
Comparison of Marine Ecosystems of Haizhou Bay and Lvsi Fishing Ground in China Based on the Ecopath Model

Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Study Area
2.1.1. Haizhou Bay
2.1.2. Lvsi Fishing Ground
2.2. Ecopath Model
2.3. Trophic Network Analysis

| Indicators | Acronym | Units | Reference | |
|---|---|---|---|---|
| Statistics and flows | Total consumption | Q | t km−2 year−1 | [32,33] |
| Total exports | Ex | t km−2 year−1 | ||
| Flows to detritus | FD | t km−2 year−1 | ||
| Total respiration | R | t km−2 year−1 | ||
| Total production | P | t km−2 year−1 | ||
| Total system throughput | TST | t km−2 year−1 | ||
| Total system throughput/TST | Q/TST | – | ||
| Total exports/TST | Ex/TST | – | ||
| Flows to detritus/TST | FD/TST | – | ||
| Total respiration/TST | R/TST | – | ||
| Total production/TST | P/TST | – | ||
| Total biomass/TST | B/TST | – | ||
| Ecosystem maturity status | Primary production/total respiration | PP/R | – | [32,34,35,36,37] |
| Primary production/total biomass | PP/B | – | ||
| Finn’s cycling index | FCI | % | ||
| Finn’s mean path length | FMPL | % | ||
| System omnivory index | SOI | – | ||
| Connectance index | CI | – | ||
| Ascendency/capacity | A/C | % | ||
| Overhead/capacity | O/C | % | ||
| Shannon diversity index | H | – | ||
| Ecosystem efficiency | Mean transfer efficiency | MTE | % | [38] |
| MTE from primary production | MTEpp | % | ||
| MTE from detritus | MTEd | % | ||
| Proportion of total flow originating from detritus | PEED | – | ||
| Trophic indices | Mean trophic level of the community | MTL | – | [39] |
| Marine trophic index | MTL0 | – | ||
| High trophic index | HTI | % | ||
| Apex predator indicator | API | % | ||
2.4. MTI
2.5. Keystoneness
3. Results
3.1. Model Parameters (Inputs and Outputs)
| HZB | Group Name | Catch (t/km2) | Biomass (t/km2) | P/B (/Year) | Q/B (/Year) | TLs | EE | P/Q (/year) |
|---|---|---|---|---|---|---|---|---|
| 1 | Gobiidae | 0.016 [30] | 0.366 | 1.980 [30] | 12.117 * | 3.674 | 0.814 | 0.163 |
| 2 | Congers | 0.207 [30] | 0.0592 | 3.500 [23] | 12.000 * | 4.145 | 0.999 | 0.292 |
| 3 | Collichthys lucidus | 0.008 [6] | 0.134 | 1.658 [30] | 7.500 * | 3.269 | 0.989 | 0.221 |
| 4 | Larimichthys polyactis | 0.008 [6] | 0.114 | 1.658 [30] | 7.500 * | 3.316 | 0.965 | 0.221 |
| 5 | Other drumfish | 0.006 [5] | 0.0360 | 1.067 [23] | 6.833 * | 3.263 | 0.988 | 0.156 |
| 6 | Seabass | 0.003 [17] | 0.139 | 1.058 [30] | 4.000 * | 3.993 | 0.728 | 0.265 |
| 7 | Sardinella zunasi | 0.013 [30] | 0.313 | 3.000 [16] | 10.000 [16] | 3.221 | 0.892 | 0.300 |
| 8 | Anchovy | 0.013 [7] | 0.0333 | 3.000 [23] | 16.275 * | 2.861 | 0.848 | 0.184 |
| 9 | Filefish | 0.100 [30] | 0.0446 | 2.500 [30] | 10.800 * | 2.912 | 0.907 | 0.231 |
| 10 | Other pelagic fishes | 1.097 [30] | 0.439 | 2.600 [30] | 9.400 * | 2.685 | 0.964 | 0.277 |
| 11 | Benthic fishes | 0.035 [30] | 0.0530 | 1.460 [17] | 8.650 * | 2.471 | 0.809 | 0.169 |
| 12 | Small fishes | 0.505 [17] | 0.257 | 2.300 [17] | 23.214 * | 2.588 | 0.863 | 0.099 |
| 13 | Shrimps | 0.400 [17] | 1.101 | 8.000 [17] | 28.000 [17] | 2.569 | 0.997 | 0.286 |
| 14 | Crabs | 0.050 [17] | 1.853 | 3.500 [17] | 12.000 [17] | 2.867 | 0.937 | 0.292 |
| 15 | Cephalopods | 0.400 [17] | 0.139 | 3.000 [17] | 10.000 [17] | 2.471 | 0.963 | 0.300 |
| 16 | Benthos | 0.0435 | 1.570 [17] | 8.600 [17] | 2.510 | 0.891 | 0.183 | |
| 17 | Zooplankton | 2.195 | 40.000 [17] | 160.000 [30] | 2.176 | 0.913 | 0.250 | |
| 18 | Phytoplankton | 26.126 | 100.000 [17] | 1.000 | 0.064 | |||
| 19 | Detritus | 80.32 [30] | 1.000 | 0.065 | ||||
| LSFG | ||||||||
| 1 | Gobiidae | 0.016 [30] | 0.0268 | 1.980 [30] | 16.000 * | 2.569 | 0.783 | 0.124 |
| 2 | Congers | 0.207 [30] | 0.192 | 2.300 [30] | 12.000 * | 3.269 | 0.468 | 0.192 |
| 3 | Collichthys lucidus | 0.150 [30] | 0.102 | 1.658 [30] | 7.500 * | 2.894 | 0.928 | 0.221 |
| 4 | Larimichthys polyactis | 0.150 [30] | 0.353 | 1.658 [30] | 7.500 * | 2.793 | 0.934 | 0.221 |
| 5 | Other drumfish | 0.281 [30] | 0.123 | 2.392 [30] | 10.000 * | 2.736 | 0.969 | 0.239 |
| 6 | Seabass | 0.003 [30] | 0.614 | 1.058 [30] | 4.000 * | 3.063 | 0.360 | 0.265 |
| 7 | Sardinella zunasi | 0.013 [30] | 0.303 | 0.902 [30] | 47.600 * | 2.769 | 0.865 | 0.019 |
| 8 | Anchovy | 0.288 [30] | 0.0573 | 5.500 [30] | 25.000 * | 2.623 | 0.976 | 0.220 |
| 9 | Filefish | 0.100 [30] | 0.0472 | 2.500 [30] | 10.800 * | 2.635 | 0.847 | 0.231 |
| 10 | Other pelagic fishes | 1.097 [30] | 0.454 | 2.600 [30] | 9.400 * | 2.501 | 0.938 | 0.277 |
| 11 | Benthic fishes | 0.035 [30] | 0.0673 | 1.754 [30] | 9.525 * | 2.541 | 0.533 | 0.184 |
| 12 | Small fishes | 1.097 [30] | 0.0410 | 30.000 [30] | 106.300 * | 2.486 | 0.918 | 0.282 |
| 13 | Shrimps | 1.807 [30] | 0.583 | 7.570 [44] | 28.000 [44] | 2.544 | 0.975 | 0.270 |
| 14 | Crabs | 1.807 [30] | 0.906 | 2.120 [30] | 8.180 [44] | 2.502 | 0.976 | 0.259 |
| 15 | Cephalopods | 0. [30] | 0.0716 | 3.000 [30] | 10.000 [30] | 2.803 | 0.927 | 0.300 |
| 16 | Benthos | 0.00333 | 5.000 [30] | 20.000 [30] | 2.100 | 0.356 | 0.250 | |
| 17 | Zooplankton | 6.532 [30] | 40.000 [30] | 160.000 [30] | 2.000 | 0.129 | 0.250 | |
| 18 | Phytoplankton | 16.173 [30] | 200.000 [30] | 1.000 | 0.295 | |||
| 19 | Detritus | 83.320 [30] | 1.000 | 0.140 |
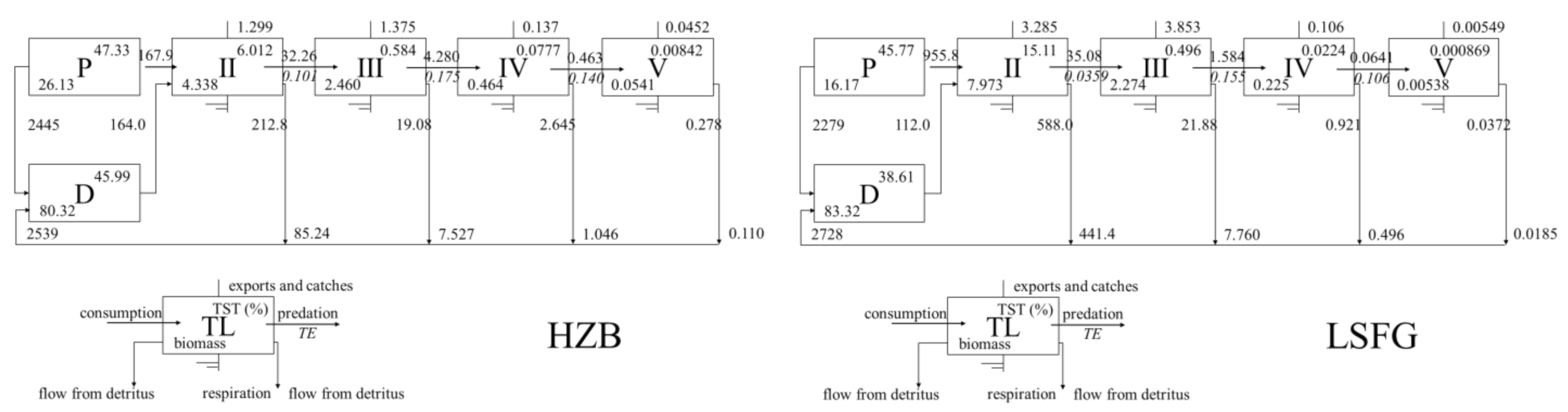
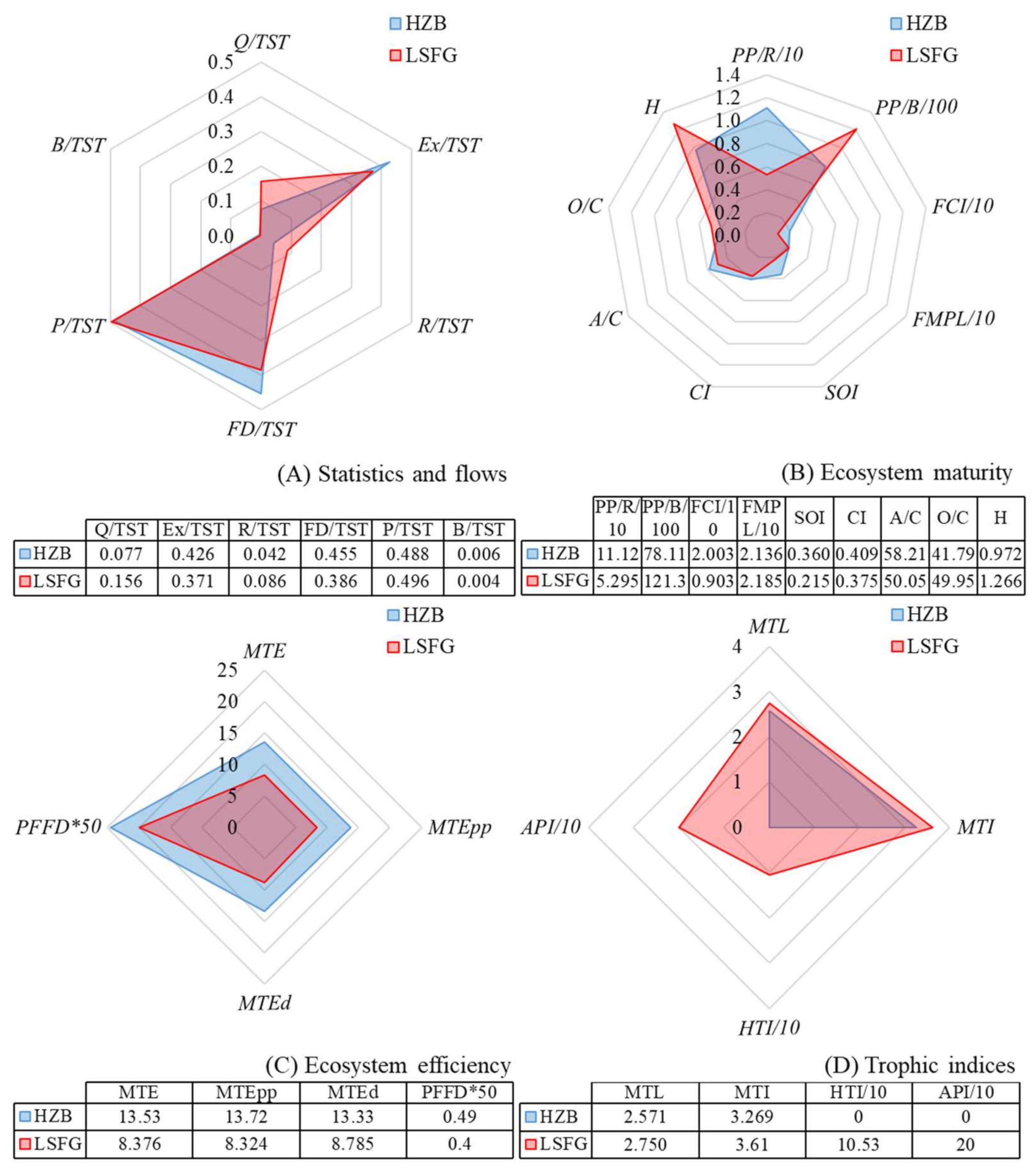
3.2. Properties and Comparison of the HZB and LSFG Ecosystems
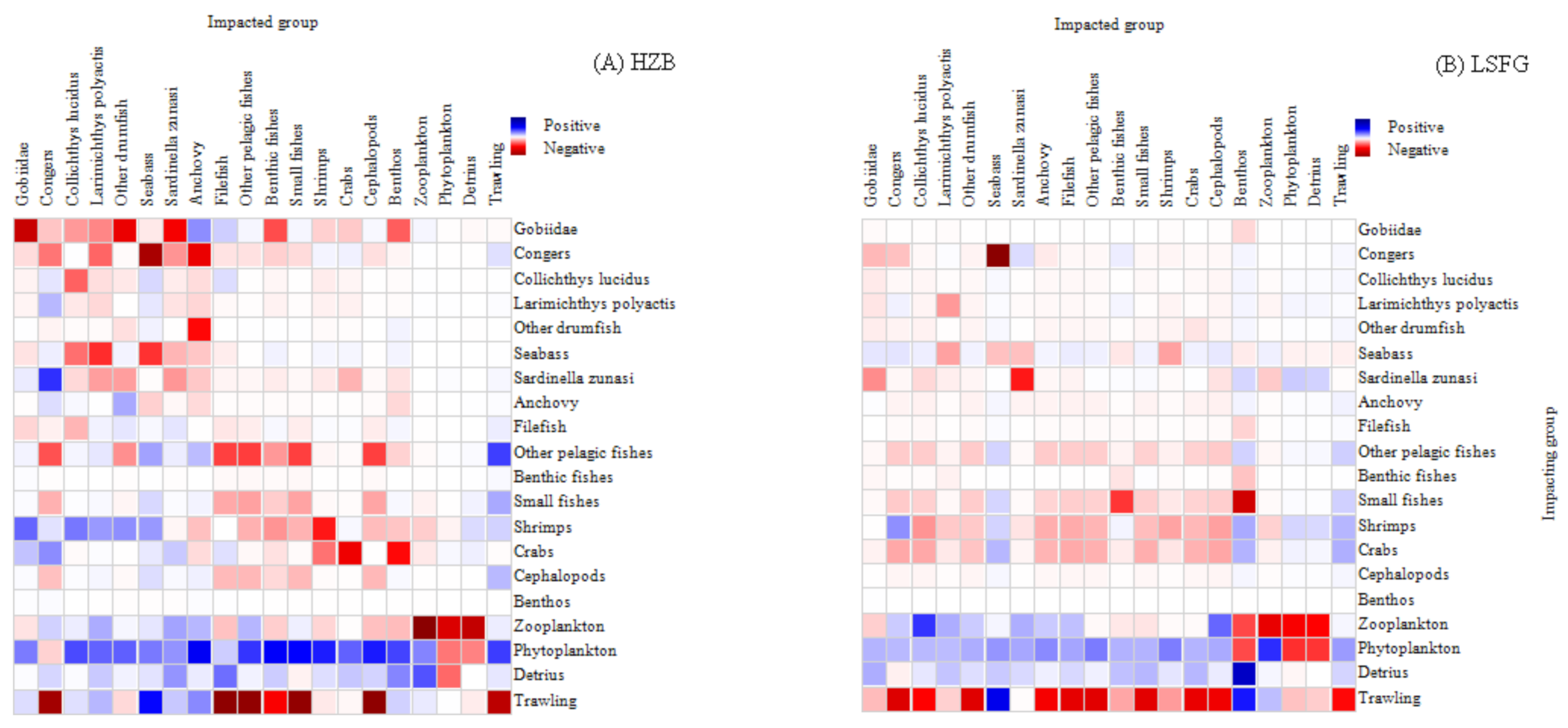
3.3. MTI Analysis
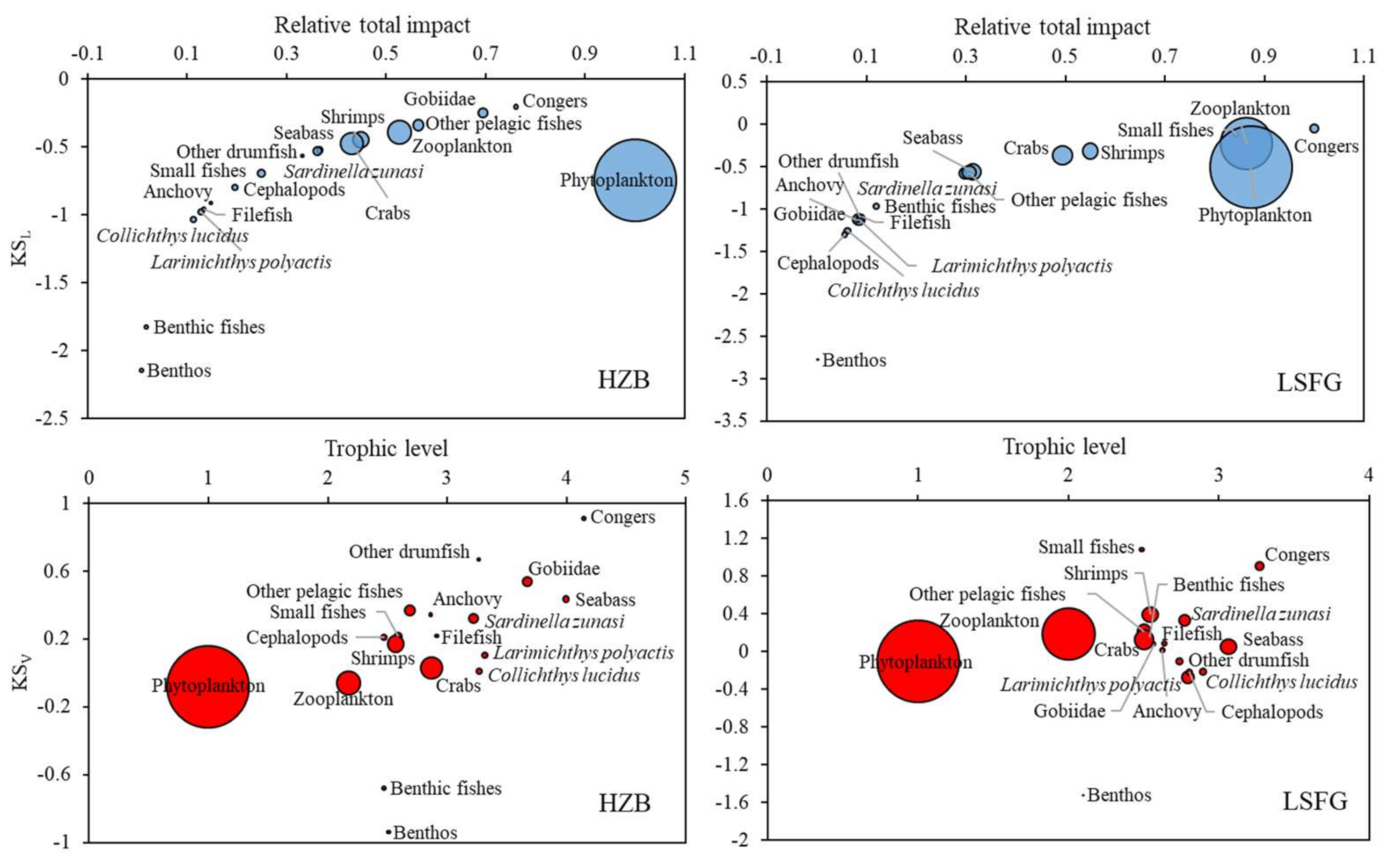
3.4. Keystoneness Analysis
4. Discussion
4.1. Structure and Function of the HZB and LSFG Ecosystems
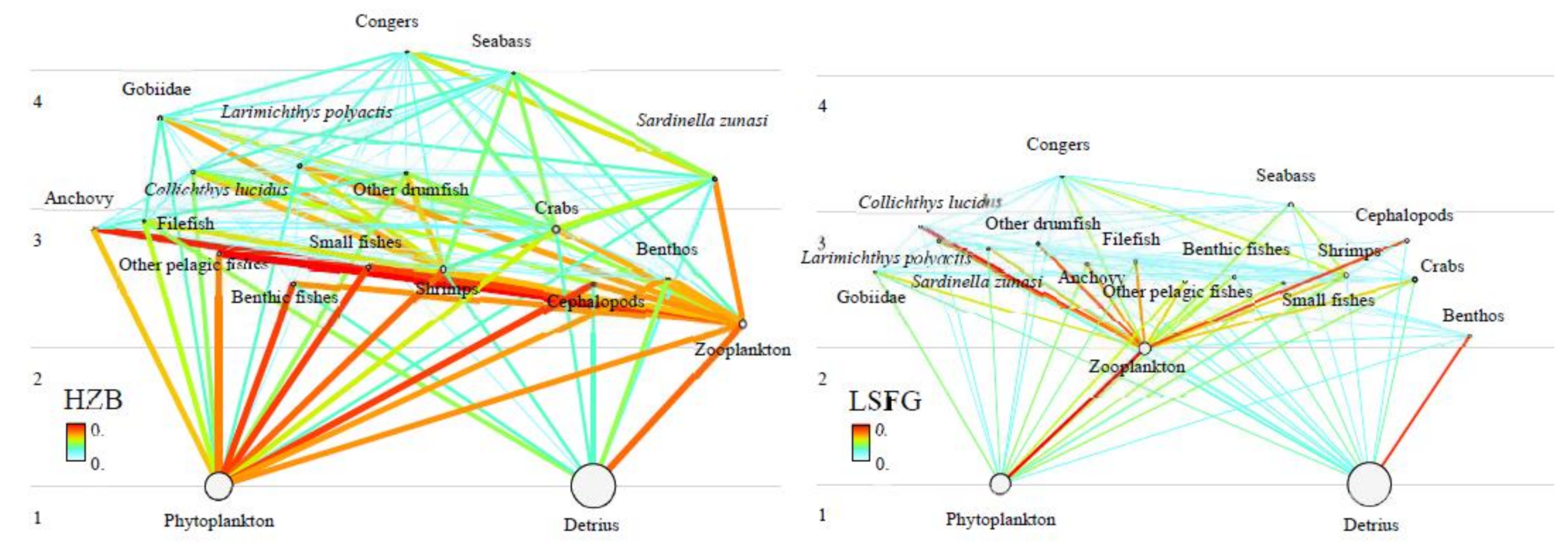
4.2. Trophic Interactions of the HZB and LSFG Ecosystems
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4.3. Ecosystem Maturity of the HZB and LSFG Ecosystems
4.4. Considerations for Assessing Ecosystems through Modelling
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moore, K.J.; Doney, S.C.; Kleypas, J.; Glover, D.; Fung, I.Y. An intermediate complexity marine ecosystem model for the global domain. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 49, 403–462. [Google Scholar] [CrossRef] [Green Version]
- Crowder, L.; Norse, E. Essential ecological insights for marine ecosystem-based management and marine spatial planning. Mar. Policy 2008, 32, 772–778. [Google Scholar] [CrossRef]
- Heim, K.C.; Thorne, L.H.; Warren, J.D.; Link, J.S.; Nye, J.A. Marine ecosystem indicators are sensitive to ecosystem boundaries and spatial scale. Ecol. Indic. 2021, 125, 107522. [Google Scholar] [CrossRef]
- Paine, R.T. Food web complexity and species diversity. Am. Nat. 1966, 100, 65–75. [Google Scholar] [CrossRef]
- Petchey, O.L.; McPhearson, T.; Casey, T.M.; Morin, P.J. Environmental warming alters food-web structure and ecosystem function. Nature 1999, 402, 69–72. [Google Scholar] [CrossRef]
- Polovina, J.J. Model of a coral reef ecosystem I. The ECOPATH model and its application to french frigate shoals. Coral Reefs 1984, 3, 1–11. [Google Scholar] [CrossRef]
- Christensen, V.; Pauly, D. ECOPATH II—A software for balancing steady-state ecosystem models and calculating network characteristics. Ecol. Model. 1992, 61, 169–185. [Google Scholar] [CrossRef]
- Christensen, V.; Walters, C.J.; Pauly, D. Ecopath with Ecosim: A User’s Guide; Fisheries Centre, University of British Columbia: Vancouver, Canada, 2005; Volume 154. [Google Scholar]
- Taghavimotlagh, S.A.; Vahabnezhad, A.; Shojaei, M.G. A trophic model of the coastal fisheries ecosystem of the northern Persian Gulf using a mass balance Ecopath models. Reg. Stud. Mar. Sci. 2021, 42, 101639. [Google Scholar] [CrossRef]
- Milessi, A.C.; Danilo, C.; Laura, R.G.; Daniel, C.; Javier, S.; Rodríguez-Gallego, L. Trophic mass-balance model of a subtropical coastal lagoon, including a comparison with a stable isotope analysis of the food-web. Ecol. Model. 2010, 221, 2859–2869. [Google Scholar] [CrossRef]
- Kong, X.Z.; He, W.; Liu, W.X.; Yang, B. Changes in food web structure and ecosystem functioning of a large, shallow Chinese lake during the 1950s, 1980s and 2000s. Ecol. Model. 2016, 319, 31–41. [Google Scholar] [CrossRef]
- Lemoine, M.; Moens, T.; Vafeiadou, A.M.; Bezerra, L.A. Resource utilization of puffer fish in a subtropical bay as revealed by stable isotope analysis and food web modeling. Mar. Ecol. Prog. Ser. 2019, 626, 161–175. [Google Scholar] [CrossRef]
- Han, D.; Tian, S.; Zhang, Y.; Chen, Y. An evaluation of temporal changes in the trophic structure of Gulf of Maine ecosystem. Reg. Stud. Mar. Sci. 2021, 42, 101635. [Google Scholar] [CrossRef]
- Elliott, M.; Burdon, D.; Hemingway, K.L.; Apitz, S.E. Estuarine, coastal and marine ecosystem restoration: Confusing management and science—A revision of concepts. Estuar. Coast. Shelf Sci. 2007, 74, 349–366. [Google Scholar] [CrossRef]
- Barbier, E.B.; Hacker, S.D.; Koch, E.W.; Stier, A.C.; Silliman, B.R. Estuarine and coastal ecosystems and their services. Treatise Estuar. Coast. Sci. 2011, 12, 109–127. [Google Scholar] [CrossRef]
- Ju, P.L.; Cheung, W.W.L.; Chen, M.R.; Xian, W.W.; Yang, S.Y.; Xiao, J.M. Comparing marine ecosystems of Laizhou and Haizhou bays, China, using ecological indicators estimated from food web models. J. Mar. Syst. 2020, 202, 103238. [Google Scholar] [CrossRef]
- Wang, T.; Li, Y.; Xie, B.; Zhang, H.; Zhang, S. Ecosystem development of haizhou bay ecological restoration area from 2003 to 2013. J. Ocean Univ. China 2017, 16, 7. [Google Scholar] [CrossRef]
- Gao, S.K.; Yu, W.W.; Zhang, S. Trophic level of main organisms in coastal water of Lyusi fishing ground based on stable carbon and nitrogen isotope method. Chin. J. Appl. Ecol. 2020, 31, 301–308. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Zhang, H.J.; Jiao, J.P.; Li, Y.S.; Zhu, K.W. Changes of ecological environment of artificial reef in Haizhou Bay. J. Fish. China 2006, 30, 457–480. [Google Scholar] [CrossRef]
- Xu, J.; Sun, Y.; Xu, Z.L. Fish assembles in the coastal water of Lvsi fishing ground during spring and summer. Chin. J. Appl. Ecol. 2014, 25, 243–250. [Google Scholar]
- Tang, F.H.; Shen, X.Q.; Wang, Y.L. Dynamics of fisheries resources near Haizhou Bay waters. Fish. Sci. 2011, 30, 335–341. [Google Scholar] [CrossRef]
- Xie, B.; Li, Y.K.; Zhang, H.; Zhang, S. Food web foundation and seasonal variation of trophic structure based on stable isotopic technique in the marine ranching of Haizhou Bay. Chin. J. Appl. Ecol. 2017, 28, 2292–2298. [Google Scholar] [CrossRef]
- Lin, Q.; Li, X.S.; Li, Z.Y.; Jin, X.S. Ecological carrying capacity of Chinese shrimp stock enhancement in Laizhou Bay of East China based on Ecopath model. Chin. J. Appl. Ecol. 2013, 24, 1131–1140, (In Chinese with English Abstract). [Google Scholar]
- Wang, T.; Zhang, H.; Zhang, H.; Zhang, S. Ecological carrying capacity of Chinese shrimp stock enhancement in Haizhou Bay of East China based on Ecopath model. J. Fish. Sci. China 2016, 23, 965–975, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Lassalle, G.; Lobry, J.; Le Loc’h, F.; Mackinson, S.; Sanchez, F.; Tomczak, M.T.; Niquil, N. Ecosystem status and functioning: Searching for rules of thumb using an intersite comparison of food-web models of Northeast Atlantic continental shelves. ICES J. Mar. Sci. 2013, 70, 135–149. [Google Scholar] [CrossRef] [Green Version]
- Compilation Committee of Chinese Gulf Records. China’s Gulf Chronicle; Ocean Press: Beijing, China, 1993. [Google Scholar]
- Luo, F.; Li, R.J. 3D Water Environment Simulation for North Jiangsu Offshore Sea Based on EFDC. J. Water Resour. Prot. 2009, 1, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Feng, Y.; Song, Z.Y. Three-dimensional numerical simulation of tidal current in offshore of Haizhou Bay. J. Hohai Univ. 2007, 35, 718–721. [Google Scholar]
- Deehr, R.A.; Luczkovich, J.J.; Hart, K.J.; Clough, L.M.; Johnson, B.J.; Johnson, J.C. Using stable isotope analysis to validate effective trophic levels from Ecopath models of areas closed and open to shrimp trawling in Core Sound, NC, USA. Ecol. Model. 2014, 282, 1–17. [Google Scholar] [CrossRef]
- Cheng, J.H.; William, W.L.; Tony, J.P. Mass-balance ecosystem model of the East China Sea. Prog. Nat. Sci. 2010, 19, 1271–1280. [Google Scholar] [CrossRef]
- Sánchez, F.; Olaso, I. Effects of fisheries on the Cantabrian Sea shelf ecosystem. Ecol. Model. 2004, 172, 151–174. [Google Scholar] [CrossRef]
- Finn, J.T. Measures of ecosystem structure and function derived from analysis of flows. J. Theor. Biol. 1976, 56, 363–380. [Google Scholar] [CrossRef]
- Ortiz, M.; Berrios, F.; Campos, L.; Uribe, R.; Ramirez, A.; Hermosillo-Núñez, B.; González, J.; Rodriguez-Zaragoza, F. Mass balanced trophic models and short-term dynamical simulations for benthic ecological systems of Mejillones and Antofagasta bays (SE Pacific): Comparative network structure and assessment of human impacts. Ecol. Model. 2015, 309, 153–162. [Google Scholar] [CrossRef]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 104, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Christensen, V.; Walters, C.J. Ecopath with Ecosim: Methods, capabilities and limitations. Ecol. Model. 2004, 172, 109–139. [Google Scholar] [CrossRef]
- Ulanowicz, R.E. Growth and Development: Ecosystem Phenomenology; Springer: New York, NY, USA, 1986. [Google Scholar] [CrossRef]
- Angelini, R.; Petrere, M., Jr. A model for the plankton system of the Broa reservoir, São Carlos, Brazil. Ecol. Model. 2000, 12, 131–137. [Google Scholar] [CrossRef]
- Pauly, D.; Christensen, V.; Dalsgaard, J.; Froese, R.; Torres, F., Jr. Fishing down marine food webs. Science 1998, 279, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Bourdaud, P.; Gascuel, D.; Bentorcha, A.; Brind’Amour, A. New trophic indicators and target values for an ecosystem-based management of fisheries. Ecol. Indic. 2016, 61, 588–601. [Google Scholar] [CrossRef]
- Pauly, D.; Watson, R. Background and interpretation of the “Marine Trophic Index” as a measure of biodiversity. Philos. Trans. R. Soc. B 2005, 360, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Christensen, V.; Walters, C.; Pauly, D.; Forrest, R. Ecopath With Ecosim Version 6. User Guide. Lenfest Ocean Futures Project 2008. November 2008. Available online: https://repository.oceanbestpractices.org/bitstream/handle/11329/498/Ewe%20User%20Guide%206.pdf?sequence=2&isAllowed=y (accessed on 20 December 2021).
- Libralato, S.; Christensen, V.; Pauly, D. A method for identifying keystone species in food web models. Ecol. Model. 2006, 195, 153–171. [Google Scholar] [CrossRef]
- Valls, A.; Coll, M.; Christensen, V. Keystone species: Toward an operational concept of marine biodiversity conservation. Ecol. Monogr. 2015, 85, 29–47. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.K.; Yu, N.; Chen, L.Q.; Chen, Y.; Feng, D.X. Ecological modeling on structure and functioning of southern East China Sea ecosystem. Mar. Fish. Res. 2010, 31, 30–39. [Google Scholar]
- Xing, F.; Wang, Y.P.; Gao, J.H.; Zou, X.Q. Seasonal distributions of the concentrations of suspended sediment along Jiangsu coastal sea. Oceanol. Limnol. Sin. 2010, 41, 459–468. [Google Scholar] [CrossRef]
- Lowry, M.; Glasby, T.; Boys, C.; Folpp, H.; Suthers, I.; Gregson, M. Response of fish communities to the deployment of estuarine artificial reefs for fisheries enhancement. Fish. Manag. Ecol. 2014, 2, 42–56. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, H.; Zhang, Q.; Zhang, H.; Zhao, J. Trophic interactions of reef-associated predatory fishes (Hexagrammos otakii and Sebastes schlegelii) in natural and artificial reefs along the coast of North Yellow Sea, China. Sci. Total Environ. 2021, 791, 148250. [Google Scholar] [CrossRef] [PubMed]
- Hattab, T.; Lasram, F.B.R.; Albouy, C.; Romdhane, M.S.; Jarboui, O.; Halouani, G.; Cury, P.; Le Loc’h, F. An ecosystem model of an exploited southern Mediterranean shelf region (Gulf of Gabes, Tunisia) and a comparison with other Mediterranean ecosystem model properties. J. Marine Syst. 2013, 128, 159–174. [Google Scholar] [CrossRef]
- Corrales, X.; Coll, M.; Tecchio, S.; Bellido, J.M.; Fernández, Á.; Palomera, I. Ecosystem structure and fishing impacts in the northwestern Mediterranean Sea using a food web model within a comparative approach. J. Marine Syst. 2015, 148, 183–199. [Google Scholar] [CrossRef]
- Tsagarakis, K.; Coll, M.; Giannoulaki, M.; Somarakis, S.; Papaconstantinou, C.; Machias, A. Food-web traits of the North Aegean Sea ecosystem (Eastern Mediterranean) and comparison with other Mediterranean ecosystems. Estuar. Coast. Shelf Sci. 2010, 88, 233–248. [Google Scholar] [CrossRef]
- Ortiz, M.; Levins, R.; Campos, L.; F Berrios, F.; Jordán, F.C.; Hermosillo-Núñez, B.B.; González, J.; Zaragoza, F.Z.R. Identifying keystone trophic groups in benthic ecosystems: Implications for fisheries management. Ecol. Indic. 2013, 25, 133–140. [Google Scholar] [CrossRef]
- Qian, W.; Chen, J.; Zhang, Q.; Wu, C.; He, Q. Top-down control of foundation species recovery during coastal wetland restoration. Sci. Total Environ. 2021, 769, 144854. [Google Scholar] [CrossRef]
- Pérez-España, H.; Arreguín-Sánchez, F. An inverse relationship between stability and maturity in models of aquatic ecosystems. Ecol. Model. 2001, 145, 189–196. [Google Scholar] [CrossRef]
- Nuttall, M.; Jordaan, A.; Cerrato, R.; Frisk, M. Identifying 120 years of decline in ecosystem structure and maturity of great south bay, New York using the ecopath modelling approach. Ecol. Model. 2011, 222, 3335–3345. [Google Scholar] [CrossRef]
- Bauer, B.; Gustafsson, B.G.; Hyytiäinen, K.; Meier, M.; Muller-Karulis, B.; Saraiva, S.; Tomczak, M.T. Food web and fisheries in the future Baltic sea. Ambio 2019, 48, 1337–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.; Mackay, M.; Novaglio, C.; Fullbrook, L.; Haward, M. The future of ocean governance. Rev. Fish Biol. Fish. 2022, 32, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Lan, C.H.; Hsui, C.Y. The deployment of artificial reef ecosystem: Modelling, simulation and application. Simul. Model. Pract. Theory 2006, 14, 663–675. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, X.; Lozanomontes, H.M.; Loneragan, N.R.; Fan, J. Trophic flows in the marine ecosystem of an artificial reef zone in the Yellow Sea. In Proceedings of the PICES 2015 Annual Meeting: Change and Sustainability of the North Pacific, Qingdao, China, 14–25 October 2015. [Google Scholar]
- Xu, C.J.; Sui, H.Z.; Xu, B.Y.; Zhang, C.L.; Ji, L.P.; Ren, Y.P. The characteristics of food web energy flow in Haizhou Bay based on LIM-MCMC model. J. Fish. Sci. China 2021, 28, 13. [Google Scholar] [CrossRef]
- Han, L.M.; Du, L.J. Investigation and proposal to marine stocking in developing countries. Chin. Fish. Econ. 2015, 1, 16–22. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, S.; Chen, Z.; Lu, Y.; Li, Z.; Zhang, S.; Yu, W. Comparison of Marine Ecosystems of Haizhou Bay and Lvsi Fishing Ground in China Based on the Ecopath Model. Water 2022, 14, 1397. https://doi.org/10.3390/w14091397
Gao S, Chen Z, Lu Y, Li Z, Zhang S, Yu W. Comparison of Marine Ecosystems of Haizhou Bay and Lvsi Fishing Ground in China Based on the Ecopath Model. Water. 2022; 14(9):1397. https://doi.org/10.3390/w14091397
Chicago/Turabian StyleGao, Shike, Ze Chen, Yanan Lu, Zhen Li, Shuo Zhang, and Wenwen Yu. 2022. "Comparison of Marine Ecosystems of Haizhou Bay and Lvsi Fishing Ground in China Based on the Ecopath Model" Water 14, no. 9: 1397. https://doi.org/10.3390/w14091397
APA StyleGao, S., Chen, Z., Lu, Y., Li, Z., Zhang, S., & Yu, W. (2022). Comparison of Marine Ecosystems of Haizhou Bay and Lvsi Fishing Ground in China Based on the Ecopath Model. Water, 14(9), 1397. https://doi.org/10.3390/w14091397