
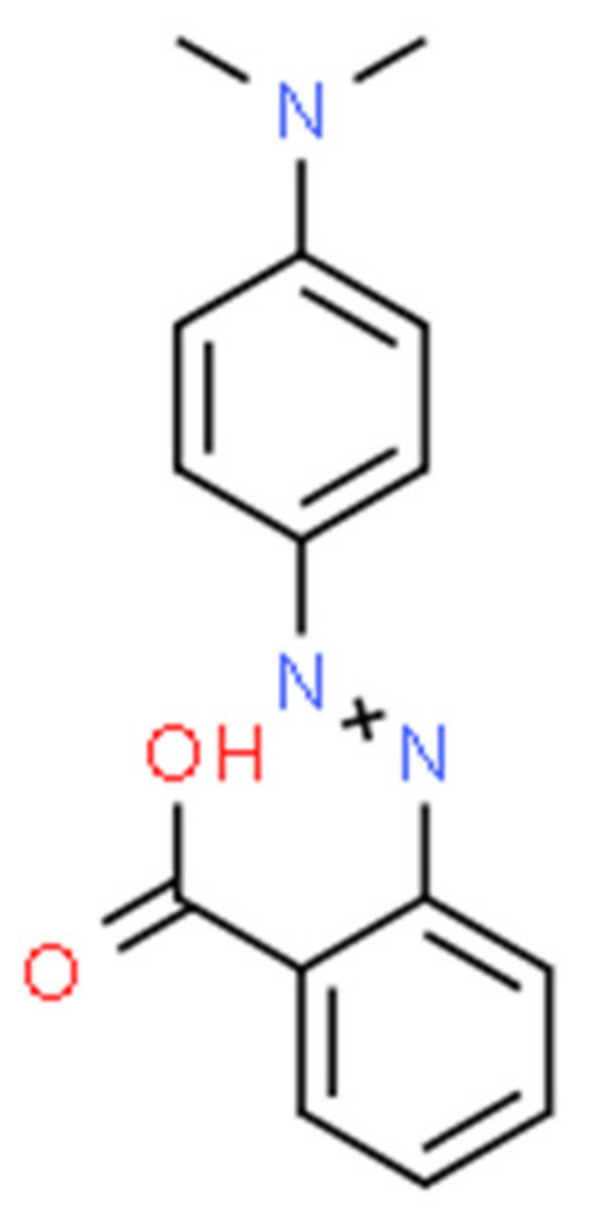
Microbial Electrochemical Treatment of Methyl Red Dye Degradation Using Co-Culture Method


 ,
,  ,
, 
Abstract
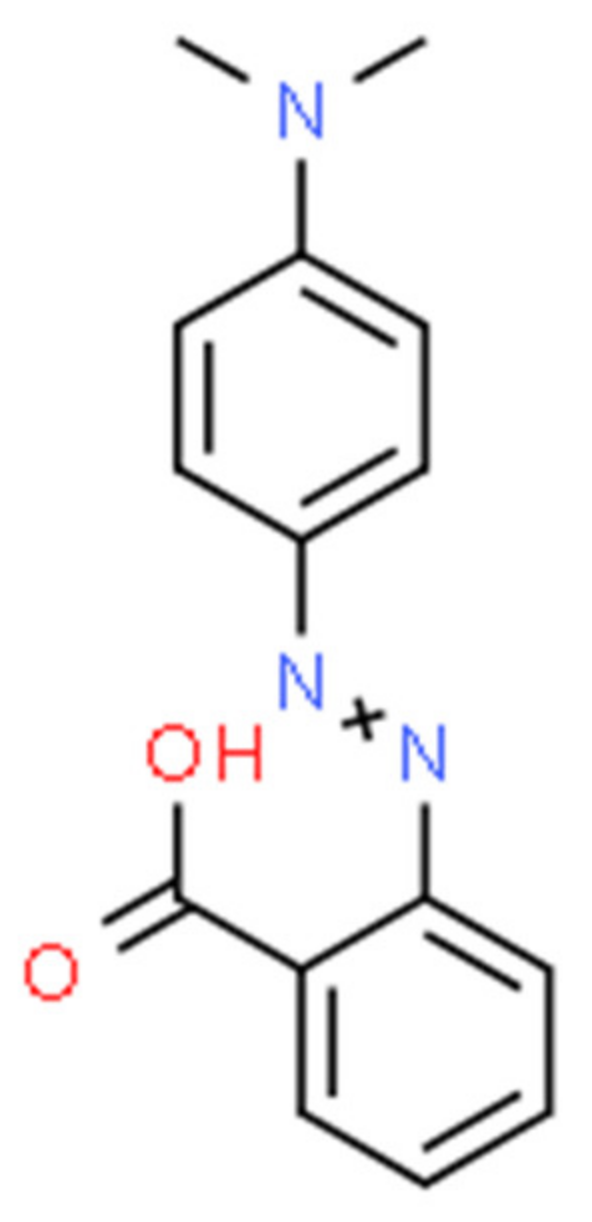
:1. Introduction
2. Materials and Methods
2.1. Soil Sample Collection
2.2. Microbial Screening and Isolation
2.3. Decolorization Study
2.4. Microbial Fuel Cell Design and Bacterial Inoculation
2.5. Anolyte pH and Substrate Concentration Optimization
2.5.1. Effect of Anolyte pH
2.5.2. Effect of Substrate Concentration
3. Results
3.1. Isolation and Screening of B1 and B2
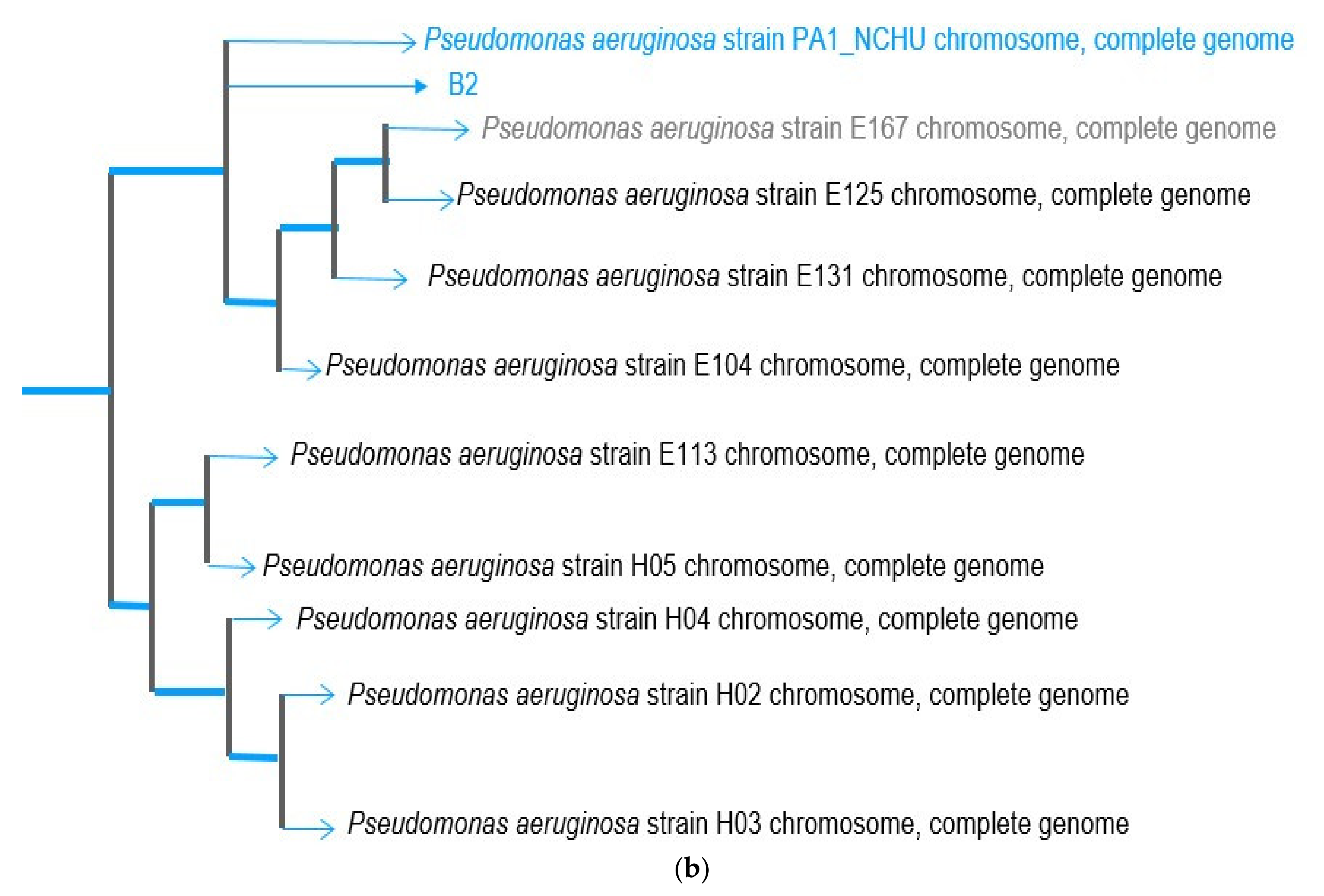
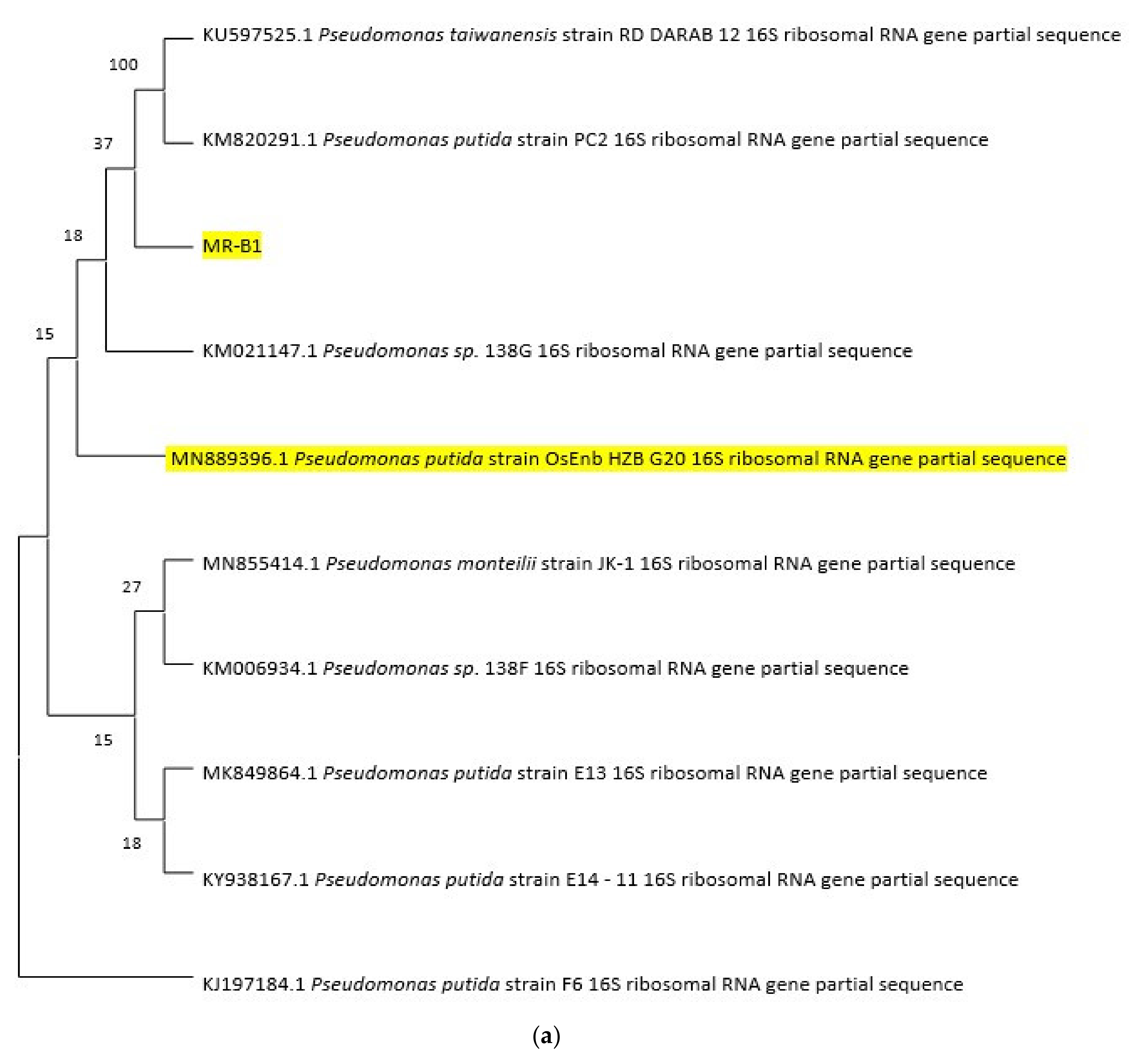
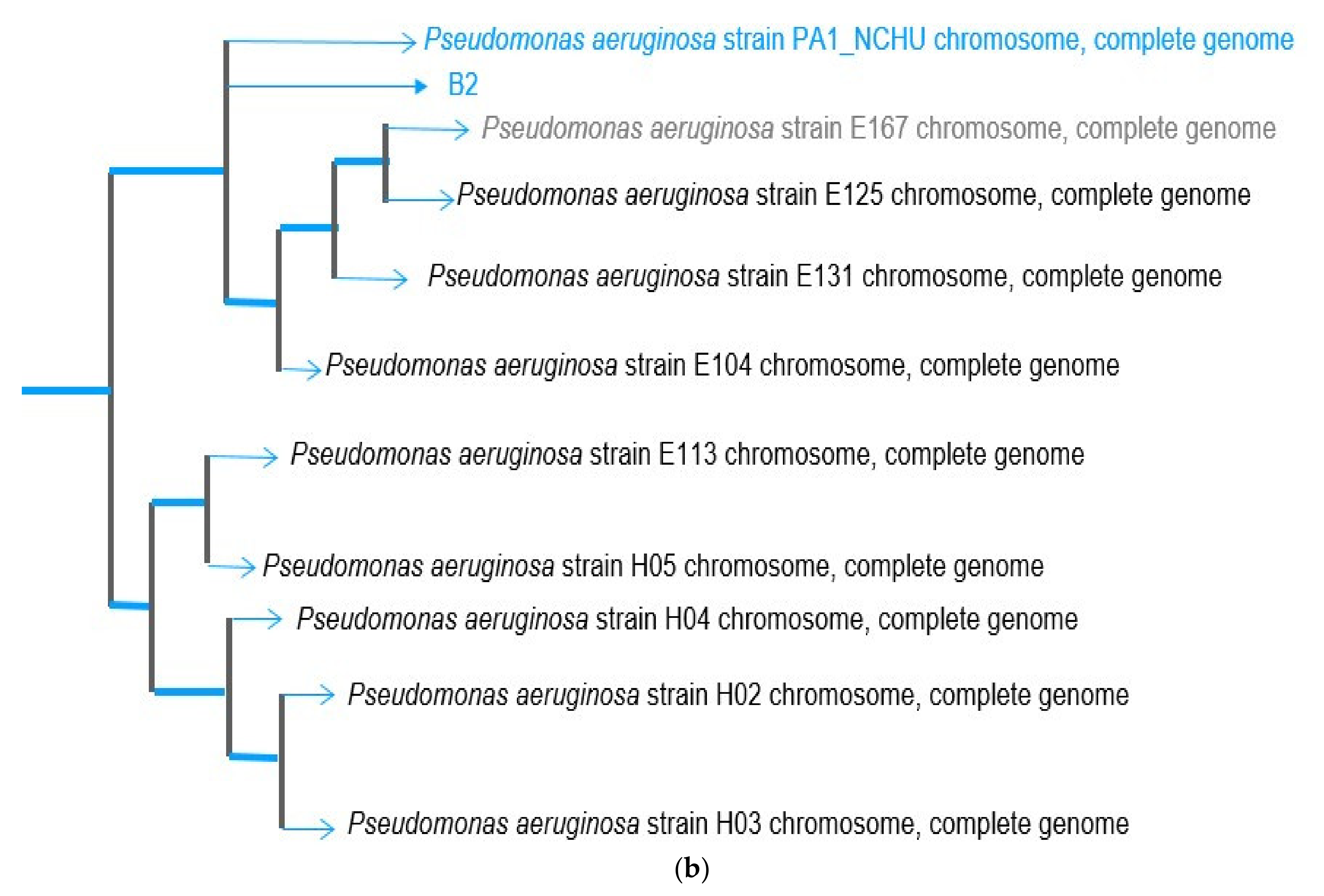
3.2. Genome Sequence of Psudomonas putida OsEnB_HZB_G20 and Pseudomonas aeruginosa PA1_NCHU
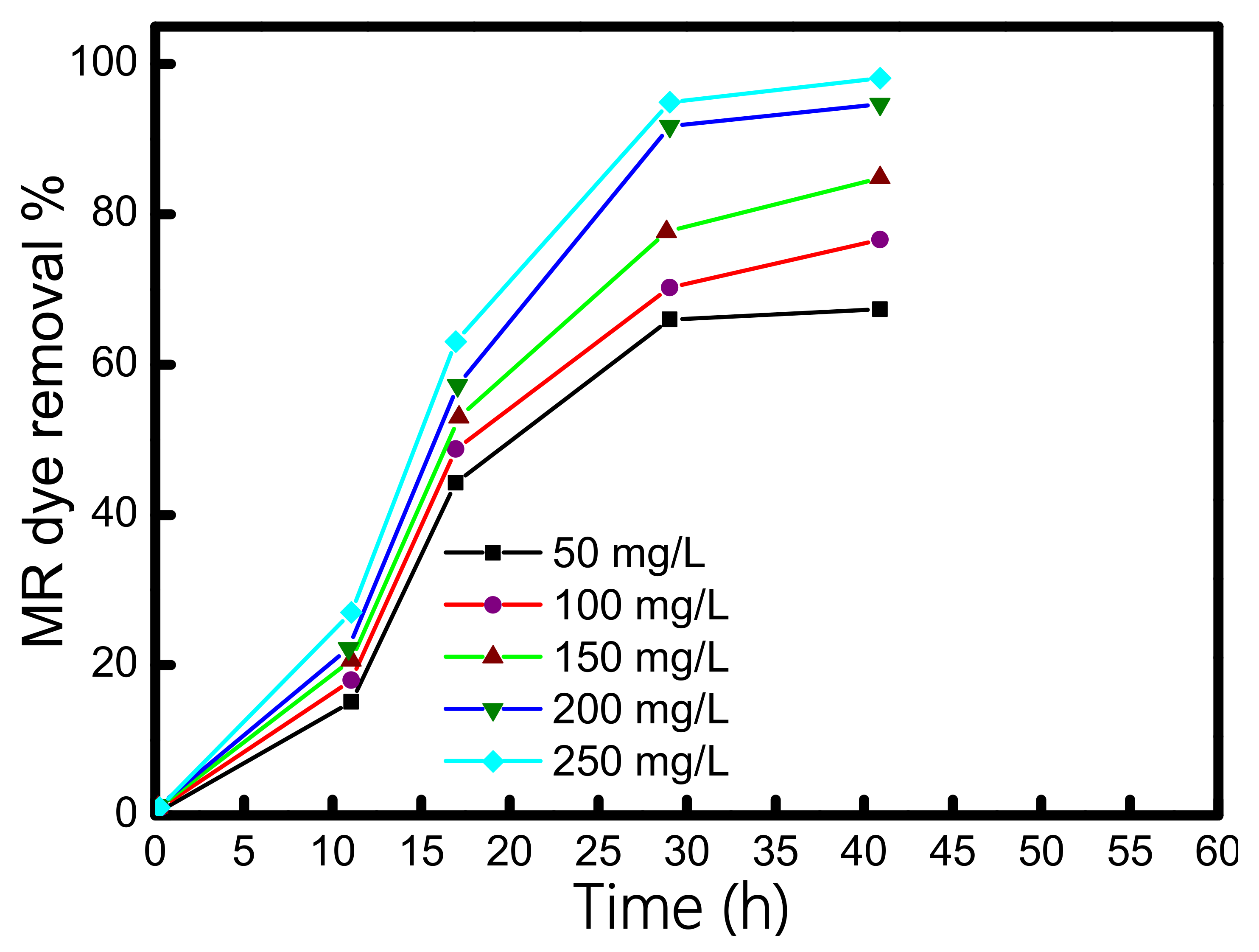
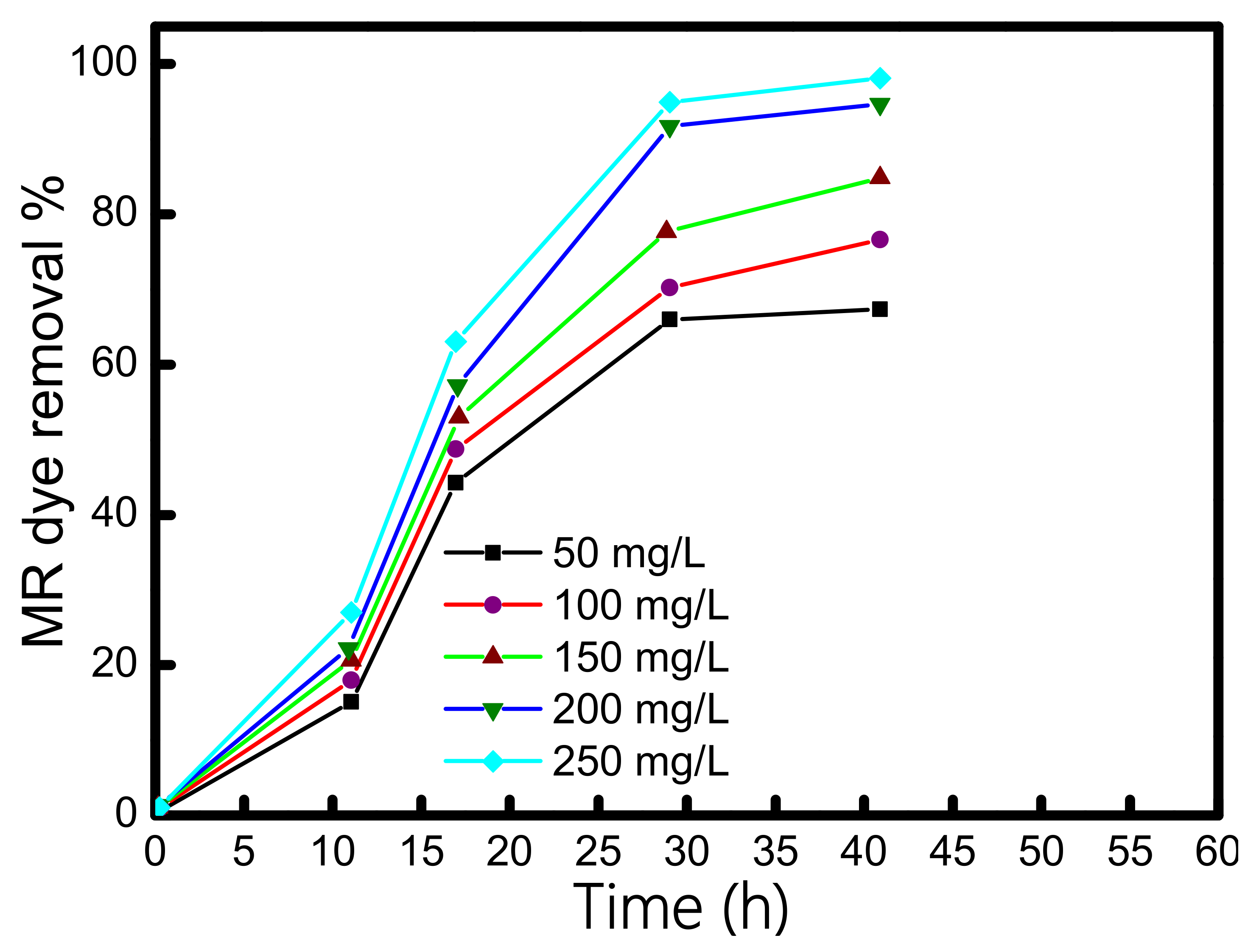
3.3. Dye Concentration Optimization
3.4. Methyl Red Dye Degradation in sMFCs
3.5. Effect of anolyte pH
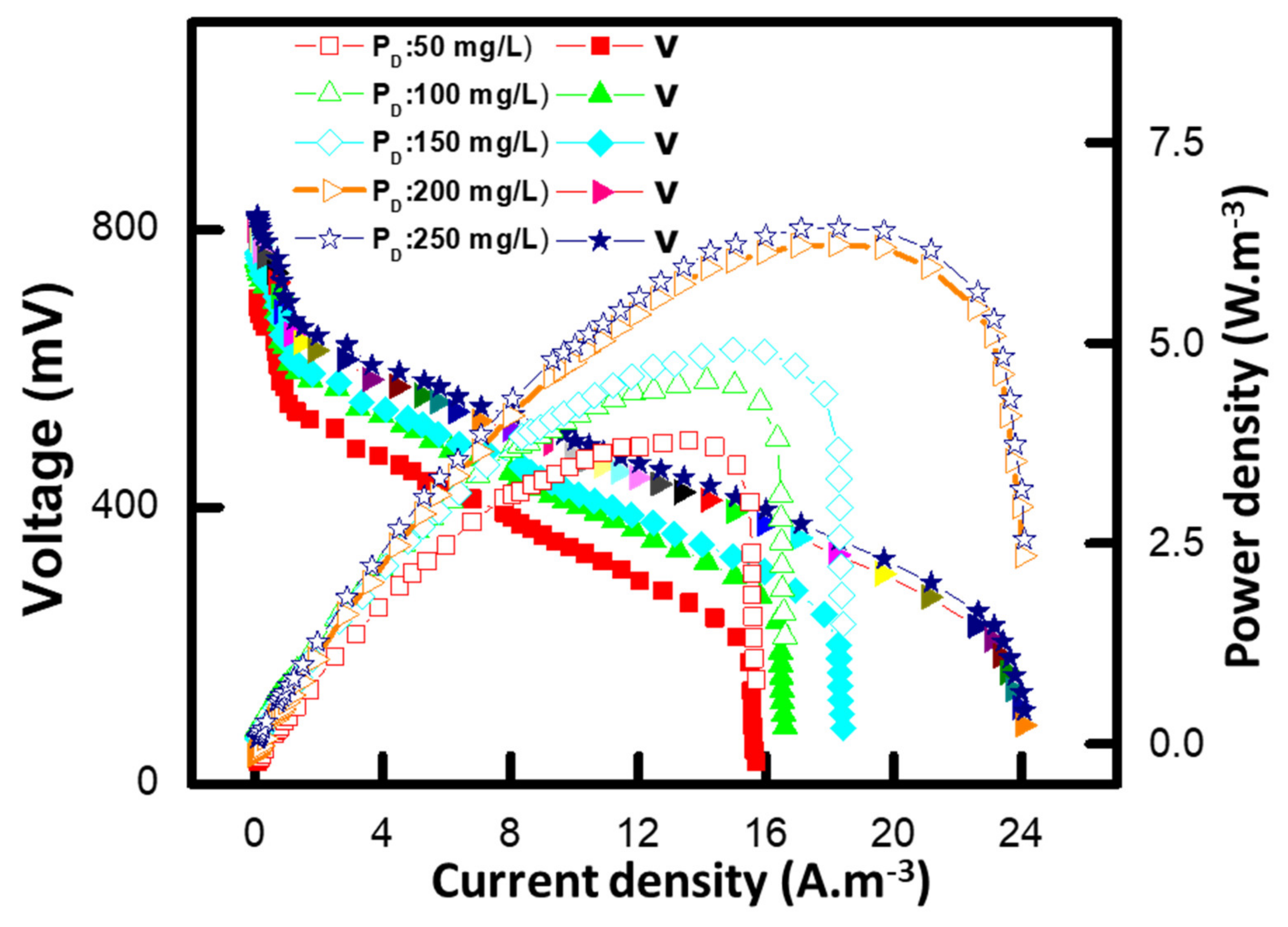
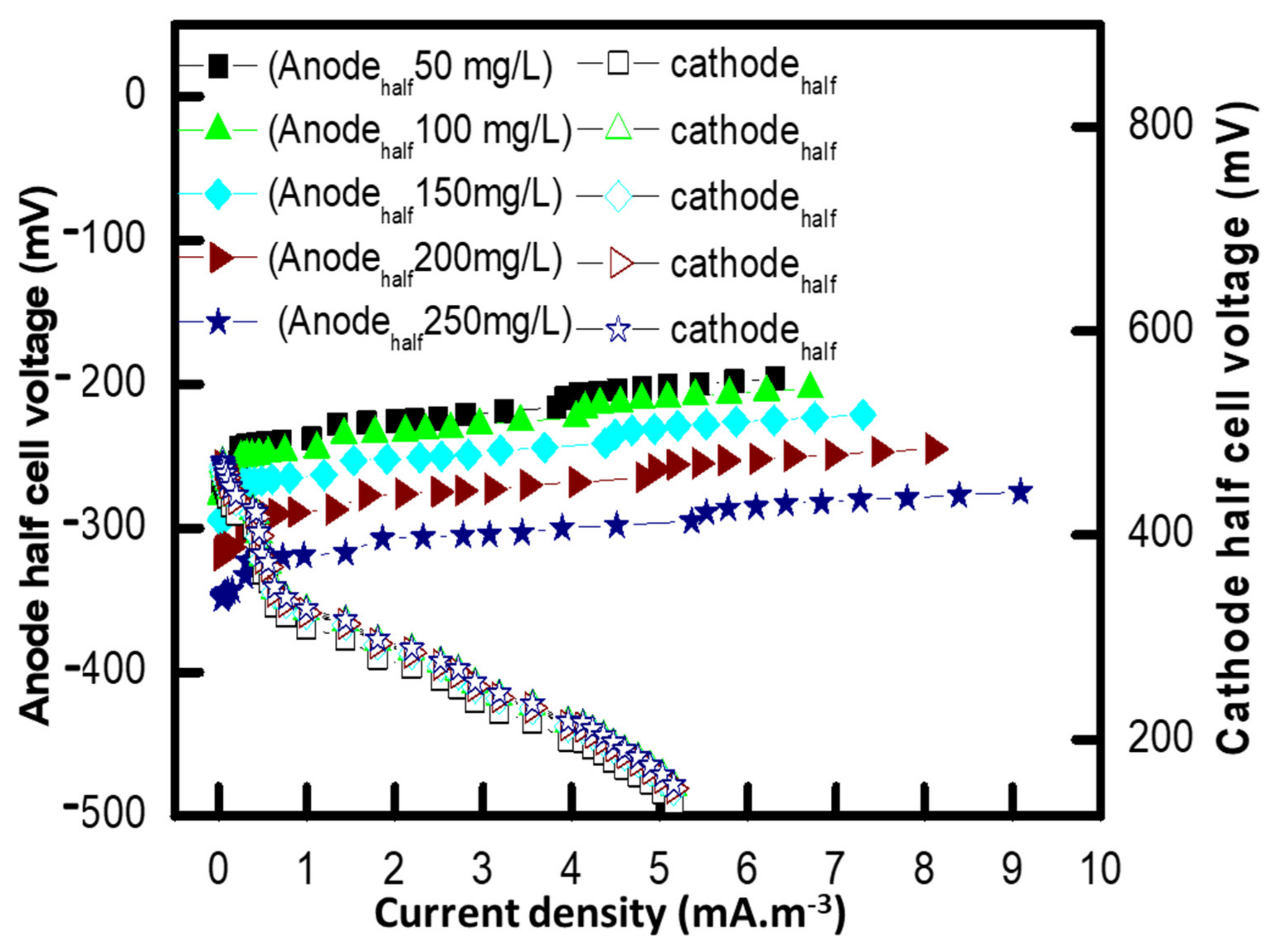
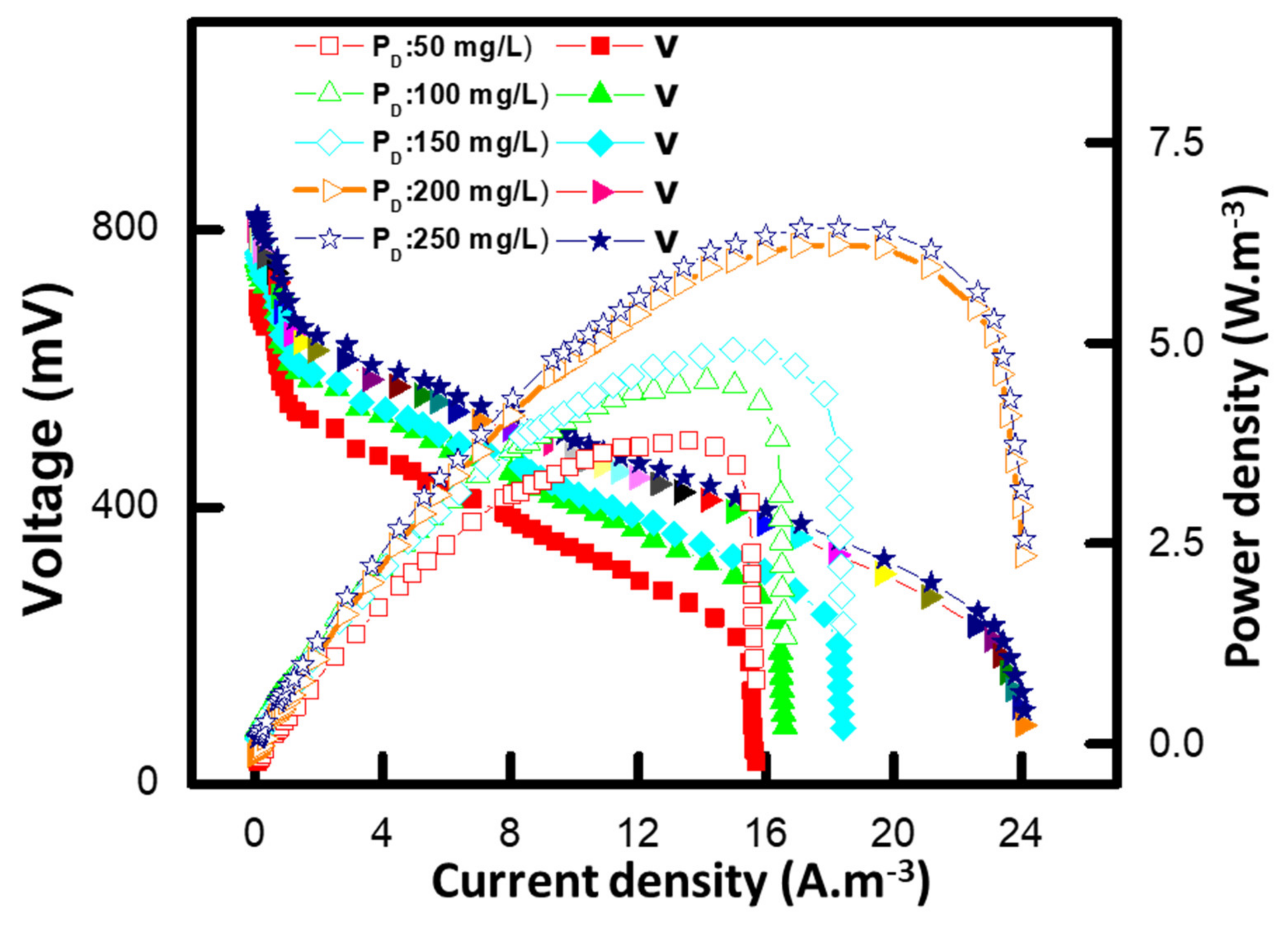
3.6. Polarization Study of MFCs
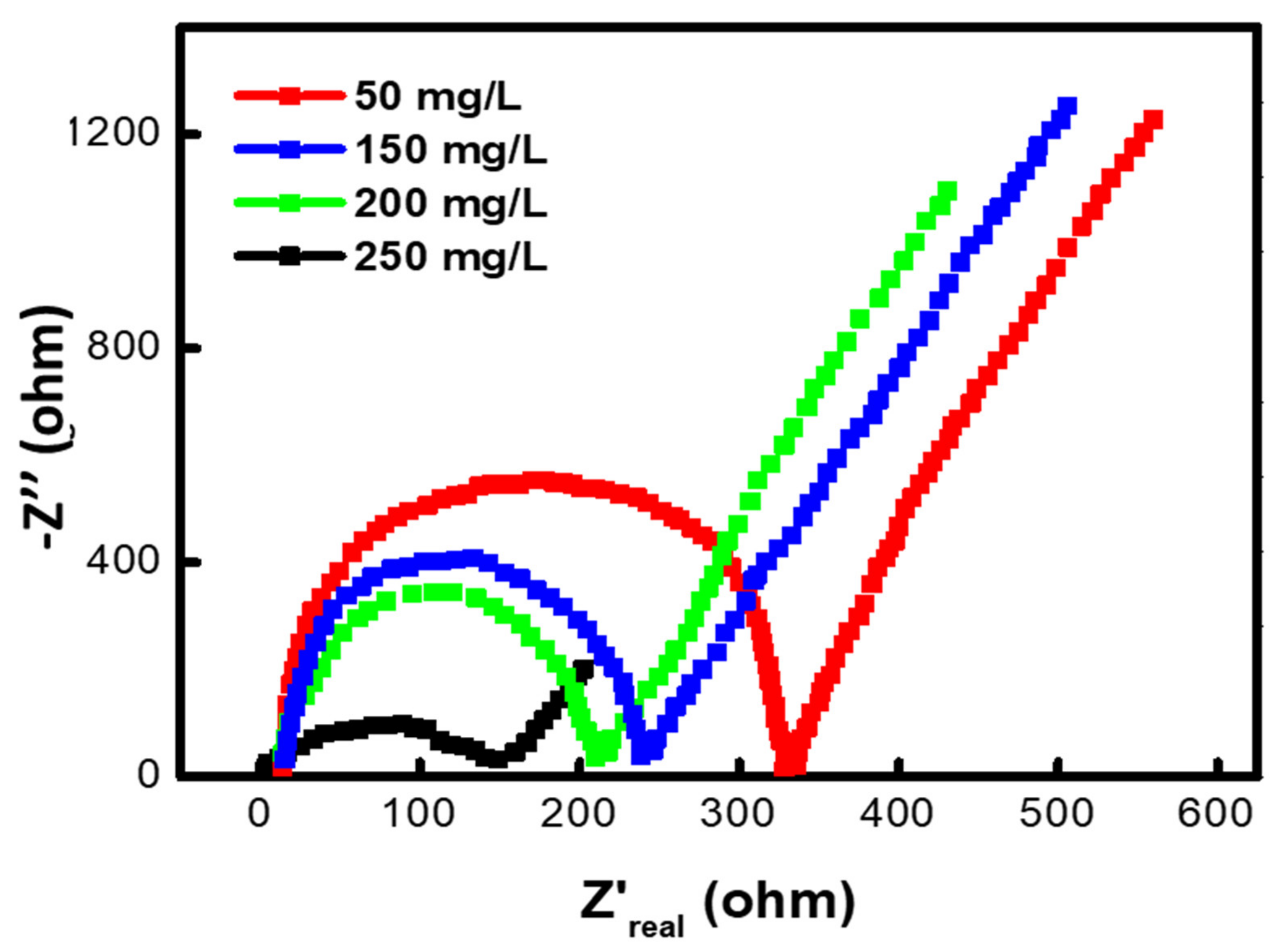
3.7. Effect of Different Methyl Red Concentrations on the Internal Resistance of MFC
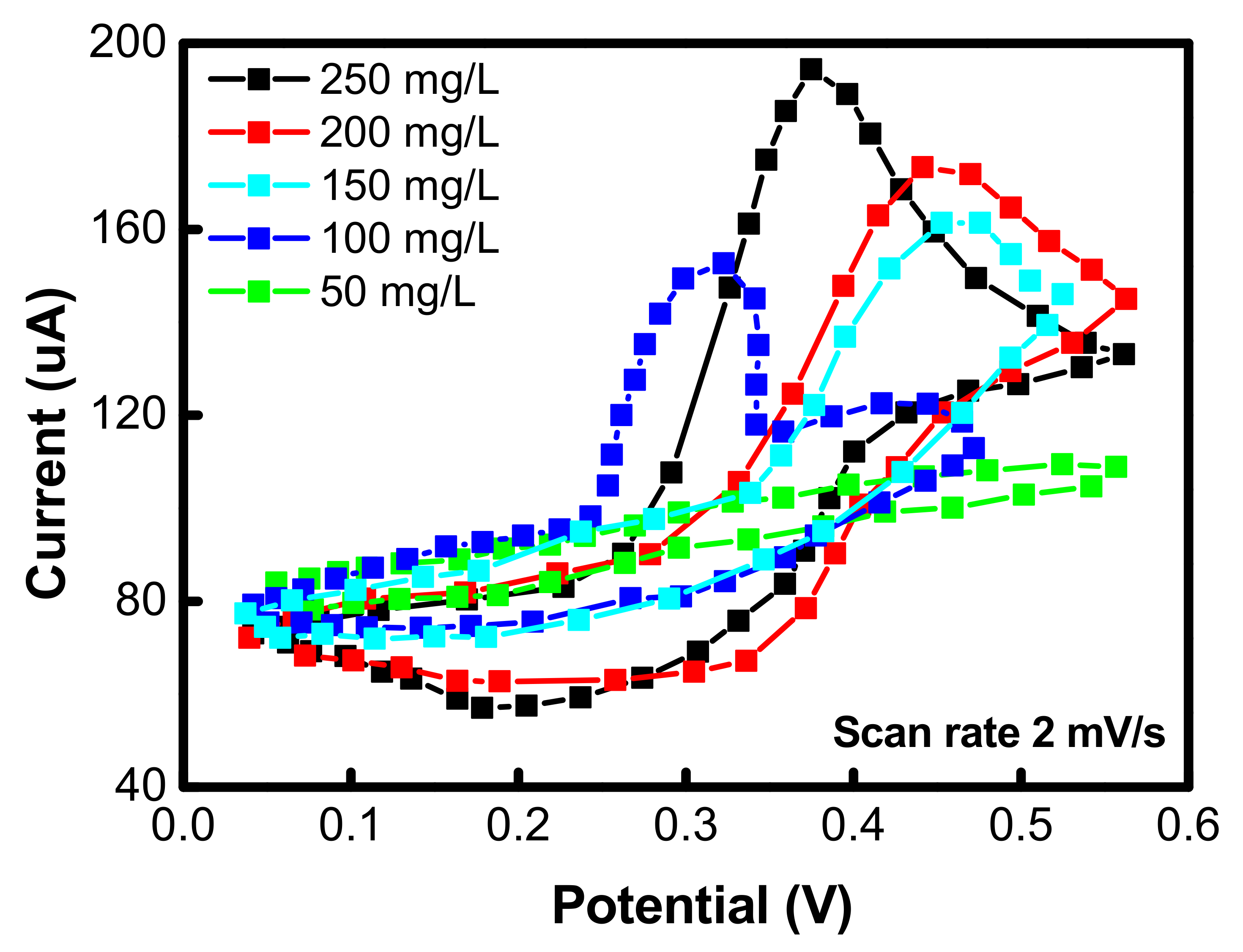
4. Cyclic Voltammetry Analyses
5. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Degrading Microorganism | Reference |
|---|---|
| Saccharomyces cerevisiae ATCC9763 | [53] |
| Galactomyces geotrichum MTCC 1360 | [54] |
| Sphingomonas paucimobilis | [55] |
| Rhodococcus strain UCC 0016 | [9] |
| Pseudomonas aeruginosa | [56] |
| Bacillus sp. strain UN2 | [57] |
| Enterobacter asburiae strain JCM6051 | [58] |
| Klebsiella sp. strain Y3 | [59] |
| Bacillus stratosphericus SCA1007 | [25] |
| Enterobacter agglomerans | [60] |
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wainwright, M. Dyes in the development of drugs and pharmaceuticals. Dye. Pigment. 2008, 76, 582–589. [Google Scholar] [CrossRef]
- Banat, I.M.; Nigam, P.; Singh, D.; Marchant, R. Microbial decolorization of textile-dyecontaining effluents: A review. Bioresour. Technol. 1996, 58, 217–227. [Google Scholar] [CrossRef]
- Khan, M.D.; Abdulateif, H.; Ismail, I.M.; Sabir, S.; Khan, M.Z. Bioelectricity Generation and Bioremediation of an Azo-Dye in a Microbial Fuel Cell Coupled Activated Sludge Process. PLoS ONE 2015, 10, e0138448. [Google Scholar] [CrossRef] [PubMed]
- Molina-Guijarro, J.M.; Pérez, J.; Muñoz-Dorado, J.; Guillén, F.; Moya, R.; Hernández, M.; Arias, M.E. Detoxification of azo dyes by a novel pH-versatile, salt-resistant laccase from Streptomyces ipomoea. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2009, 12, 13–21. [Google Scholar]
- Murali, V.; Ong, S.-A.; Ho, L.-N.; Wong, Y.-S.; Hamidin, N. Comprehensive Review and Compilation of Treatment for Azo Dyes Using Microbial Fuel Cells. Water Environ. Res. 2013, 85, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Nimje, V.R.; Chen, C.-Y.; Chen, C.-C.; Jean, J.-S.; Reddy, A.S.; Fan, C.-W.; Pan, K.-Y.; Liu, H.-T.; Chen, J.-L. Stable and high energy generation by a strain of Bacillus subtilis in a microbial fuel cell. J. Power Sources 2009, 190, 258–263. [Google Scholar] [CrossRef]
- Doble, M.; Kumar, A. Chapter 10—Degradation of Dyes. In Biotreatment of Industrial Effluents; Doble, M., Kumar, A., Eds.; Butterworth-Heinemann: Burlington, NJ, USA, 2005; pp. 111–122. ISBN 978-0-7506-7838-4. [Google Scholar]
- Sudha, M.; Arul, S.; Gopal, S.; Natesan, S. Microbial degradation of Azo Dyes: A review. Int. J. Curr. Microbiol. App. Sci. 2014, 3, 670–690. [Google Scholar]
- Maniyam, M.N.; Ibrahim, A.L.; Cass, A.E.G. Decolourization and biodegradation of azo dye methyl red by Rhodococcus strain UCC 0016. Environ. Technol. 2020, 41, 71–85. [Google Scholar] [CrossRef]
- Litter, M.I. Introduction to Photochemical Advanced Oxidation Processes for Water Treatment. In Environmental Photochemistry Part II; Boule, P., Bahnemann, D.W., Robertson, P.K.J., Eds.; The Handbook of Environmental Chemistry; Springer: Berlin/Heidelberg, Germany, 2005; pp. 325–366. ISBN 978-3-540-31494-3. [Google Scholar]
- Sehar, S.; Rasool, T.; Syed, H.M.; Mir, M.A.; Naz, I.; Rehman, A.; Shah, M.S.; Akhter, M.S.; Mahmood, Q.; Younis, A. Recent advances in biodecolorization and biodegradation of environmental threatening textile finishing dyes. 3 Biotech 2022, 12, 186. [Google Scholar] [CrossRef]
- Galai, S.; Pérez de los Ríos, A.; Hernández-Fernández, F.J.; Kacem, S.H.; Ramírez, F.M.; Quesada-Medina, J. Microbial Fuel Cell Application for Azoic Dye Decolorization with Simultaneous Bioenergy Production Using Stenotrophomonas sp. Chem. Eng. Technol. 2015, 38, 1511–1518. [Google Scholar] [CrossRef]
- Guo, W.; Feng, J.; Song, H.; Sun, J. Simultaneous bioelectricity generation and decolorization of methyl orange in a two-chambered microbial fuel cell and bacterial diversity. Environ. Sci. Pollut. Res. 2014, 21, 11531–11540. [Google Scholar] [CrossRef] [PubMed]
- Hesselman, M.C.; Odoni, D.I.; Ryback, B.M.; de Groot, S.; van Heck, R.G.A.; Keijsers, J.; Kolkman, P.; Nieuwenhuijse, D.; van Nuland, Y.M.; Sebus, E.; et al. A Multi-Platform Flow Device for Microbial (Co-) Cultivation and Microscopic Analysis. PLoS ONE 2012, 7, e36982. [Google Scholar] [CrossRef] [PubMed]
- Hirose, A.; Kasai, T.; Koga, R.; Suzuki, Y.; Kouzuma, A.; Watanabe, K. Understanding and engineering electrochemically active bacteria for sustainable biotechnology. Bioresour. Bioprocess. 2019, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Logan, B.E.; Hamelers, B.; Rozendal, R.; Schröder, U.; Keller, J.; Freguia, S.; Aelterman, P.; Verstraete, W.; Rabaey, K. Microbial Fuel Cells: Methodology and Technology. Environ. Sci. Technol. 2006, 40, 5181–5192. [Google Scholar] [CrossRef] [PubMed]
- Méndez-García, C.; Peláez, A.I.; Mesa, V.; Sánchez, J.; Golyshina, O.V.; Ferrer, M. Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 2015, 6, 475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jadhav, D.A.; Park, S.-G.; Pandit, S.; Yang, E.; Ali Abdelkareem, M.; Jang, J.-K.; Chae, K.-J. Scalability of microbial electrochemical technologies: Applications and challenges. Bioresour. Technol. 2022, 345, 126498. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.; Savla, N.; Sonawane, J.M.; Sani, A.M.; Gupta, P.K.; Mathuriya, A.S.; Rai, A.K.; Jadhav, D.A.; Jung, S.P.; Prasad, R. Agricultural Waste and Wastewater as Feedstock for Bioelectricity Generation Using Microbial Fuel Cells: Recent Advances. Fermentation 2021, 7, 169. [Google Scholar] [CrossRef]
- Roy, A.S.; Sharma, A.; Thapa, B.S.; Pandit, S.; Lahiri, D.; Nag, M.; Sarkar, T.; Pati, S.; Ray, R.R.; Shariati, M.A.; et al. Microbiomics for enhancing electron transfer in an electrochemical system. Front. Microbiol. 2022, 13, 2663. [Google Scholar] [CrossRef] [PubMed]
- Menon, A.; Mathuriya, A.S.; Prasad, R.; Mahanta, A.; Kankeo, E.F.; Patil, R.; Pandit, S.; Wagmare, S. Microbial Electrochemical Dye Degradation: Present State of Art. In Recent Developments in Microbial Technologies; Prasad, R., Kumar, V., Singh, J., Upadhyaya, C.P., Eds.; Springer: Singapore, 2021; pp. 323–353. ISBN 9789811544392. [Google Scholar]
- Miran, W.; Jang, J.; Nawaz, M.; Shahzad, A.; Lee, D.S. Sulfate-reducing mixed communities with the ability to generate bioelectricity and degrade textile diazo dye in microbial fuel cells. J. Hazard. Mater. 2018, 352, 70–79. [Google Scholar] [CrossRef]
- Guo, G.; Hao, J.; Tian, F.; Liu, C.; Ding, K.; Zhang, C.; Yang, F.; Xu, J. Decolorization of Metanil Yellow G by a halophilic alkalithermophilic bacterial consortium. Bioresour. Technol. 2020, 316, 123923. [Google Scholar] [CrossRef]
- Akansha, K.; Chakraborty, D.; Sachan, S.G. Decolorization and degradation of methyl orange by Bacillus stratosphericus SCA1007. Biocatal. Agric. Biotechnol. 2019, 18, 101044. [Google Scholar] [CrossRef]
- Srivastava, A.; Dangi, L.K.; Kumar, S.; Rani, R. Microbial decolorization of Reactive Black 5 dye by Bacillus albus DD1 isolated from textile water effluent: Kinetic, thermodynamics & decolorization mechanism. Heliyon 2022, 8, e08834. [Google Scholar] [CrossRef]
- Diatraea Saccharalis Harbors Microorganisms that Can Affect Growth of Sugarcane Stalk-Dwelling Fungi. Available online: https://link.springer.com/article/10.1007/s42770-021-00647-4 (accessed on 2 October 2022).
- Endophytes as Source of Polypeptide and Polyketide Antibiotics Fengycin and Bacillaene: Process Development for Fengycin Production—ProQuest. Available online: https://www.proquest.com/openview/3bf992c2aaec8534ada923bfc7500c66/1?pq-origsite=gscholar&cbl=2026366&diss=y (accessed on 2 October 2022).
- Karim, M.E.; Dhar, K.; Hossain, M.T. Decolorization of Textile Reactive Dyes by Bacterial Monoculture and Consortium Screened from Textile Dyeing Effluent. J. Genet. Eng. Biotechnol. 2018, 16, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.; Khilari, S.; Roy, S.; Ghangrekar, M.M.; Pradhan, D.; Das, D. Reduction of start-up time through bioaugmentation process in microbial fuel cells using an isolate from dark fermentative spent media fed anode. Water Sci. Technol. J. Int. Assoc. Water Pollut. Res. 2015, 72, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Pandit, S.; Khilari, S.; Roy, S.; Pradhan, D.; Das, D. Improvement of power generation using Shewanella putrefaciens mediated bioanode in a single chambered microbial fuel cell: Effect of different anodic operating conditions. Bioresour. Technol. 2014, 166, 451–457. [Google Scholar] [CrossRef]
- Xinhua, T. Power Enhancement And Applications Of Microbial Fuel Cells. Ph.D. Thesis, Zhejiang University, Zhejiang, China, 2016. [Google Scholar]
- Tripathi, B.; Pandit, S.; Sharma, A.; Chauhan, S.; Mathuriya, A.S.; Dikshit, P.K.; Gupta, P.K.; Singh, R.C.; Sahni, M.; Pant, K.; et al. Modification of Graphite Sheet Anode with Iron (II, III) Oxide-Carbon Dots for Enhancing the Performance of Microbial Fuel Cell. Catalysts 2022, 12, 1040. [Google Scholar] [CrossRef]
- Jadhav, D.A.; Jain, S.C.; Ghangrekar, M.M. Cow’s urine as a yellow gold for bioelectricity generation in low cost clayware microbial fuel cell. Energy 2016, 113, 76–84. [Google Scholar] [CrossRef]
- Establishment of Regeneration and Transient Transformation Systems for Australian Native Resurrection Plant Tripogon Loliiformis. Available online: https://eprints.qut.edu.au/132682/ (accessed on 2 October 2022).
- Full Article: Biosynthesized α-MnO2-Based Polyaniline Binary Composite as Efficient Bioanode Catalyst for High-Performance Microbial Fuel Cell. Available online: https://www.tandfonline.com/doi/full/10.1080/26895293.2021.1934123 (accessed on 2 October 2022).
- Bajracharya, S.; Sharma, M.; Mohanakrishna, G.; Dominguez Benneton, X.; Strik, D.P.B.T.B.; Sarma, P.M.; Pant, D. An overview on emerging bioelectrochemical systems (BESs): Technology for sustainable electricity, waste remediation, resource recovery, chemical production and beyond. Renew. Energy 2016, 98, 153–170. [Google Scholar] [CrossRef]
- Bhatt, P. Smart Bioremediation Technologies: Microbial Enzymes; Academic Press: Cambridge, MA, USA, 2019; ISBN 978-0-12-818308-3. [Google Scholar]
- Jadhav, D.A.; Ghadge, A.N.; Mondal, D.; Ghangrekar, M.M. Comparison of oxygen and hypochlorite as cathodic electron acceptor in microbial fuel cells. Bioresour. Technol. 2014, 154, 330–335. [Google Scholar] [CrossRef]
- Microbial Fuel Cells: An Overview of Current Technology—ScienceDirect. Available online: https://www.sciencedirect.com/science/article/pii/S1364032118306920 (accessed on 2 October 2022).
- Jayaprakash, J.; Parthasarathy, A.; Viraraghavan, R. Decolorization and degradation of monoazo and diazo dyes in Pseudomonas catalyzed microbial fuel cell. Environ. Prog. Sustain. Energy 2016, 35, 1623–1628. [Google Scholar] [CrossRef]
- Fernando, E.; Keshavarz, T.; Kyazze, G. Simultaneous co-metabolic decolourisation of azo dye mixtures and bio-electricity generation under thermophillic (50 °C) and saline conditions by an adapted anaerobic mixed culture in microbial fuel cells. Bioresour. Technol. 2013, 127, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Osman, M.E.; Khattab, O.-K. Acid Black 172 Dye Decolorization and Bioelectricity Generation by Microbial Fuel Cell with Filamentous Fungi on Anode. Biosci. Biotechnol. Res. Asia 2018, 15, 981–986. [Google Scholar] [CrossRef]
- Simões, M.F.; Maiorano, A.E.; dos Santos, J.G.; Peixoto, L.; de Souza, R.F.B.; Neto, A.O.; Brito, A.G.; Ottoni, C.A. Microbial fuel cell-induced production of fungal laccase to degrade the anthraquinone dye Remazol Brilliant Blue R. Environ. Chem. Lett. 2019, 17, 1413–1420. [Google Scholar] [CrossRef] [Green Version]
- Ikram, M.; Naeem, M.; Zahoor, M.; Rahim, A.; Hanafiah, M.M.; Oyekanmi, A.A.; Shah, A.B.; Mahnashi, M.H.; Al Ali, A.; Jalal, N.A.; et al. Biodegradation of Azo Dye Methyl Red by Pseudomonas aeruginosa: Optimization of Process Conditions. Int. J. Environ. Res. Public. Health 2022, 19, 9962. [Google Scholar] [CrossRef]
- Razika, B.; Abbes, B.; Messaoud, C.; Soufi, K. Phenol and Benzoic Acid Degradation by Pseudomonas aeruginosa. J. Water Resour. Prot. 2010, 2, 788–791. [Google Scholar] [CrossRef] [Green Version]
- Kahraman, H.; Geckil, H. Degradation of Benzene, Toluene and Xylene by Pseudomonas aeruginosa Engineered with the Vitreoscilla Hemoglobin Gene. Eng. Life Sci. 2005, 5, 363–368. [Google Scholar] [CrossRef]
- Mohanty, S.S.; Kumar, A. Enhanced degradation of anthraquinone dyes by microbial monoculture and developed consortium through the production of specific enzymes. Sci. Rep. 2021, 11, 7678. [Google Scholar] [CrossRef]
- Vatandoostarani, S.; Bagheri Lotfabad, T.; Heidarinasab, A.; Yaghmaei, S. Degradation of azo dye methyl red by Saccharomyces cerevisiae ATCC 9763. Int. Biodeterior. Biodegrad. 2017, 125, 62–72. [Google Scholar] [CrossRef]
- Jadhav, S.U.; Kalme, S.D.; Govindwar, S.P. Biodegradation of Methyl red by Galactomyces geotrichum MTCC 1360. Int. Biodeterior. Biodegrad. 2008, 62, 135–142. [Google Scholar] [CrossRef]
- Ayed, L.; Mahdhi, A.; Cheref, A.; Bakhrouf, A. Decolorization and degradation of azo dye Methyl Red by an isolated Sphingomonas paucimobilis: Biotoxicity and metabolites characterization. Desalination 2011, 274, 272–277. [Google Scholar] [CrossRef]
- Jadhav, D.A.; Ghadge, A.N.; Ghangrekar, M.M. Simultaneous organic matter removal and disinfection of wastewater with enhanced power generation in microbial fuel cell. Bioresour. Technol. 2014, 163, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Sun, P.-F.; Du, L.-N.; Wang, G.; Jia, X.-M.; Zhao, Y.-H. Biodegradation of methyl red by Bacillus sp. strain UN2: Decolorization capacity, metabolites characterization, and enzyme analysis. Environ. Sci. Pollut. Res. 2014, 21, 6136–6145. [Google Scholar] [CrossRef] [PubMed]
- Swati; Singh, P. Bioremediation of hazardous azo dye methyl red by a newly isolated Enterobacter asburiae strain JCM6051 from industrial effluent of Uttarakhand regions. J. Appl. Biol. Biotechnol. 2022, 10, 64–72. [Google Scholar] [CrossRef]
- Cui, D.; Li, G.; Zhao, M.; Han, S. Decolourization of azo dyes by a newly isolated Klebsiella sp. strain Y3, and effects of various factors on biodegradation. Biotechnol. Biotechnol. Equip. 2014, 28, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Moutaouakkil, A.; Zeroual, Y.; Zohra Dzayri, F.; Talbi, M.; Lee, K.; Blaghen, M. Purification and partial characterization of azoreductase from Enterobacter agglomerans. Arch. Biochem. Biophys. 2003, 413, 139–146. [Google Scholar] [CrossRef]
- Jadhav, D.A.; Ghangrekar, M.M. Optimising the proportion of pure and mixed culture in inoculum to enhance the performance of microbial fuel cells. Int. J. Environ. Technol. Manag. 2020, 23, 50–67. [Google Scholar] [CrossRef]
- Gunaseelan, K.; Jadhav, D.A.; Gajalakshmi, S.; Pant, D. Blending of microbial inocula: An effective strategy for performance enhancement of clayware Biophotovoltaics microbial fuel cells. Bioresour. Technol. 2021, 323, 124564. [Google Scholar] [CrossRef]
- Jadhav, D.A.; Chendake, A.D.; Vinayak, V.; Atabani, A.; Abdelkareem, M.A.; Chae, K.J. Scale-up of the bioelectrochemical system: Strategic perspectives and normalization of performance indices. Bioresour. Technol. 2022, 363, 127935. [Google Scholar] [CrossRef]
- Yadav, A.; Jadhav, D.A.; Ghangrekar, M.M.; Mitra, A. Effectiveness of constructed wetland integrated with microbial fuel cell for domestic wastewater treatment and to facilitate power generation. Environ. Sci. Pollut. Res. 2022, 29, 51117–51129. [Google Scholar] [CrossRef]
- Jadhav, D.A.; Park, S.G.; Eisa, T.; Mungray, A.K.; Madenli, E.C.; Olabi, A.G.; Abdelkareem, M.A.; Chae, K.J. Current outlook towards feasibility and sustainability of ceramic membranes for practical scalable applications of microbial fuel cells. Renew. Sustain. Energy Rev. 2022, 167, 112769. [Google Scholar] [CrossRef]
- Mathuriya, A.S.; Kaur, A.; Gupta, P.K.; Pandit, S.; Jadhav, D.A. Potential of microbial fuel cells for wastewater treatment. In Bioremediation, Nutrients, and Other Valuable Product Recovery; Elsevier: Amsterdam, The Netherlands, 2021; pp. 115–124. [Google Scholar]
- Jadhav, D.A.; Ghangrekar, M.M. Effect of salinity, acetate addition and alteration of sediment onperformance of benthic microbial fuel cells. Recent Adv. Bio-Energy Res. 2014, 3, 394–408. [Google Scholar]
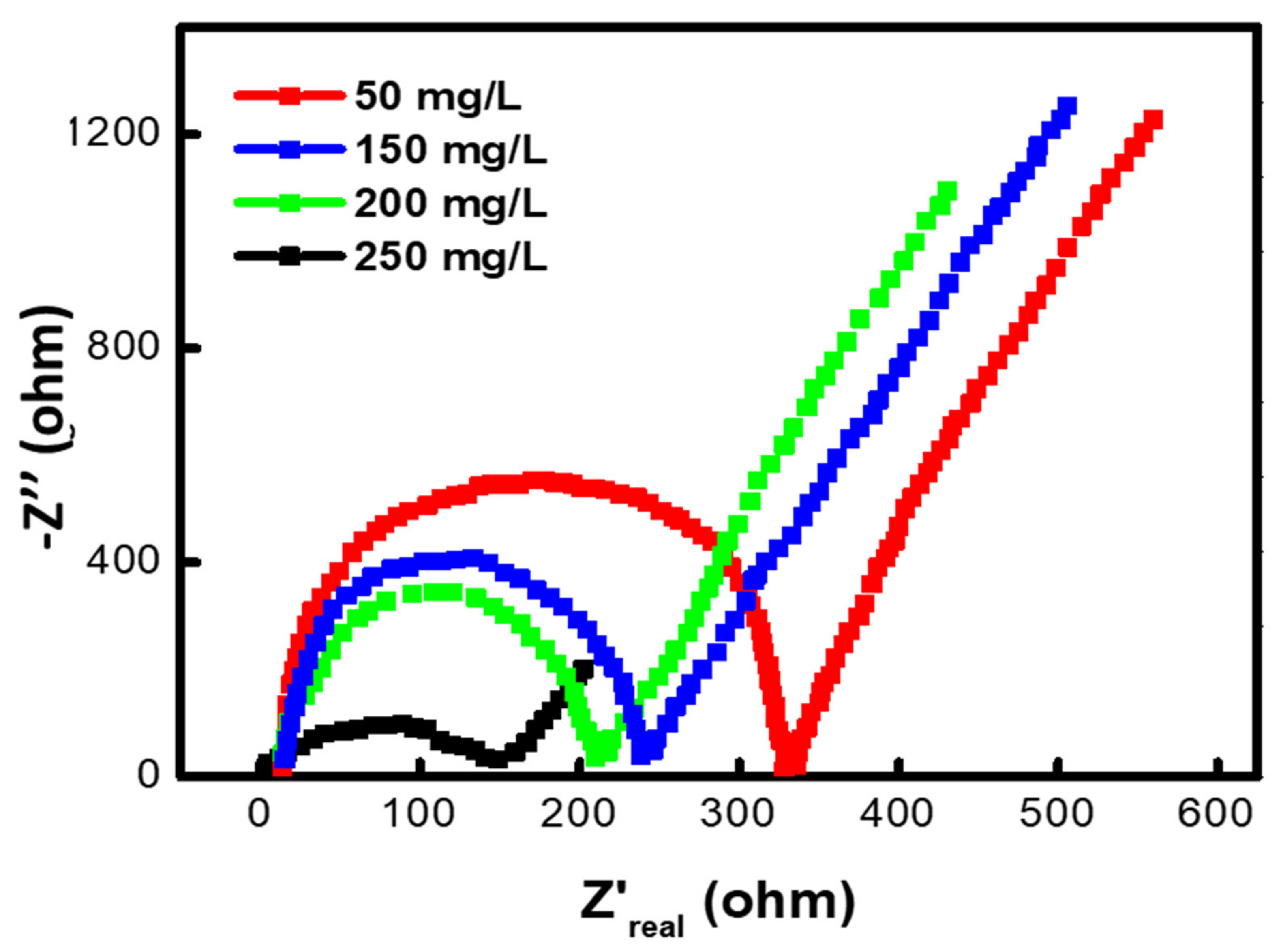
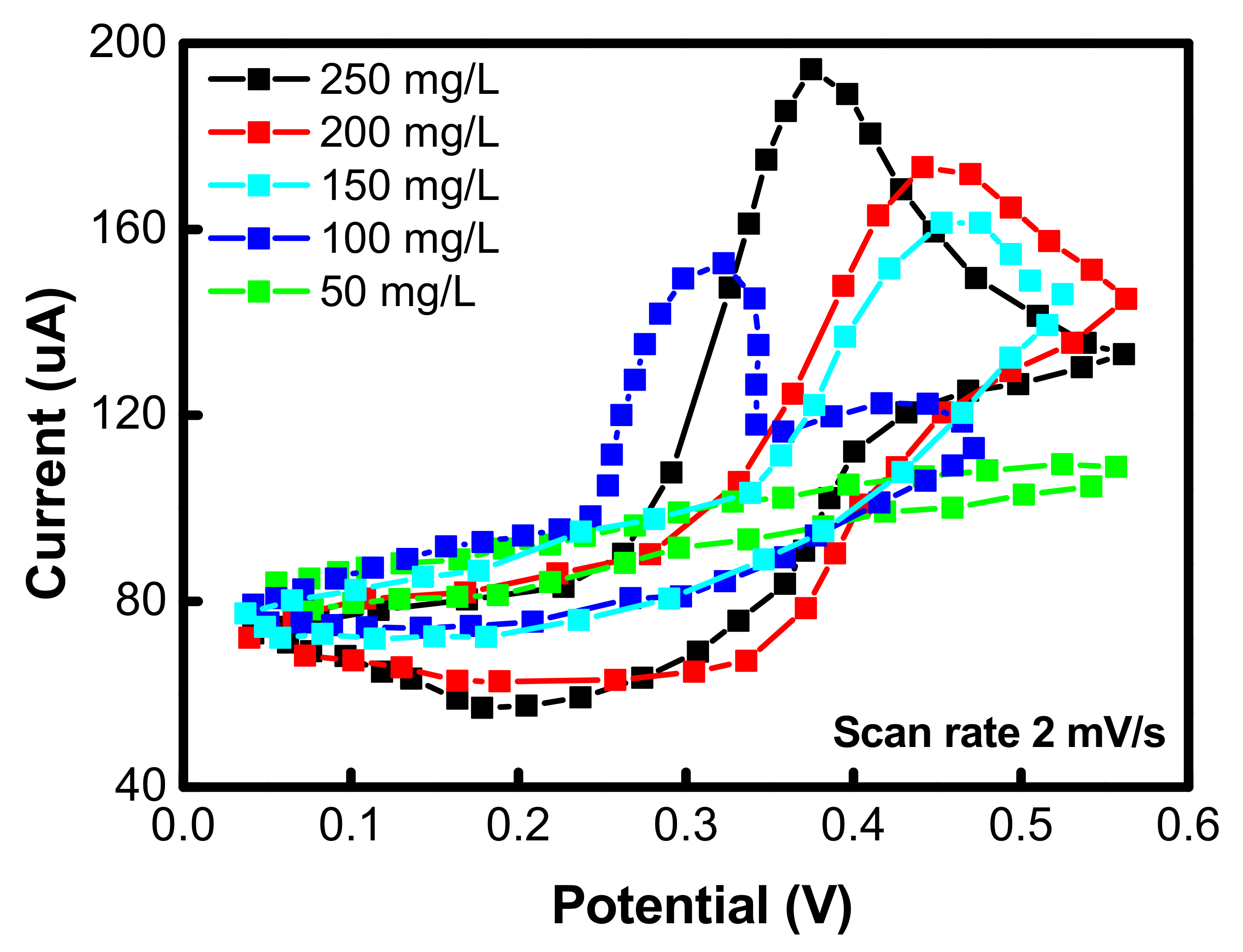
| Factors | Range Investigated | Operational Range | Optimized Factor |
|---|---|---|---|
| Methyl red concentration | 50–300 mg | 50–250 mg | 250 mg |
| pH | 5–8 | 5–8 | 7.0 |
| Dye | Bacteria | Decolorization Efficiency | Power Density | Resistor | References |
|---|---|---|---|---|---|
| Reactive Blue | Pseudomonas aeruginosa | 90% | 2004 µW/m2 | 220 Ω | [44] |
| Reactive red | Pseudomonas aeruginosa | 74% | 4100 µW/m2 | 220 Ω | [44] |
| Congo red | Pseudomonas aeruginosa | 80% | 586 µW/m2 | 220 Ω | [44] |
| Methyl orange | Pseudomonas aeruginosa | 94% | 4100 µW/m3 | 220 Ω | [44] |
| Acid orange | Shewanella oeidensis | 93% | - | - | [45] |
| Acid black 172 | Trichoderma | 85% | 129 mA/m2 | NA | [46] |
| Acid black 172 | Aspergillus | 88% | 133 mA/m2 | NA | [46] |
| Remazol brilliant blue R | Pleurotus ostreatus | 80–90% | 180.5 mW/m2 | NA | [47] |
| Methyl red | Pseudomonasputida + Pseudomonas aeruginosa | 95% | 7.3 W/m3 | 50 Ω | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, K.; Pandit, S.; Mathuriya, A.S.; Gupta, P.K.; Pant, K.; Jadhav, D.A. Microbial Electrochemical Treatment of Methyl Red Dye Degradation Using Co-Culture Method. Water 2023, 15, 56. https://doi.org/10.3390/w15010056
Sharma K, Pandit S, Mathuriya AS, Gupta PK, Pant K, Jadhav DA. Microbial Electrochemical Treatment of Methyl Red Dye Degradation Using Co-Culture Method. Water. 2023; 15(1):56. https://doi.org/10.3390/w15010056
Chicago/Turabian StyleSharma, Kalpana, Soumya Pandit, Abhilasha Singh Mathuriya, Piyush Kumar Gupta, Kumud Pant, and Dipak A. Jadhav. 2023. "Microbial Electrochemical Treatment of Methyl Red Dye Degradation Using Co-Culture Method" Water 15, no. 1: 56. https://doi.org/10.3390/w15010056
APA StyleSharma, K., Pandit, S., Mathuriya, A. S., Gupta, P. K., Pant, K., & Jadhav, D. A. (2023). Microbial Electrochemical Treatment of Methyl Red Dye Degradation Using Co-Culture Method. Water, 15(1), 56. https://doi.org/10.3390/w15010056