
Life-History Traits and Acclimation Ability of a Copepod Species from the Dripping Waters of the Corchia Cave (Apuan Alps, Tuscany, Italy)

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
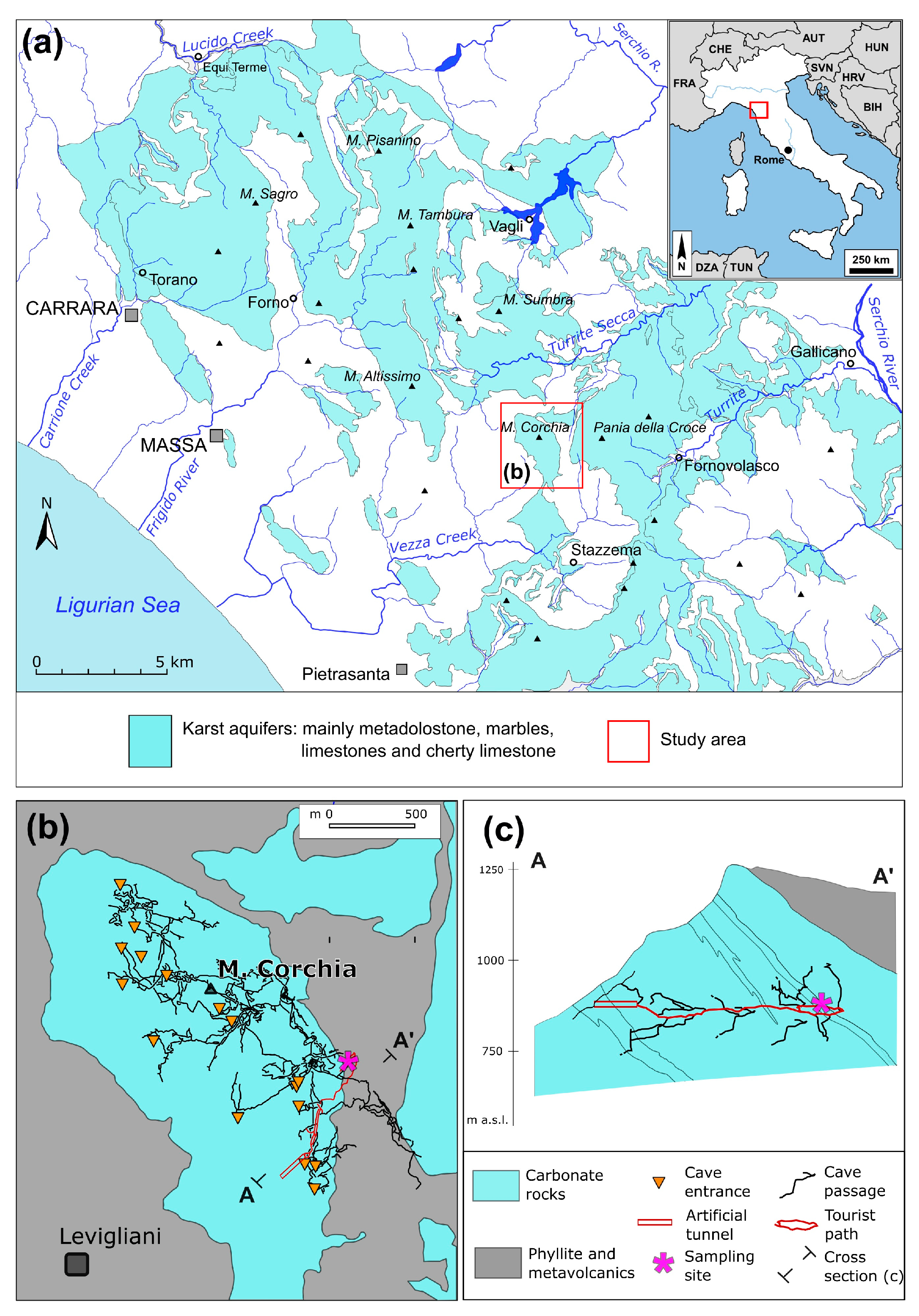
2.1. Study Area
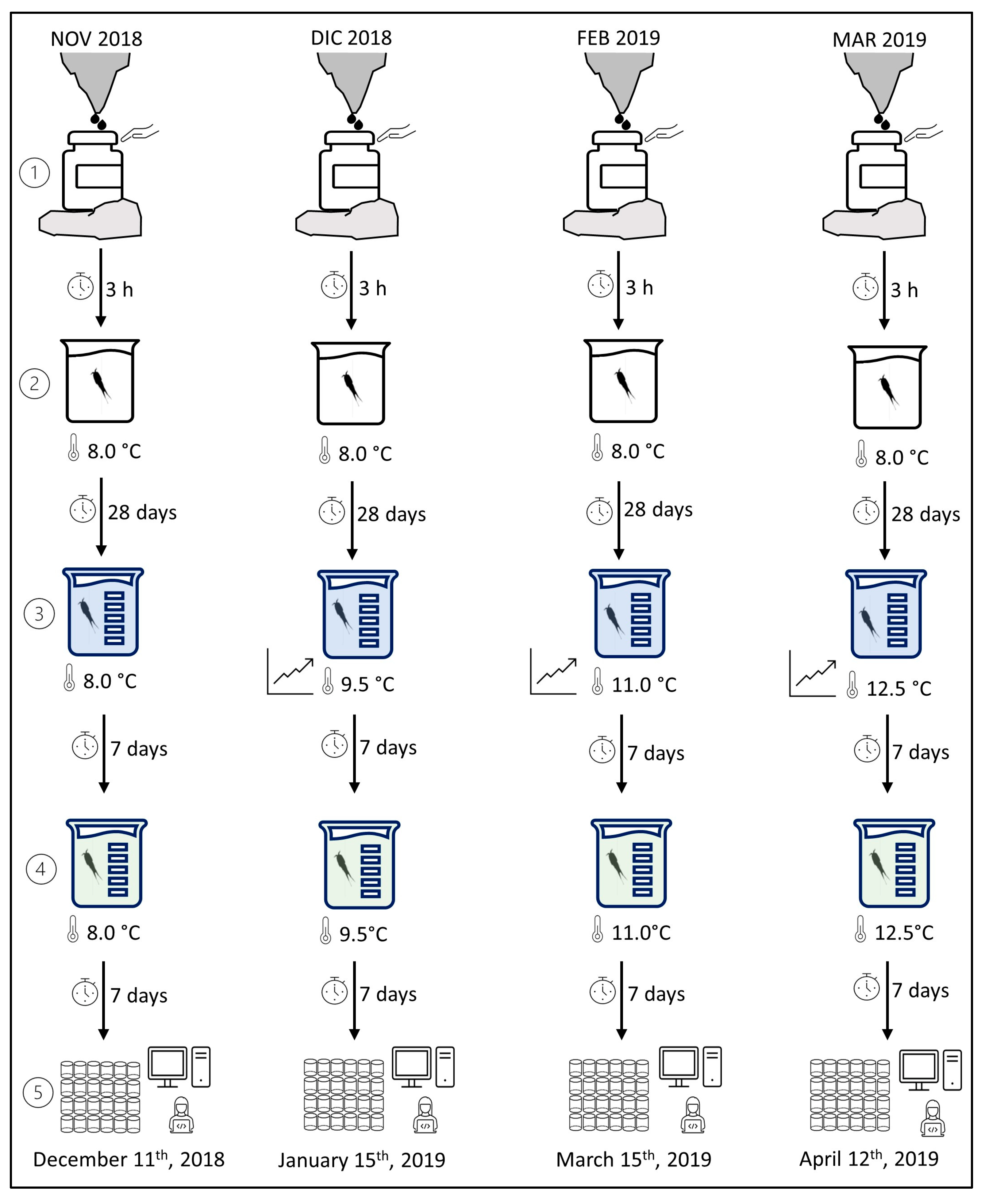
2.2. Biological Monitoring and Rearing
2.3. Protocol of Respirometry Trials
2.4. Statistical Analyses
3. Results
3.1. Life-History Traits
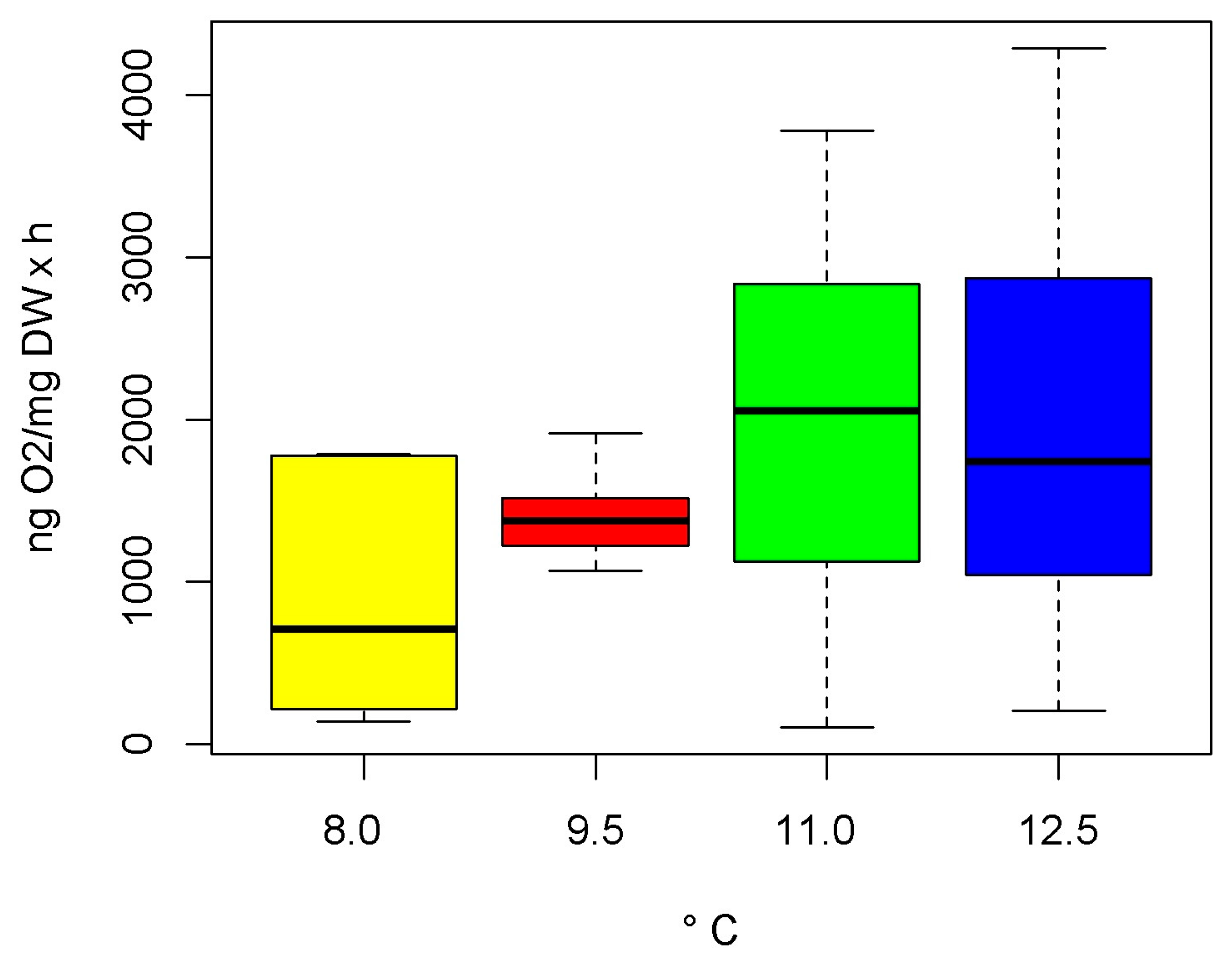
3.2. Oxygen Consumption Rates
4. Discussion
4.1. Life-History Traits
4.2. Oxygen Consumption Rates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mammola, S.; Amorim, I.R.; Bichuette, M.E.; Borges, P.A.; Cheeptham, N.; Cooper, S.J.B.; Culver, D.C.; Deharveng, L.; Eme, D.; Ferreira, R.L.; et al. Fundamental research questions in subterranean biology. Biol. Rev. 2020, 95, 1855–1872. [Google Scholar] [CrossRef] [PubMed]
- Mammola, S.; Lunghi, E.; Bilandžija, H.; Cardoso, P.; Grimm, V.; Schmidt, S.I.; Hesselberg, T.; Martínez, A. Collecting eco-evolutionary data in the dark: Impediments to subterranean research and how to overcome them. Ecol. Evol. 2021, 11, 5911–5926. [Google Scholar] [CrossRef] [PubMed]
- Hose, G.C.; Chariton, A.A.; Daam, M.A.; Di Lorenzo, T.; Galassi, D.M.P.; Halse, S.A.; Reboleira, A.S.P.; Robertson, A.L.; Schmidt, S.I.; Korbel, K.L. Invertebrate traits, diversity and the vulnerability of groundwater ecosystems. Funct. Ecol. 2022, 36, 2200–2214. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Di Marzio, W.D.; Spigoli, D.; Baratti, M.; Messana, G.; Cannicci, S.; Galassi, D.M.P. Metabolic rates of a hypogean and an epigean species of copepod in an alluvial aquifer. Freshw. Biol. 2015, 60, 426–435. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Reboleira, A.S.P. Thermal acclimation and metabolic scaling of a groundwater asellid in the climate change scenario. Sci. Rep. 2022, 12, 1–11. [Google Scholar] [CrossRef]
- Carpenter, J.H. Forty-year natural history study of Bahalana geracei Carpenter, 1981, an anchialine cave-dwelling isopod (Crustacea, Isopoda, Cirolanidae) from San Salvador Island, Bahamas: Reproduction, growth, longevity, and population structure. Subterran. Biol. 2021, 37, 105–156. [Google Scholar] [CrossRef]
- Rütz, N.K.; Marxsen, J.; Wolters, V. Long-term cultivation of the groundwater amphipod Niphargus aquilex (Crustacea). Hydrobiologia 2023, 850, 269–281. [Google Scholar] [CrossRef]
- Gillooly, J.F.; Brown, J.H.; West, G.B.; Savage, V.M.; Charnov, E.L. Effects of size and temperature on metabolic rate. Science 2001, 293, 2248–2251. [Google Scholar] [CrossRef]
- Fossen, E.I.F.; Pélabon, C.; Einum, S. Genetic and environmental effects on the scaling of metabolic rate with body size. J. Exp. Biol. 2019, 222, jeb193243. [Google Scholar] [CrossRef]
- Shokri, M.; Cozzoli, F.; Vignes, F.; Bertoli, M.; Pizzul, E.; Basset, A. Metabolic rate and climate change across latitudes: Evidence of mass-dependent responses in aquatic amphipods. J. Exp. Biol. 2022, 225, jeb244842. [Google Scholar] [CrossRef]
- Willmer, P.; Stone, G.; Johnston, I. Environmental Physiology of Animals, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2009; pp. 1–768. [Google Scholar]
- Huey, R.B.; Kingsolver, J.G. Evolution of thermal sensitivity of ectotherm performance. Trends Ecol. Evol. 1989, 4, 131–135. [Google Scholar] [CrossRef]
- Mermillod-Blondin, F.; Lefour, C.; Lalouette, L.; Renault, D.; Malard, F.; Simon, L.; Douady, C.J. Thermal tolerance breadths among groundwater crustaceans living in a thermally constant environment. J. Exp. Biol. 2013, 216, 1683–1694. [Google Scholar] [CrossRef]
- Peck, L.S.; Morley, S.A.; Richard, J.; Clark, M.S. Acclimation and thermal tolerance in Antarctic marine ectotherms. J. Exp. Biol. 2014, 217, 16–22. [Google Scholar] [CrossRef]
- Iepure, S. Ice cave fauna. In Ice Caves, 1st ed.; Perşoiu, A., Lauritzen, A.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 163–171. [Google Scholar] [CrossRef]
- Issartel, J.; Hervant, F.; Voituron, Y.; Renault, D.; Vernon, P. Behavioural, ventilatory and respiratory responses of epigean and hypogean crustaceans to different temperatures. Comp. Biochem. Physiol. Mol. Amp Integr. Physiol. 2005, 141, 1–7. [Google Scholar] [CrossRef]
- Colson-Proch, C.; Morales, A.; Hervant, F.; Konecny, L.; Moulin, C.; Douady, C.J. First cellular approach of the effects of global warming on groundwater organisms: A study of the HSP70 gene expression. Cell Stress Chaperones 2010, 15, 259–270. [Google Scholar] [CrossRef]
- Pallarés, S.; Colado, R.; Botella-Cruz, M.; Montes, A.; Balart-García, P.; Bilton, D.T.; Millán, A.; Ribera, I.; Sánchez-Fernández, D. Loss of heat acclimation capacity could leave subterranean specialists highly sensitive to climate change. Anim. Conserv. 2021, 24, 482–490. [Google Scholar] [CrossRef]
- Deharveng, L.; Stoch, F.; Gibert, J.; Bedos, A.; Galassi, D.M.P.; Zagmajster, M.; Brancelj, A.; Camacho, A.; Fiers, F.; Martin, P.; et al. Groundwater biodiversity in Europe. Freshw. Biol. 2009, 54, 709–726. [Google Scholar] [CrossRef]
- Galassi, D.M.P. Groundwater copepods: Diversity patterns over ecological and evolutionary scales. Hydrobiologia 2001, 453, 227–253. [Google Scholar] [CrossRef]
- Galassi, D.M.P.; Huys, R.; Reid, J.W. Diversity, ecology and evolution of groundwater copepods. Freshw. Biol. 2009, 54, 691–708. [Google Scholar] [CrossRef]
- Ikeda, T. Metabolism in mesopelagic and bathypelagic copepods: Reply to Childress et al. (2008). Mar. Ecol. Prog. Ser. 2008, 373, 193–198. [Google Scholar] [CrossRef]
- Dole-Olivier, M.J.; Galassi, D.M.P.; Marmonier, P.; Creuzé des Châtelliers, M. The biology and ecology of lotic microcrustaceans. Freshw. Biol. 2000, 44, 63–91. [Google Scholar] [CrossRef]
- Fattorini, S.; Borges, P.A.V.; Fiasca, B.; Galassi, D.M.P. Trapped in the web of water: Groundwater-fed springs are island-like ecosystems for the meiofauna. Ecol. Evol. 2006, 6, 8389–8401. [Google Scholar] [CrossRef] [PubMed]
- Pleşa, C. Recherches sur la périodicité de reproduction chez les cavernicoles. Spelunca Mémoires 1967, 5, 295–299. [Google Scholar]
- Pleşa, C. Cercetări asupra periodicităţii reproductive la unele crustacee cavernicole troglobionte. In Résumé de Thése de Doctorat en Biologie; Inst. Biologie “Tr. Săvulescu”: București, Romania, 1969; p. 30. [Google Scholar]
- Lescher-Moutoue, F. Sur la biologie et l’écologie des copépodes cyclopides hypogés (Crustacés). Ann. Spéliol. 1973, 28, 429–502. [Google Scholar]
- Notenboom, J.; Boessenkool, J.J. Toxicity of Selected Pesticides to the Groundwater Copepod Parastenocaris germanica (Crustacea); RIVM Rapport 710302005; RIVM: Bilthoven, The Nederlands, 1994. [Google Scholar]
- Baneschi, I.; Piccini, L.; Regattieri, E.; Isola, I.; Guidi, M.; Lotti, L.; Mantelli, F.; Menichetti, M.; Drysdale, R.N.; Zanchetta, G. Hypogean microclimatology and hydrology of the 800–900 m asl in the Monte Corchia Cave (Tuscany, Italy): Preliminary considerations and implications for paleoclimatological studies. Acta Carsologica 2011, 40, 175–187. [Google Scholar] [CrossRef]
- Piccini, L.; Zanchetta, G.; Drysdale, R.N.; Hellstrom, J.; Isola, I.; Fallick, A.E.; Leone, G.; Doveri, M.; Mussi, M.; Mantelli, F.; et al. The environmental features of the Monte Corchia cave system (Apuan Alps, central Italy) and their effects on speleothem growth. Int. J. Speleol. 2008, 37, 153–172. [Google Scholar] [CrossRef]
- Doveri, M.; Piccini, L.; Menichini, M. Hydrodynamic and geochemical features of metamorphic carbonate aquifers and implications for water management: The Apuan Alps (NW Tuscany, Italy) case study. In The Handbook of Environmental Chemistry; Younos, T., Schreiber, M., Kosič Ficco, K., Eds.; Springer International Publishing: New York, NY, USA, 2018; Volume 68, pp. 209–249. [Google Scholar] [CrossRef]
- Piccini, L. Speleogenesis in highly geodynamic contexts: The quaternary evolution of Monte Corchia multi-level karst system (Alpi Apuane, Italy). Geomorphology 2011, 134, 49–61. [Google Scholar] [CrossRef]
- Piccini, L.; Mantelli, F.; Montigiani, A.; Cecconi, E.; Lotti, L. Idrogeologia del sistema carsico del Monte Corchia: Sintesi dei dati e delle attuali conoscenze. Acta Apuana 2015, 2012, 23–32. [Google Scholar]
- Mantelli, F.; Piccini, L.; Scala, C.; Menichetti, S.; Lotti, L.; Montigiani, A.; De Sio, F.; Occhini, F. Antro Del Corchia-1997–2017 20 Anni Di Monitoraggio e Ricerche; ARPAT: Florence, Italy, 2021; ISBN 978-88-96693-27-8. Available online: https://www.arpat.toscana.it/documentazione/catalogo-pubblicazioni-arpat/antro-del-corchia-1997-2017 (accessed on 21 February 2023).
- Pipan, T. Epikarst—A promising habitat: Copepod fauna, its diversity and ecology: A case study from Slovenia (Europe). In Carsologica, 1st ed.; Gabrovšek, F., Ed.; Založba ZRC: Ljubljana, Slovenia, 2005; Volume 5. [Google Scholar] [CrossRef]
- Pipan, T.; Culver, D.C. Estimating biodiversity in the epikarstic zone of a West Virginia cave. J. Cave Karst. Stud. 2005, 67, 103–1091. [Google Scholar]
- Pipan, T.; Culver, D.C. Regional species richness in an obligate subterranean dwelling fauna–epikarst copepods. J. Biogeogr. 2007, 34, 854–861. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Di Marzio, W.D.; Fiasca, B.; Galassi, D.M.P.; Korbel, K.; Iepure, S.; Hose, G.C. Recommendations for ecotoxicity testing with stygobiotic species in the framework of groundwater environmental risk assessment. Sci. Total Environ. 2019, 681, 292–304. [Google Scholar] [CrossRef]
- Taylor, C.A.; Stefan, H.G. Shallow groundwater temperature response to climate change and urbanization. J. Hydrol. 2009, 375, 601–612. [Google Scholar] [CrossRef]
- Bovolo, C.I.; Parkin, G.; Sophocleous, M. Groundwater Resources, Climate and Vulnerability. Environ. Res. Lett. 2009, 4, 035001. [Google Scholar] [CrossRef]
- Menberg, K.; Blum, P.; Kurylyk, B.L.; Bayer, P. Observed groundwater temperature response to recent climate change. Hydrol. Earth Syst. Sci. 2014, 18, 4453–4466. [Google Scholar] [CrossRef]
- Di Lorenzo, T.; Galassi, D.M.P. Effect of temperature rising on the stygobitic crustacean species Diacyclops belgicus: Does global warming affect groundwater populations? Water 2017, 9, 951. [Google Scholar] [CrossRef]
- Cifoni, M.; Galassi, D.M.P.; Faraloni, C.; Di Lorenzo, T. Test procedures for measuring the (sub) chronic effects of chemicals on the freshwater cyclopoid Eucyclops serrulatus. Chemosphere 2017, 173, 89–98. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Feller, R.J.; Warwick, R.M. Energetics. In Introduction to the Study of Meiofauna, 1st ed.; Higgins, R.P., Thiel, H., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1988; pp. 181–196. [Google Scholar]
- Reiss, J.; Schmid-Araya, J.M. Existing in plenty: Abundance, biomass and diversity of ciliates and meiofauna in small streams. Freshw. Biol. 2008, 53, 652–668. [Google Scholar] [CrossRef]
- Reiss, J.; Schmid-Araya, J.M. Life history allometries and production of small fauna. Ecology 2010, 91, 497–507. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial; PRIMER-E Ltd.: Plymouth, UK, 2015. [Google Scholar]
- Galassi, D.M.P.; Fiasca, B.; Di Lorenzo, T.; Montanari, A.; Porfirio, S.; Fattorini, S. Groundwater biodiversity in a chemoautotrophic cave ecosystem: How geochemistry regulates microcrustacean community structure. Aquat. Ecol. 2017, 51, 75–90. [Google Scholar] [CrossRef]
- Dickson, G.W. The Importance of Cave Mud Sediments in Food Preference, Growth and Mortality of the Troglobitic Amphipod Crustacean Crangonyx antennatus Packard (Crangonyctidae). Crustaceana 1979, 36, 129–140. [Google Scholar] [CrossRef]
- Giere, O. Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments, 1st ed.; Springer Science: Berlin/Heidelberg, Germany, 2009; pp. 1–527. [Google Scholar]
- Di Lorenzo, T.; Di Cicco, M.; Di Censo, D.; Galante, A.; Boscaro, F.; Messana, G.; Galassi, D.M.P. Environmental risk assessment of propranolol in the groundwater bodies of Europe. Environ. Pollut. 2019, 255, 113189. [Google Scholar] [CrossRef] [PubMed]
- Sarvala, J. A parthenogenetic life cycle in a population of Canthocamptus staphylinus (Copepoda, Harpacticoida). Hydrobiologia 1979, 62, 113–129. [Google Scholar] [CrossRef]
- Rouch, R. Sur l’ecologie des eaux souterraines dans la karst. Stygologia 1986, 2, 352–398. [Google Scholar]
- Glatzel, T. On the biology of Parastenocaris phyllura Kiefer (Copepoda, Harpacticoida). Stygologia 1990, 5, 131–136. [Google Scholar]
- Di Lorenzo, T.; Cifoni, M.; Baratti, M.; Pieraccini, G.; Di Marzio, W.D.; Galassi, D.M.P. Four scenarios of environmental risk of diclofenac in European groundwater ecosystems. Environ. Pollut. 2021, 287, 117315. [Google Scholar] [CrossRef]
- Suárez-Morales, E. Class Maxillopoda. In Thorp and Covich’s Freshwater Invertebrates, 1st ed.; Thorp, H., Covich, A.P., Eds.; Academic Press: New York, NY, USA, 2015; pp. 709–755. [Google Scholar]
- Elgmork, K.; Eie, J.A. Two-and three-year life cycles in the planktonic copepod Cyclops scutifer in two high mountain lakes. Ecography 1989, 12, 60–69. [Google Scholar] [CrossRef]
- Van den Bosch, F.; Santer, B. Cannibalism in Cyclops abyssorum. Oikos 1993, 67, 19–28. [Google Scholar] [CrossRef]
- Glatzel, T.; Schminke, H.K. Mating behaviour of the groundwater copepod Parastenocaris phyllura Kiefer, 1938 (Copepoda: Harpacticoida). Bijdr. Dierkd. 1996, 66, 103–108. [Google Scholar] [CrossRef]
- Reid, J.W.; Williamson, C.E. Copepoda. In Ecology and Classification of North American Freshwater Invertebrates, 2nd ed.; Thorp, H., Covich, A.P., Eds.; Academic Press: New York, NY, USA, 2010; pp. 829–899. [Google Scholar]
- Ikeda, T. Metabolic rates of epipelagic marine zooplankton as a function of body mass and temperature. Mar. Biol. 1985, 85, 1–11. [Google Scholar] [CrossRef]
- Ikeda, T.; Kanno, Y.; Ozaki, K.; Shinada, A. Metabolic rates of epipelagic marine copepods as a function of body mass and temperature. Mar. Biol. 2001, 139, 587–596. [Google Scholar] [CrossRef]
- Hervant, F.; Mathieu, J.; Barré, H.; Simon, K.; Pinon, C. Comparative study on the behavioral, ventilatory, and respiratory responses of hypogean and epigean crustaceans to long-term starvation and subsequent feeding. Comp. Biochem. Physiol. Mol. Amp Integr. Physiol. 1997, 118, 1277–1283. [Google Scholar] [CrossRef]
- Wilhelm, F.M.; Taylor, S.J.; Adams, G.L. Comparison of routine metabolic rates of the stygobite, Gammarus acherondytes (Amphipoda: Gammaridae) and the stygophile, Gammarus troglophilus. Freshw. Biol. 2006, 51, 1162–1174. [Google Scholar] [CrossRef]
- Scheffler, M.L.; Barreto, F.S.; Mueller, C.A. Rapid metabolic compensation in response to temperature change in the intertidal copepod, Tigriopus californicus. Comp. Biochem. Physiol. Mol. Amp Integr. Physiol. 2019, 230, 131–137. [Google Scholar] [CrossRef]
- Olkova, A. Intraspecific sensitivity to toxicants—A methodological problem of bioassay. J. Ecol. Eng. 2021, 22, 113–122. [Google Scholar] [CrossRef]
- Culver, D.C.; Poulson, T.L. Oxygen consumption and activity in closely related amphipod populations from cave and surface habitats. Am. Midl. Nat. 1971, 85, 74–84. [Google Scholar] [CrossRef]
- Hervant, F.; Mathieu, J.; Messana, G. Oxygen consumption and ventilation in declining oxygen tension and posthypoxic recovery in epigean and hypogean crustaceans. J. Crustac. Biol. 1998, 18, 717–727. [Google Scholar] [CrossRef]
- Beasley-Hall, P.G.; Bertozzi, T.; Bradford, T.M.; Foster, C.S.P.; Jones, K.; Tierney, S.M.; Humphreys, W.F.; Austin, A.D.; Cooper, S.J.B. Differential transcriptomic responses to heat stress in surface and subterranean diving beetles. Sci. Rep. 2022, 12, 16194. [Google Scholar] [CrossRef]
- Wilson, R.; Franklin, C.; Davison, W.; Kraft, P. Stenotherms at sub-zero temperatures: Thermal dependence of swimming performance in Antarctic fish. J. Comp. Physiol. 2001, 171, 263–269. [Google Scholar] [CrossRef]
- Clarke, A.; Johnston, N.M. Scaling of metabolic rate with body mass and temperature in teleost fish. J. Anim. Ecol. 1999, 68, 893–905. [Google Scholar] [CrossRef]
- Peck, L.S.; Conway, L.Z. The myth of metabolic cold adaptation: Oxygen consumption in stenothermal Antarctic bivalves. Geol. Soc. Spec. Publ. 2000, 177, 441–450. [Google Scholar] [CrossRef]
- Issartel, J.; Voituron, Y.; Hervant, F. Impact of temperature on the survival, the activity and the metabolism of the cave-dwelling Niphargus virei, the ubiquitous stygobiotic N. rhenorhodanensis and the surface-dwelling Gammarus fossarum (Crustacea, Amphipoda). Subterr. Bio. 2007, 5, 9–14. [Google Scholar]
- Tanaka, K.; Udagawa, T. Cold adaptation of the terrestrial isopod, Porcellio scaber, to subnivean environments. J. Comp. Physiol. 1993, 163, 439–444. [Google Scholar]
- Paul, R.J.; Zeis, B.; Lamkemeyer, T.; Seidl, M.; Pirow, R. Control of oxygen transport in the microcrustacean Daphnia: Regulation of haemoglobin expression as central mechanism of adaptation to different oxygen and temperature conditions. Acta Physiol. Scand. 2004, 182, 259–275. [Google Scholar] [CrossRef]
- Hochachka, P.W.; Somero, G.N. Biochemical Adaptation: Mechanism and Process in Physiological Evolution, 1st ed.; Oxford University Press: New York, NY, USA, 2002. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | T | μ | sd | Min | Max |
|---|---|---|---|---|---|
| 10 | 8.0 | 913 | 701 | 140 | 1789 |
| 21 | 9.5 | 1388 | 202 | 1068 | 1914 |
| 8 | 11.0 | 1989 | 1214 | 104 | 3779 |
| 15 | 12.5 | 1941 | 1287 | 208 | 4286 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Lorenzo, T.; Galassi, D.M.P.; Tabilio Di Camillo, A.; Pop, M.M.; Iepure, S.; Piccini, L. Life-History Traits and Acclimation Ability of a Copepod Species from the Dripping Waters of the Corchia Cave (Apuan Alps, Tuscany, Italy). Water 2023, 15, 1356. https://doi.org/10.3390/w15071356
Di Lorenzo T, Galassi DMP, Tabilio Di Camillo A, Pop MM, Iepure S, Piccini L. Life-History Traits and Acclimation Ability of a Copepod Species from the Dripping Waters of the Corchia Cave (Apuan Alps, Tuscany, Italy). Water. 2023; 15(7):1356. https://doi.org/10.3390/w15071356
Chicago/Turabian StyleDi Lorenzo, Tiziana, Diana Maria Paola Galassi, Agostina Tabilio Di Camillo, Maria Mirabela Pop, Sanda Iepure, and Leonardo Piccini. 2023. "Life-History Traits and Acclimation Ability of a Copepod Species from the Dripping Waters of the Corchia Cave (Apuan Alps, Tuscany, Italy)" Water 15, no. 7: 1356. https://doi.org/10.3390/w15071356
APA StyleDi Lorenzo, T., Galassi, D. M. P., Tabilio Di Camillo, A., Pop, M. M., Iepure, S., & Piccini, L. (2023). Life-History Traits and Acclimation Ability of a Copepod Species from the Dripping Waters of the Corchia Cave (Apuan Alps, Tuscany, Italy). Water, 15(7), 1356. https://doi.org/10.3390/w15071356