolata reefs on the west coast of Cotentin (France). SpringerPlus 2016, 5, 1260. [Google Scholar] [CrossRef]
- Holt, T.J.; Rees, E.I.; Hawkins, S.J.; Seed, R. Biogenic Reefs: An overview of dynamic and sensitivity characteristics for conservation management of marine SACs; UK Marine SACs Project. Scott. Assoc. Mar. Sci. 1998, IX, 170. [Google Scholar]
- Desroy, N.; Dubois, S.F.; Fournier, J.; Ricquiers, I.; Le Mao, P.; Guerine, L.; Gerla, D.; Rougerie, M.; Legendre, A. The conservation status of Sabellaria alveolata (L.) (Polychaeta: Sabellariidae) reefs in the Bay of Mont Saint Michel. Aquat. Conserv. Mar. Freshw. Ecosyst. 2011, 21, 462–471. [Google Scholar] [CrossRef]
- Le Cam, J.B.; Fournier, J.; Etienne, S.; Couden, J. The strength of biogenic sand reefs: Visco–elastic behaviour of cement secreted by the tube building polychaete Sabellaria alveolata, Linnaeus, 1767. Coast. Shelf Sci. 2011, 91, 333–339. [Google Scholar] [CrossRef]
- Nicoletti, L.; Lattanzi, L.; La Porta, B.; La Valle, P.; Gambi, M.C.; Tomassetti, P.; Chimenz Gusso, C. Biocostruzioni a Sabellaria delle coste del Lazio (Tirreno centrale). Biol. Mar. Mediterr. 2001, 8, 252–258. [Google Scholar]
- La Porta, B.; La Valle, P.; Chimenz Gusso, C. Sabellaria alveolata (L.) (Polychaeta Sabellariidae): La selezione dei granuli di sedimento per la costruzione dei tubi. Biol. Mar. Mediterr. 2006, 13, 593–596. [Google Scholar]
- Schimmenti, E.; Musco, L.; Lo Brutto, S.; Mikac, B.; Nygren, A.; Badalamenti, F. Mediterranean record of Eulalia ornata (Annelida: Phyllodocida) corroborating its fidelity link with the Sabellaria alveolata reef habitat. Mediterr. Mar. Sci. 2016, 17, 359–370. [Google Scholar] [CrossRef]
- Bertocci, I.; Badalamenti, F.; Lo Brutto, S.; Mikac, B.; Pipitone, C.; Schimmenti, E.; Fernández, T.V.; Musco, L. Reducing the data–deficiency of threatened European habitats: Spatial variation of Sabellariid worm reefs and associated fauna in the Sicily Channel, Mediterranean Sea. Mar. Environ. Res. 2017, 130, 325–337. [Google Scholar] [CrossRef]
- Sanfilippo, R.; Guido, A.; Insacco, G.; Deias, C.; Catania, G.; Reitano, A.; Leonardi, R.; Rosso, A. Distribution of Sabellaria alveolata (Polychaeta Sabellariidae) in the Mediterranean Sea: Update and new findings. Zoosymposia 2020, 19, 198–208. [Google Scholar] [CrossRef]
- Bonifazi, A.; Lezzi, M.; Ventura, D.; Lisco, S.; Cardone, F.; Gravina, M.F. Macrofaunal biodiversity associated with different developmental phases of a threatened Mediterranean Sabellaria alveolata (Linnaeus, 1767) reef. Mar. Environ. Res. 2019, 145, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Borghese, J.; Arduini, D.; Schimmenti, E.; Iaciofano, D.; Mikac, B.; Badalamenti, F.; Giangrande, A.; Gravina, M.F.; Musco, L.; Lo Brutto, S. Assessment of the Sabellaria alveolata reefs’ structural features along the Southern coast of Sicily (Strait of Sicily, Mediterranean Sea). Mediterr. Mar. Sci. 2022, 23, 890–899. [Google Scholar] [CrossRef]
- Ingrosso, G.; Abbiati, M.; Badaloamenti, F.; Bavestrello, G.; Belmonte, G.; Cannas, R.; Benedetti-Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Bianchi, C.N.; et al. Mediterranean Bioconstructions Along the Italian Coast. Adv Mar Biol. 2018, 79, 61–136. [Google Scholar]
- Kirtley, D.W. Worm reefs as related to beach stabilization. Shore Beach 1967, 35, 31–34. [Google Scholar]
- Bruschetti, M. Role of reef–building, ecosystem engineering polychaetes in shallow water ecosystems. Diversity 2019, 11, 168. [Google Scholar] [CrossRef]
- Plicanti, A.; Domínguez, R.; Dubois, S.F.; Bertocci, I. Human impacts on biogenic habitats: Effects of experimental trampling on Sabellaria alveolata (Linnaeus, 1767) reefs. J. Exp. Mar. Biol. Ecol. 2016, 478, 34–44. [Google Scholar] [CrossRef]
- Muller, R.A.; Stone, G.W. A climatology of tropical storm and hurricane strikes to enhance vulnerability prediction for the southeast U.S. coast. J. Coast. Res. 2001, 17, 949–956. [Google Scholar]
- Wolff, N.H.; Wong, A.; Vitolo, R.; Stolberg, K.; Anthony, K.R.N.; Mumby, P.J. Temporal clustering of tropical cyclones on the Great Barrier Reef and its ecological importance. Coral Reefs 2016, 35, 613–623. [Google Scholar] [CrossRef]
- Vorberg, R. Effects of shrimp fisheries on reefs of Sabellaria spinulosa (Polychaeta). ICES J. Mar. Sci. 2000, 57, 1416–1420. [Google Scholar] [CrossRef]
- Plicanti, A.; Iaciofano, D.; Bertocci, I.; Lo Brutto, S. The amphipod assemblages of Sabellaria alveolata reefs from the NW coast of Portugal: An account of the present knowledge, new records, and some biogeographic considerations. Mar. Biodivers. 2016, 47, 521–534. [Google Scholar] [CrossRef]
- Jones, A.G.; Dubois, S.F.; Desroy, N.; Fournier, J. Interplay between abiotic factors and species assemblages mediated by the ecosystem engineer Sabellaria alveolata (Annelida: Polychaeta). Estuar. Coast. Shelf Sci. 2018, 200, 1–18. [Google Scholar] [CrossRef]
- Gubbay, S.; Sanders, N.; Haynes, T.; Janssen, J.R.; Rodwell, J.A.M.; Nieto, M.; García Criado, M.; Beal, S.; Borg, J.; Kennedy, M.; et al. European Red List of Habitats. Marine Habitats; European Union Publication Offices: Luxembourg, 2016; Volume 1, p. 48. [Google Scholar]
- Coclet, C.; Garnier, C.; D’Onofrio, S.; Durrieu, G.; Pasero, E.; Le Poupon, C.; Omanović, D.; Mullot, J.U.; Misson, B.; Briand, J.F. Trace metal contamination impacts predicted functions more than structure of marine prokaryotic biofilm communities in an anthropized coastal area. Front. Microbiol. 2021, 12, 589948. [Google Scholar] [CrossRef]
- Little, C.; Williams, G.A.; Trowbridge, C.D. The Biology of Rocky Shores; Oxford University Press: Oxford, England, 2009; p. 376. [Google Scholar]
- Gillet, P. Bioaccumulation du cuivre et du zinc chez Nereis diversicolor (Annélide Polychète) de l’estuaire du Bou Regreg (Maroc). Cah. Biol. Mar. 1987, 28, 339–350. [Google Scholar]
- Cheggour, M.; Texier, H.; Moguedet, G.; Elkaïm, B. Metal exchange in the fauna–sediment system. The case of Nereis diversicolor and Scrobicularia plana in the Bou Regreg estuary (Morocco). Hydrobiologia 1990, 207, 209–219. [Google Scholar] [CrossRef]
- Ait Alla, A.; Gillet, P.; Deutsch, B.; Moukrim, A.; Bergayou, H. Response of Nereis diversicolor (Polychaeta, Nereididae) populations to reduced wastewater discharge in the polluted estuary of Oued Souss, Bay of Agadir, Morocco Estuar. Coast. Shelf Sci. 2006, 70, 633–642. [Google Scholar] [CrossRef]
- Ferssiwi, A. Accumulation du Cadmium, Cuivre et zinc dans le Sédiment et chez Quatre espèces D’annélides Polychètes du Littoral d’El Jadida (Côte Atlantique Marocaine): Implication des Proteins Type Metallothioneines. Ph.D. Thesis, University of Chouaïb Doukkali, El Jadida, Morocco, 2007. [Google Scholar]
- Dean, H.K. The use of polychaetes (Annelida) as indicator species of marine pollution: A review. Rev. Biol. Trop. 2008, 56, 11–38. [Google Scholar]
- Rouhi, A.; Sif, J.; Chemaa, A. Evaluation de la pollution métallique du littoral de la ville d’El Jadida (Maroc) utilisation de l’annélide Arenicola grubii comme indicateur biologique. Bull. Inst. Sci. Rabat. Sect. Sci. Vie. 2012, 34, 163–171. [Google Scholar]
- Idardare, Z.; Moukrim, A.; Chiffoleau, J.F.; Ait Alla, A.; Auger, D.; Rozuel, E. Evaluation de la contamination métallique dans deux lagunes marocaines: Khnifiss et Oualidia. Rev. Mar. Sci. Agron. Vét. 2013, 2, 58–67. [Google Scholar]
- Pini, J.M.; Richir, J.; Watson, G.J. Metal bioavailability and bioaccumulation in the polychaete Nereis (Alitta) virens (Sars): The effects of site–specific sediment characteristics. Mar. Pollut. Bull. 2015, 95, 565–575. [Google Scholar] [CrossRef]
- Rouhi, A.; Sif, J.; Ferssiwi, A.; Chemaa, A. Bioaccumulation de quelques éléments métalliques par deux espèces d’Annélides Polychètes du littoral de Jorf Lasfar (Région d’El Jadida, Maroc). Bull. Inst. Sci. Rabat. Sect. Sci. Vie. 2007, 29, 81–87. [Google Scholar]
- Chouikh, N.; Gillet, P.; Langston, W.J.; Cheggour, M.; Maarouf, A.; El Hachimi, Y.; Mouabad, A. Spatial and temporal assessment of metals contamination in the surface sediments of biogenic intertidal reefs of Sabellaria alveolata (Annelida: Polychaeta) from Essaouira protected coastal area (Atlantic coast of Morocco). Reg. Stud. Mar. Sci. 2021, 48, 101998. [Google Scholar] [CrossRef]
- Chouikh, N.; Gillet, P.; Cheggour, M.; Maarouf, A.; EL Hachimi, Y.; Mounir, A.; Alahyane, H.; Mouabad, A. Bioaccumulation of trace metals in Sabellaria alveolata (Annelida: Polychaeta) and their controlling factors along the Essaouira Protected Coastal Area (Atlantic Coast of Morocco). Ocean Sci. J. 2022, 57, 224–238. [Google Scholar] [CrossRef]
- Flammang, P.; Lambert, A. Cement ultrastructure and adhesive glands morphology in the tube–dwelling polychaete Sabellaria alveolata. J. Morphol. 2008, 269, 1479–11479. [Google Scholar]
- Fournier, J.; Etienne, S.; Le Cam, J.B. Inter– and intraspecific variability in the chemical composition of the mineral phase of cements from several tube–building polychaetes. Geobios 2010, 43, 191–200. [Google Scholar] [CrossRef]
- Sanfilippo, R.; Rosso, A.; Mastandrea, A.; Viola, A.; Deias, C.; Guido, A. Sabellaria alveolata sandcastle worm from the Mediterranean Sea: New insights on tube architecture and biocement. J. Morphol. 2019, 280, 1839–1849. [Google Scholar] [CrossRef]
- Stewart, R.J.; Weaver, J.C.; Morse, D.E.; Waite, J.H. The tube cement of Phragmatopoma californica: A solid foam. J. Exp. Biol. 2004, 207, 4727–4734. [Google Scholar] [CrossRef]
- Stewart, R.J.; Wang, C.S.; Song, I.T.; Jones, J.P. The role of coacervation and phase transitions in the sandcastle worm adhesive system. Adv. J. Colloid Interface Sci. 2017, 239, 88–96. [Google Scholar] [CrossRef]
- Stevens, M.J.; Steren, R.E.; Hlady, V.; Stewart, R.J. Multiscale structure of the underwater adhesive of Phragmatopoma californica: A nanostructured latex with a steep microporosity gradient. Langmuir 2007, 23, 5045–5049. [Google Scholar] [CrossRef]
- Sun, C.J.; Fantner, G.E.; Adams, J.; Hansma, P.K.; Waite, J.H. The role of calcium and magnesium in the concrete tubes of the sandcastle worm. J. Exp. Biol. 2007, 210, 1481–1488. [Google Scholar] [CrossRef]
- Wang, C.S.; Svendsen, K.K.; Stewart, R.J. Morphology of the adhesive system in the sandcastle worm, Phragmatopoma californica. In Biological Adhesive Systems: From Nature to Technical and Medical Applications; von Byern, J., Grunwald, I., Eds.; Springer: Wien, NY, USA, 2010; pp. 1–11. [Google Scholar]
- Wang, C.S.; Stewart, R.J. Localization of the bioadhesive precursors of the sandcastle worm, Phragmatopoma californica (Fewkes). J. Exp. Biol. 2012, 215, 351–361. [Google Scholar] [CrossRef]
- Langston, W.J.; Spence, S.K. Biological factors involved in metal concentrations observed in aquatic organisms. In Metal Speciation and Bioavailability in Aquatic System; IU–PAC; Teissier, A., Turner, D.R., Eds.; John Wiley and Sons: Chichester, UK; London, UK, 1995; Volume 3, pp. 407–478. [Google Scholar]
- Batten, S.D.; Bamber, R.N. The effects of acidified seawater on the polychaete Nereis virens (Sars, 1835). Mar. Pollut. Bull. 1996, 32, 283–287. [Google Scholar] [CrossRef]
- Amiard, J.C.; Geffard, A.; Amiard–Triquet, C. La métallothionéine chez la moule Mytilus edulis comme biomarqueur de pollution métallique: Variabilité entre sites, saisons et organes. J. Rech. Oceangr. 1998, 23, 25–30. [Google Scholar]
- Fattorini, D.; Notti, A.; Di Mento, R.; Cicero, A.M.; Gabellini, M.; Russo, A.; Regoli, F. Seasonal, spatial and inter–annual variations of trace metals in mussels from the Adriatic Sea: A regional gradient for arsenic and implications for monitoring the impact of offshore activities. Chemosphere 2008, 72, 1524–1533. [Google Scholar] [CrossRef]
- Coccioni, R.; Frontalini, F.; Marsili, A.; Mana, D. Benthic foraminifera and trace element distribution: A case–study from the heavily polluted lagoon of Venice (Italy). Mar. Pollut. Bull. 2009, 59, 257–267. [Google Scholar] [CrossRef]
- Frontalini, F.; Coccioni, R.; Bucci, C. Benthic foraminiferal assemblages and trace element contents from the lagoons of Orbetello and Lesina. Environ. Monit. Assess. 2010, 170, 245–260. [Google Scholar] [CrossRef]
- Stanković, S.; Jović, M.; Stanković, A.R.; Katsikas, L. Heavy metals in seafood mussels. Risks for human health. In Environmental Chemistry for A Sustainable World; nanotechnology and health risk; Lichtfouse, E., Schwarzbauer, J., Robert, D., Eds.; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2012; Volume 1, pp. 311–373. [Google Scholar]
- Perošević, A.; Pezo, L.; Joksimović, D.; Đurović, D.; Milašević, I.; Radomirović, M.; Stanković, S. The impacts of seawater physicochemical parameters and sediment metal contents on trace metal concentrations in mussels—A chemometric approach. Environ. Sci. Pollut. Res. 2018, 25, 28248–28263. [Google Scholar] [CrossRef]
- Karbe, L.; Antonacopoulos, N.; Schnier, C. The influence of water quality on accumulation of heavy metals in aquatic organisms. SIL Proc. 1975, 19, 2094–2101. [Google Scholar] [CrossRef]
- Lea, D.W. Trace elements in foraminiferal calcite. In Modern Foraminifera; Springer: Dordrecht, The Netherlands, 1999; pp. 259–277. [Google Scholar]
- Lemaire, N. Variations saisonnières de paramètres physiologiques chez la moule bleue Mytilus spp. dans différents sites d’élevage mytilicole de l’Est du Québec. Master’s Thesis, University of Québec, Rimouski, QC, Canada, 2005. [Google Scholar]
- Lemaire, N.; Pellerin, J.; Fournier, M.; Girault, L.; Tamigneaux, E.; Cartier, S.; Pelletier, E. Seasonal variations of physiological parameters in the blue mussel Mytilus spp. from farm sites of eastern Quebec. Aquaculture 2006, 261, 729–751. [Google Scholar] [CrossRef]
- Jitar, O.; Teodosiu, C.; Oros, A.; Plavan, G.; Nicoara, M. Bioaccumulation of heavy metals in marine organisms from the Romanian sector of the Black Sea. N. Biotechnol. 2015, 32, 369–378. [Google Scholar] [CrossRef]
- Kefi, J.J.; Mleiki, A.; Béjaoui, J.M.; El Menif, N.T. Seasonal variations of trace metal concentrations in the soft tissue of Lithophaga lithophaga collected from the Bizerte bay (Northern Tunisia, Mediterranean Sea). J. Aquac. Res. Dev. 2016, 7, 432. [Google Scholar]
- Marchitto, T.M.; Demenocal, P.B. Late Holocene variability of upper North Atlantic Deep Water temperature and salinity. Geochem. Geophys. Geosystems 2003, 4, 1100. [Google Scholar] [CrossRef]
- McGann, M. High–Resolution foraminiferal, isotopic, and trace element records from Holocene estuarine deposits of San Francisco Bay, California. J. Coast. Res. 2008, 24, 1092–1109. [Google Scholar] [CrossRef]
- Kocsis, L. Geochemical compositions of marine fossils as proxies for reconstructing ancient environmental conditions. Chimia 2011, 65, 787–791. [Google Scholar] [CrossRef]
- Grasso, M.; Licorish, W.H.; Diliberto, S.E.; Geremia, F.; Maniscalco, R.; Maugeri, S.; Pappalardo, G.; Rapisarda, F.; Scamarda, G. Geological Map of the Licata fold belt (South Central Sicily): Explanatory Notes; Annales Tectonicae: Firenze, Italy, 1998; Volume XII, pp. 51–58. [Google Scholar]
- Longhitano, S. Sedimentary features of incipient beachrock deposits along the coast of Simeto river delta (eastern Sicily, Italy). GeoActa 2003, 1, 99–113. [Google Scholar]
- Longhitano, S.; Colella, A. Geomorphology, sedimentology and recent evolution of the anthropogenically modified Simeto River delta system (eastern Sicily, Italy). Sediment. Geol. 2007, 194, 195–221. [Google Scholar] [CrossRef]
- Di Stefano, A.; De Pietro, R.; Monaco, C.; Zanini, A. Anthropogenic influence on coastal evolution: A case history from the Catania Gulf shoreline (eastern Sicily, Italy). Ocean Coast. Manag. 2013, 80, 133–148. [Google Scholar] [CrossRef]
- Lentini, F.; Carbone, S. Geologia della Sicilia–Geology of Sicily; Carta Geologica d’Italia: Roma, Italy, 2014; Volume 95, pp. 7–414. [Google Scholar]
- Sanfilippo, R.; Serio, D.; Deias, C.; Rosso, A. Sabellaria alveolata (Annelida, Polychaeta) bioconstructions and associated macroalgal community from Portopalo di Capo Passero (SE Sicily). Mediterr. Mar. Sci. 2022, 23, 150–156. [Google Scholar] [CrossRef]
- PAI. Piano Stralcio di Bacino per L’assetto Idrogeologico Della Regione Siciliana; Assessorato Territorio e Ambiente, Regione Sicilia: Palermo, Italy, 2004; p. 176. [Google Scholar]
- Martino, C.; Di Stefano, A.; Monaco, C.; Zanini, A.; Curcuruto, E.P. Fenomeni erosivi lungo il litorale di Marina di Butera (CL), Sicilia centro–meridionale. Geol. di Sicil. 2011, 3, 4–15. [Google Scholar]
- Borzì, L.; Anfuso, G.; Manno, G.; Distefano, S.; Urso, S.; Chiarella, D.; Di Stefano, A. Shoreline Evolution and Environmental Changes at the NW Area of the Gulf of Gela (Sicily, Italy). Land 2021, 10, 1034. [Google Scholar] [CrossRef]
- Nordstrom, D.K. Thermochemical redox equilibria of ZoBell’s solution. Geochim. Cosmochim. Acta 1977, 41, 1835–1841. [Google Scholar] [CrossRef]
- Drever, J.I. The Geochemistry of Natural Waters; Prentice Hall, Inc.: Englewood Cliffs, NJ, USA, 1988. [Google Scholar]
- Randazzo, P.; Caracaus, A.; Aiuppa, A.; Cardellini, C.; Chiodini, G.; Apollaro, C.; Paternoster, M.; Rosiello, A.; Vespasiano, G. Active degassing of crustal CO2 in areas of tectonic collision: A case study from the Pollino and Calabria sectors (Southern Italy). Front. Earth Sci. 2022, 10, 1–18. [Google Scholar] [CrossRef]
- Vespasiano, G.; Marini, L.; Muto, F.; Auqué, L.F.; Cipriani, M.; De Rosa, R.; Critelli, S.; Gimeno, M.J.; Blasco, M.; Dotsika, E.; et al. Chemical, isotopic and geotectonic relations of the warm and cold waters of the Cotronei (Ponte Coniglio), Bruciarello and Repole thermal areas, (Calabria—Southern Italy). Geothermics 2021, 96, 102228. [Google Scholar] [CrossRef]
- Nisi, B.; Vaselli, O.; Elio, J.; Giannini, L.; Tassi, F.; Guidi, M.; Darrah, T.H.; Maletic, E.L.; Delgado Huertas, A.; Marchionni, S. The Campo de Calatrava Volcanic Field (central Spain): Fluid geochemistry in a CO2–rich area. Appl. Geochem. 2019, 102, 153–170. [Google Scholar] [CrossRef]
- Vespasiano, G.; Cianflone, G.; Marini, L.; De Rosa, R.; Polemio, M.; Walraevens, K.; Vaselli, O.; Pizzino, L.; Cinti, D.; Capecchiacci, F.; et al. Hydrogeochemical and isotopic characterization of the Gioia Tauro coastal Plain (Calabria–southern Italy): A multidisciplinary approach for a focused management of vulnerable strategic systems. Sci. Total Environ. 2023, 862, 160694. [Google Scholar] [CrossRef]
- Apollaro, C.; Tripodi, V.; Vespasiano, G.; De Rosa, R.; Dotsika, E.; Fuoco, I.; Critelli, S.; Muto, F. Chemical, isotopic and geotectonic relations of the warm and cold waters of the Galatro and Antonimina thermal areas, southern Calabria, Italy. Mar. Pet. Geo. 2019, 109, 469–483. [Google Scholar] [CrossRef]
- Riding, R. Microbialites, stromatolites, and thrombolites. In Encyclopedia of Geobiology; Reitner, J., Thiel, V., Eds.; Encyclopedia of Earth Science Series; Springer: Heidelberg, Germany, 2011; pp. 635–654. [Google Scholar]
- Riding, R. Microbial carbonates: The geological record of calcified bacterial-algal mats and biofilms. Sedimentology 2000, 47, 179–214. [Google Scholar] [CrossRef]
- Van Driessche, A.E.S.; Stawski, T.M.; Kellermeier, M. Calcium sulfate precipitation pathways in natural and engineered environments. Chem. Geol. 2019, 530, 119274. [Google Scholar] [CrossRef]
- Lowenstam, H.A.; Weiner, S. On Biomineralization; Oxford University Press: New York, NY, USA, 1989. [Google Scholar]
- Mastandrea, A.; Barca, D.; Guido, A.; Tosti, F.; Russo, F. Rare earth element signatures in the Messinian pre-evaporitic Calcare di Base formation (Northern Calabria, Italy): Evidence of normal seawater deposition. Carbonates Evaporites 2010, 25, 133–143. [Google Scholar] [CrossRef]
- Benzerara, K.; Miot, J.; Morin, G.; Ona-Nguema, G.; Skouri-Panet, F.; Ferard, C. Significance, mechanisms and environmental implications of microbial biomineralization. CR Geosci. 2011, 343, 160–167. [Google Scholar] [CrossRef]
- Phillips, A.J.; Gerlach, R.; Lauchnor, E.; Mitchell, A.C.; Cunningham, A.B.; Spangler, L. Engineered applications of ureolytic biomineralization: A review. Biofouling 2013, 29, 715–733. [Google Scholar] [CrossRef] [PubMed]
- Anbu, P.; Kang, C.H.; Shin, Y.J.; So, J.S. Formations of calcium carbonate minerals by bacteria and its multiple applications. SpringerPlus 2016, 5, 250. [Google Scholar] [CrossRef] [PubMed]
- Riding, R.; Virgone, A. Hybrid Carbonates: In situ abiotic, microbial and skeletal coprecipitates. Earth Sci. Rev. 2020, 208, 103300. [Google Scholar] [CrossRef]
- Guido, A.; Gerovasileiou, V.; Russo, F.; Rosso, A.; Sanfilippo, R.; Voultsiadou, E.; Mastandrea, A. Composition and biostratinomy of sponge-rich biogenic crusts in sub marine caves (Aegean Sea, Eastern Mediterranean). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 534, 109338. [Google Scholar] [CrossRef]
- Guido, A.; Mastandrea, A.; Rosso, A.; Sanfilippo, R.; Russo, F. Micrite precipitation induced by sulphate reducing bacteria in serpulid bioconstructions from submarine caves (Syracuse, Sicily). Rend. Online Soc. Geol. Ital. 2012, 21, 933–934. [Google Scholar]
- Guido, A.; Sposato, M.; Palladino, G.; Vescogni, A.; Miriello, D. Biomineralization of primary carbonate cements: A new biosignature in the fossil record from the Anisian of Southern Italy. Lethaia 2022, 55, 1–21. [Google Scholar] [CrossRef]
- Riding, R.; Liang, L. Geobiology of microbial carbonates: Metazoan and seawater saturation state influences on secular trends during the Phanerozoic. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 101–115. [Google Scholar] [CrossRef]
- Guido, A.; Rosso, A.; Sanfilippo, R.; Russo, F.; Mastandrea, A. Microbial biomineralization in biotic crusts from a Pleistocene marine cave (NW Sicily, Italy). Geomicrobiol. J. 2017, 34, 864–872. [Google Scholar] [CrossRef]
- Guido, A.; Rosso, A.; Sanfilippo, R.; Miriello, D.; Belmonte, G. Skeletal vs microbialite geobiological role in bioconstructions of confined marine environments. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 593, 110920. [Google Scholar] [CrossRef]
- Bouthir, F.Z.; Chafik, A.; Benbrahim, S.; Souabi, S.; El Merdhy, H.; Messoudi, A. Qualité physico–chimique des eaux côtières du littoral de la Wilaya du grand Casablanca (océan Atlantique marocain) utilisant la moule Mytilus galloprovincialis comme indicateur de la contamination métallique. Mar. life 2004, 14, 59–70. [Google Scholar]
- Cole, V.J.; Chapman, M.G.; Underwood, A.J. Landscapes and life-histories influence colonisation of polychaetes to intertidal biogenic habitats. J. Exp. Mar. Biol. Ecol. 2007, 348, 191–199. [Google Scholar] [CrossRef]
- La Porta, B.; Nicoletti, L. Sabellaria alveolata (Linnaeus) reefs in the central Tyrrhenian Sea (Italy) and associated polychaete fauna. Zoosymposia 2009, 2, 527–536. [Google Scholar] [CrossRef]
- Gravina, M.F.; Cardone, F.; Bonifazi, A.; Bertrandino, M.S.; Chimienti, G.; Longo, C.; Marzano, C.N.; Moretti, M.; Lisco, S.; Moretti, V.; et al. Sabellaria spinulosa (Polychaeta, Annelida) reefs in the Mediterranean Sea: Habitat mapping, dynamics and associated fauna for conservation management. Estuar. Coast. Shelf Sci. 2018, 200, 248–257. [Google Scholar] [CrossRef]
- Curd, A.; Fabrice Pernet, F.; Corporeau, C.; Delisl, L.; Firt, L.B.; Nunes, F.L.D.; Dubois, S. Connecting organic to mineral: How the physiological state of an ecosystem-engineer is linked to its habitat structure. Ecol. Indic. 2019, 98, 49–60. [Google Scholar] [CrossRef]






| ID | Sampling Period | T | pH | EC | Ca | Mg | K | Na | Cl | SO4 | HCO3 | F− | Br |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| °C | mS/cm | mg L−1 | mg L−1 | mg L−1 | mg L−1 | mg L−1 | mg L−1 | mg L−1 | mg L−1 | mg L−1 | |||
| Falconara | Autumn 2020 | 24.1 | 8.4 | 56.9 | 618.5 | 1119.0 | 337.0 | 9046.9 | 21,766.9 | 2602.1 | 151.8 | 12.8 | 67.3 |
| Falconara | Spring 2021 | 24.1 | 8.2 | 44.6 | 521.4 | 1506.1 | 490.8 | 11,721.3 | 21,712.1 | 2465.4 | 158.6 | 14.1 | 64.6 |
| Falconara | Summer 2021 | 25.5 | 8.2 | 44.9 | 585.8 | 1504.9 | 441.5 | 12,419.4 | 26,157.9 | 2772.6 | 183.1 | 25.5 | 68.7 |
| Falconara | Autumn 2021 | 19.4 | 8.3 | 47.1 | 646.7 | 1471.0 | 479.2 | 11,761.7 | 21,629.7 | 2652.8 | 158.6 | n.d. | 59.4 |
| Portopalo | Autumn 2020 | 26.9 | 8.5 | 57.8 | 571.2 | 1133.9 | 351.2 | 9534.9 | 21,985.2 | 2623.8 | 154.1 | 8.9 | 70.9 |
| Portopalo | Spring 2021 | 22.9 | 8.3 | 45.1 | 842.4 | 1585.1 | 512.0 | 11,701.5 | 21,293.4 | 2675.6 | 122.0 | 13.4 | 71.7 |
| Portopalo | Summer 2021 | 27.2 | 8.3 | 45.9 | 649.6 | 1548.8 | 448.0 | 12,742.9 | 27,273.1 | 2723.8 | 198.3 | 13.2 | 64.9 |
| Portopalo | Autumn 2021 | 22.7 | 8.2 | 47.3 | 587.4 | 1487.2 | 451.8 | 12,052.2 | 24,159.0 | 2610.3 | 146.4 | 11.0 | 66.0 |
| Simeto | Autumn 2020 | 23.0 | 8.3 | 57.1 | 631.6 | 1145.3 | 350.5 | 8963.3 | 21,492.8 | 2622.4 | 157.1 | 11.4 | 63.8 |
| Simeto | Spring 2021 | 22.1 | 8.2 | 41.3 | 759.6 | 1438.2 | 467.1 | 10,944.5 | 19,631.3 | 2067.4 | 183.1 | 20.7 | 63.0 |
| Simeto | Summer 2021 | 28.1 | 8.2 | 43.3 | 605.8 | 1453.4 | 416.7 | 12,096.1 | 25,313.1 | 2647.6 | 225.8 | 16.2 | 68.5 |
| Simeto | Autumn 2021 | 22.8 | 8.2 | 45.2 | 832.2 | 1406.8 | 413.7 | 11,558.5 | 23,082.0 | 2443.5 | 173.9 | 14.5 | 61.3 |
| ID | Sampling Period | Li | B | Cr | Ni | Cu | Zn | As | Se | Rb | Sr | Ti |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| μg L−1 | μg L−1 | μg L−1 | μg L−1 | μg L−1 | μg L−1 | μg L−1 | μg L−1 | μg L−1 | μg L−1 | μg L−1 | ||
| Falconara | Autumn 2020 | 227.9 | 5980.4 | 109.9 | 24.1 | 51.2 | 345.5 | 117.4 | 701.4 | 132.2 | 9153.2 | 672.8 |
| Falconara | Spring 2021 | 210.0 | 5989.4 | 75.4 | 27.1 | 54.9 | 332.6 | 69.1 | 734.6 | 136.9 | 9194.9 | 653.2 |
| Falconara | Summer 2021 | 210.1 | 6063.6 | 101.1 | 35.0 | 64.1 | 269.9 | 75.0 | 775.8 | 128.5 | 9466.0 | 635.5 |
| Falconara | Autumn 2021 | 191.3 | 6020.3 | 25.8 | 90.3 | 216.9 | 623.9 | 48.4 | 346.0 | 133.2 | 9144.3 | 674.7 |
| Portopalo | Autumn 2020 | 222.0 | 6066.1 | 106.3 | 31.9 | 50.8 | 385.3 | 97.9 | 718.7 | 141.0 | 9393.2 | 700.1 |
| Portopalo | Spring 2021 | 217.9 | 6048.7 | 97.6 | 24.9 | 56.8 | 421.2 | 91.3 | 718.6 | 133.4 | 9567.5 | 685.5 |
| Portopalo | Summer 2021 | 223.1 | 6327.6 | 87.2 | 31.1 | 62.9 | 325.8 | 81.9 | 791.8 | 138.1 | 9635.5 | 671.8 |
| Portopalo | Autumn 2021 | 201.2 | 8038.4 | 65.0 | 137.2 | 268.9 | 8090.7 | 54.2 | 361.6 | 120.7 | 8701.2 | 688.1 |
| Simeto | Autumn 2020 | 216.1 | 6181.1 | 94.4 | 23.3 | 55.9 | 271.7 | 86.1 | 722.4 | 134.7 | 9321.8 | 675.5 |
| Simeto | Spring 2021 | 192.8 | 5381.4 | 78.6 | 23.8 | 48.7 | 289.5 | 76.7 | 694.5 | 117.8 | 8420.3 | 603.1 |
| Simeto | Summer 2021 | 223.9 | 6525.2 | 108.2 | 29.5 | 59.0 | 291.7 | 70.9 | 681.9 | 130.8 | 9260.7 | 615.5 |
| Simeto | Autumn 2021 | 187.6 | 6954.8 | 23.8 | 74.9 | 270.3 | 7508.9 | 50.7 | 307.2 | 112.9 | 8236.4 | 600.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deias, C.; Guido, A.; Sanfilippo, R.; Apollaro, C.; Dominici, R.; Cipriani, M.; Barca, D.; Vespasiano, G. Elemental Fractionation in Sabellariidae (Polychaeta) Biocement and Comparison with Seawater Pattern: A New Environmental Proxy in a High-Biodiversity Ecosystem? Water 2023, 15, 1549. https://doi.org/10.3390/w15081549
Deias C, Guido A, Sanfilippo R, Apollaro C, Dominici R, Cipriani M, Barca D, Vespasiano G. Elemental Fractionation in Sabellariidae (Polychaeta) Biocement and Comparison with Seawater Pattern: A New Environmental Proxy in a High-Biodiversity Ecosystem? Water. 2023; 15(8):1549. https://doi.org/10.3390/w15081549
Chicago/Turabian StyleDeias, Claudia, Adriano Guido, Rossana Sanfilippo, Carmine Apollaro, Rocco Dominici, Mara Cipriani, Donatella Barca, and Giovanni Vespasiano. 2023. "Elemental Fractionation in Sabellariidae (Polychaeta) Biocement and Comparison with Seawater Pattern: A New Environmental Proxy in a High-Biodiversity Ecosystem?" Water 15, no. 8: 1549. https://doi.org/10.3390/w15081549
APA StyleDeias, C., Guido, A., Sanfilippo, R., Apollaro, C., Dominici, R., Cipriani, M., Barca, D., & Vespasiano, G. (2023). Elemental Fractionation in Sabellariidae (Polychaeta) Biocement and Comparison with Seawater Pattern: A New Environmental Proxy in a High-Biodiversity Ecosystem? Water, 15(8), 1549. https://doi.org/10.3390/w15081549